Зоол.бесп.Л.31.Филог.Насекомых..ppt
- Количество слайдов: 17



Зоол. бесп. Л. 31. Филогения насекомых

• Членистоногие возникли от некого предка, сходного с кольчецами, по всей вероятности, в конце докембрия, а первая значительная радиация этой группы началась в кембрии. Среди ископаемых остатков, найденных в нижнепалеозойских отложениях, обнаружены представители всех групп членистоногих, за исключением насекомых. Насекомые появляются в верхнем палеозое, т. е. примерно 350 млн. лет назад. Если в отношении происхождения членистоногих от кольчецов (или от какойто сходной с ними группы) существует известное единодушие, то по вопросу о том, являются ли главные группы членистоногих моно- или полифилетическими, единого мнения нет. Сконцентрируем внимание на той части филогении, которая относится к группе членистоногих Uniramia (одноветвистые) и от которой в конечном счете произошли насекомые.

• К признакам, позволяющим считать кольчецов предками членистоногих, относятся лежащая в основе их строения метамерия, расположение нервной системы на вентральной стороне, а сердца - на дорсальной стороне тела; это признаки, которыми обладают все членистоногие. Несмотря на эти общие для обеих групп признаки взрослых особей, они сильно различаются по ранним стадиям развития. Для кольчецов, так же как для моллюсков, характерно спиральное дробление, тогда как у членистоногих дробление в общем центролецитальное, поэтому одним из крупнейших событий в возникновении членистоногих было резкое изменение раннего развития при сохранении взрослыми особями основного плана строения.

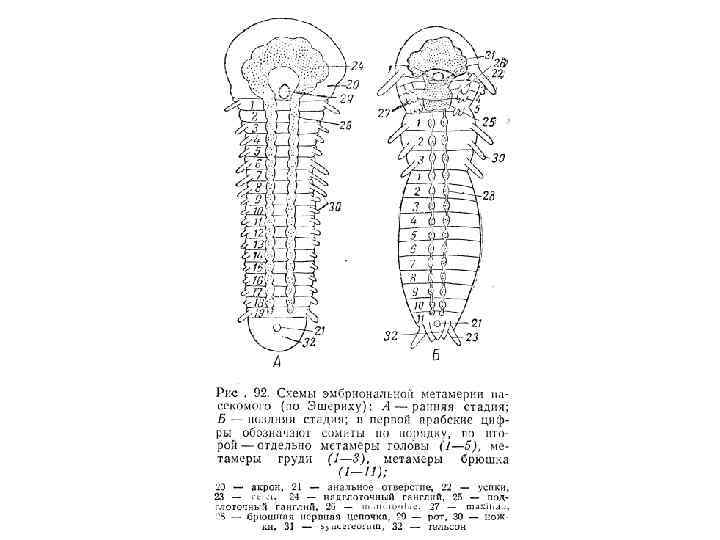
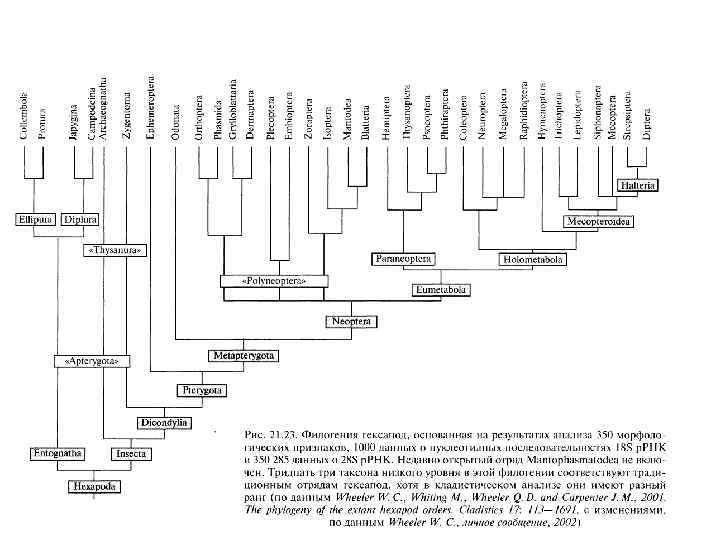
Рис. 8 -10. Филогенетические связи между главными группами членистоногих с упором на линию, давшую начало современным насекомым (Hexapoda).

