

Генетика 2. Презентации лекций.ppt
- Количество слайдов: 88
 Жизненный цикл клетки: интерфаза и период деления.
Жизненный цикл клетки: интерфаза и период деления.
, обладающая собственным обменом") Клетка – элементарная единица строения и жизнедеятельности всех организмов (кроме вирусов), обладающая собственным обменом веществ, способная к самостоятельному существованию, самовоспроизведению и развитию.
Клетка – элементарная единица строения и жизнедеятельности всех организмов (кроме вирусов), обладающая собственным обменом веществ, способная к самостоятельному существованию, самовоспроизведению и развитию.
 Последовательность событий в период существования клетки, происходящих между образованием клетки, ее делением на дочерние клетки, а затем гибелью, называют клеточным циклом.
Последовательность событий в период существования клетки, происходящих между образованием клетки, ее делением на дочерние клетки, а затем гибелью, называют клеточным циклом.
 Вновь появившаяся клетка сначала растет и дифференцируется, затем выполняет свои специфические функции. Это время существования клетки называется периодом покоя. Обязательной частью жизненного цикла является образование новых клеток, которое возможно только путем деления.
Вновь появившаяся клетка сначала растет и дифференцируется, затем выполняет свои специфические функции. Это время существования клетки называется периодом покоя. Обязательной частью жизненного цикла является образование новых клеток, которое возможно только путем деления.
 Клетки лука в различных фазах клеточного цикла
Клетки лука в различных фазах клеточного цикла
 В настоящее время известно три типа деления эукариотических клеток: амитоз, мейоз. Амитоз – прямое деление. Клетка в этом случае делится путем прямой перетяжки.
В настоящее время известно три типа деления эукариотических клеток: амитоз, мейоз. Амитоз – прямое деление. Клетка в этом случае делится путем прямой перетяжки.
 Равномерного распределения наследственного материала между образовавшимися клетками не происходит. Возможно образование двуядерных клеток. Амитоз характерен для погибающих или измененных клеток.
Равномерного распределения наследственного материала между образовавшимися клетками не происходит. Возможно образование двуядерных клеток. Амитоз характерен для погибающих или измененных клеток.
 При амитозе морфологически сохраняется интерфазное состояние ядра, хорошо видны ядрышко и ядерная оболочка. Репликация ДНК отсутствует, спирализация хроматина не происходит, хромосомы не выявляются. При амитозе делится только ядро.
При амитозе морфологически сохраняется интерфазное состояние ядра, хорошо видны ядрышко и ядерная оболочка. Репликация ДНК отсутствует, спирализация хроматина не происходит, хромосомы не выявляются. При амитозе делится только ядро.
 Наследственный материал делится случайным образом. Отсутствие цитокинеза приводит к образованию двуядерных клеток, которые в последующем не способны к митозу. При повторных амитозах могут образовываться многоядерные клетки.
Наследственный материал делится случайным образом. Отсутствие цитокинеза приводит к образованию двуядерных клеток, которые в последующем не способны к митозу. При повторных амитозах могут образовываться многоядерные клетки.
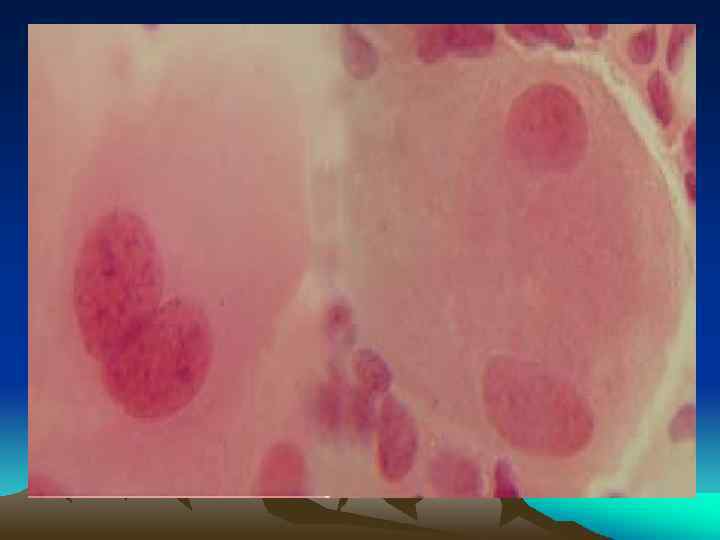
 Митоз – непрямое деление