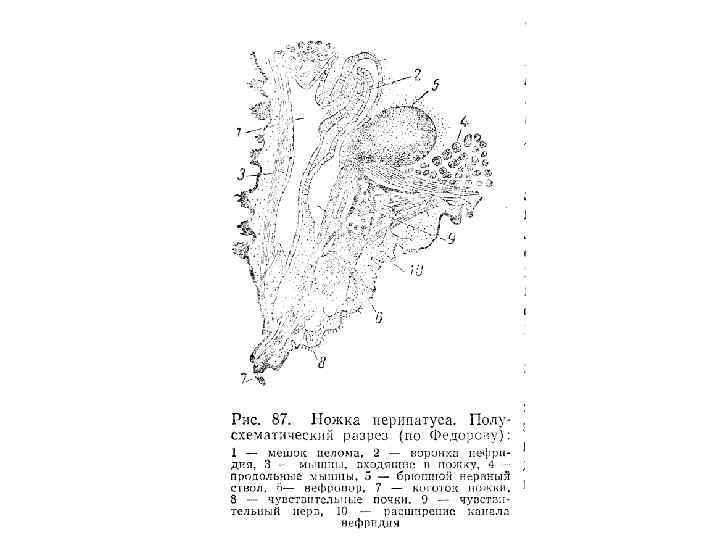
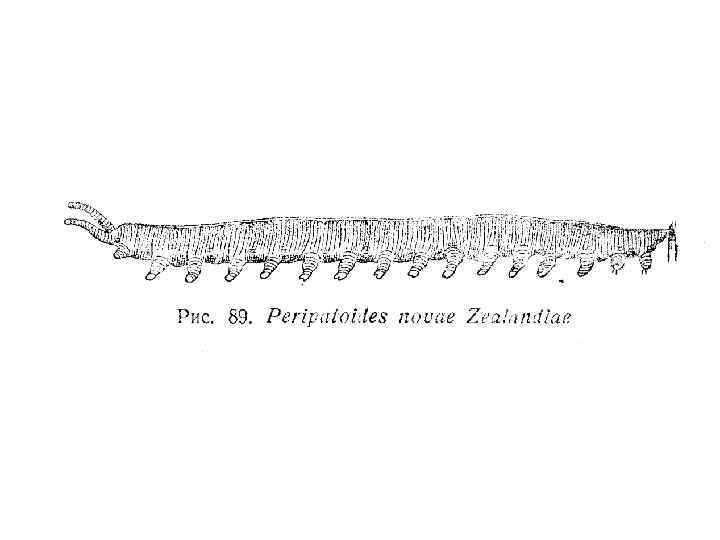
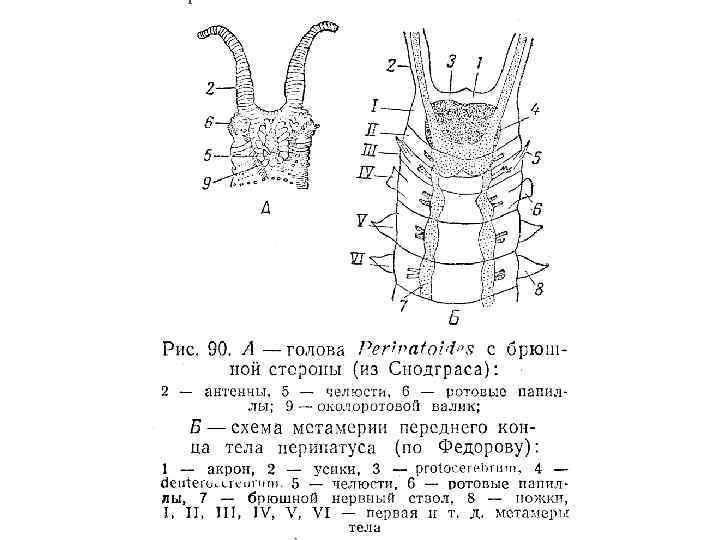
Рис. 8 -11. А-В. Сегментация тела морской полихеты Polidora. А. Ранняя личиночная стадия. Б. Более поздняя личиночная стадия после добавления нескольких сегментов к зоне нарастания на заднем конце тела. В. Молодая взрослая особь. Г-Е. Зародыши и взрослая форма онихофор. Г. Вентральная сторона зародыша Peripatoides novaezealandiae на ранней стадии образования зародышевой полоски; как и у кольчецов, добавление новых сегментов происходит на заднем конце тела. Д. Вентральная сторона головы зародыша P. capensis, состоящей из трех сегментов антеннального, челюстного и ротового и головных сосочков. Е. Обобщенная взрослая онихофора (рисунок сделан с зафиксированных экземпляров); все туловищные сегменты одинаковы, и на каждом из них имеется по одной паре членистых ходильных ножек с коготком на конце (А-В - Wilson, 1928; Г - Sheldon, 1889; Д - Sedgwick, 1888).
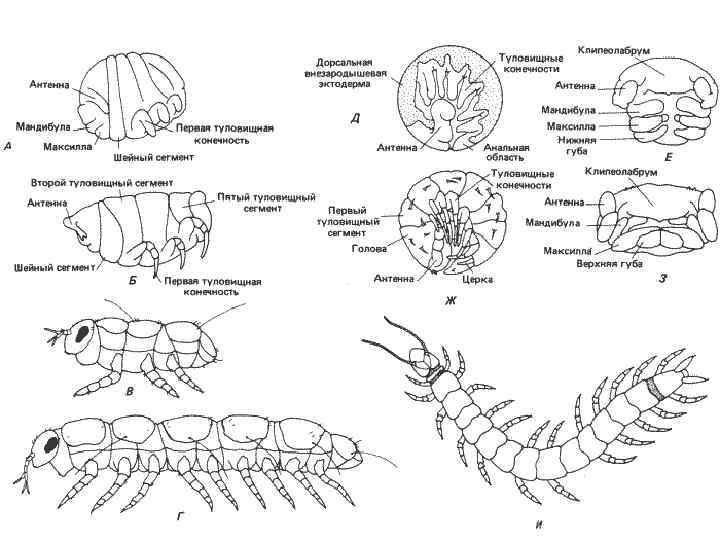
 дигнатической многоножки Pauropus,")
• Рис. 8 -12. Зародышевая, личиночная и взрослая стадия (А-Г) дигнатической многоножки Pauropus, зародышевые стадии тригнатической многоножки Hanseniella (Д-3) и взрослая тригнатическая многоножка Scutigerella (И). А. Десятидневный зародыш с намечающейся сегментацией тела. Б. Стадия ранней куколки. В. Личинка первого возраста вскоре после вылупления. Г. Взрослая особь после добавления туловищных сегментов в зоне нарастания на заднем конце тела. Д. Вся зародышевая полоска 8 -дневного зародыша (вид сбоку). Е. Ротовые части того же зародыша, что на Д (вид снизу). Ж. Зародыш непосредственно перед вылуплением (вид сбоку). 3. Голова и ротовые части того же зародыша, что на Ж (вид снизу). И. Взрослая особь (вид сверху). (А, Б, Д-3 - Anderson, 1973; В, Г, И - по Snodgrass, 1952; с изменениями).
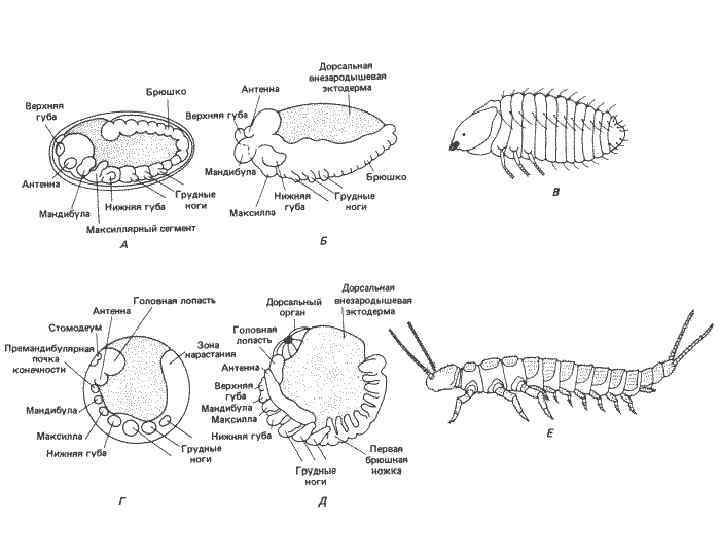

• Рис. 8 -13. Вид сбоку сегментированных зародышей и вылупившихся особей крылатого насекомого Bruchidius (А-В) и бескрылого Campodea (Г-Ε). А. Полностью сегментированная зародышевая полоска. Б. Укорочение зародышевой полоски. В. Стадия вылупления. Г. Сегментирующаяся зародышевая полоска; сегментация происходит на заднем конце полоски. Д. Полностью сформировавшаяся и сегментированная зародышевая полоска. Е. Взрослая особь. При сравнении рис. В и Ε с многоножками, изображенными на рис. 8 -12, В и Г, бросается в глаза сильное сходство между этими формами, которое отчасти и послужило основанием для гипотезы о происхождении примитивных насекомых в результате педогенеза от предка, сходного с многоножками (А-В - Anderson, 1973; E Snodgrass, 1952).

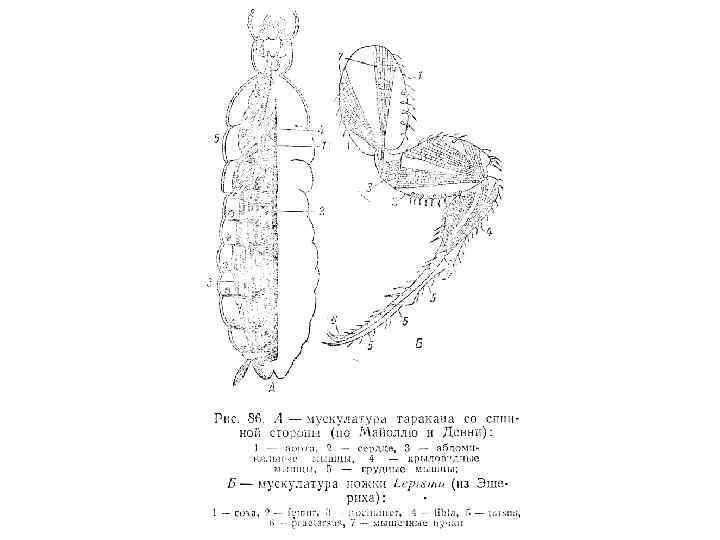
Рис. 8 -14. Схема предполагаемой эволюции характера сегментации насекомых в процессе филогенеза. А. Кольчецы. Б. Онихофоры. В. Многоножки. Г. Бескрылые насекомые. Д. Крылатые насекомые. Сопоставляя эту схему с рис. 8 -9, можно видеть, что постепенное выпадение гомеозисных локусов приводит к прогрессирующему упрощению сегментации у дрозофилы, которое до некоторой степени имитирует филогению насекомых. (Snodgrass, 1935).

Зоол.бесп.Л.31.Филог.Насекомых..ppt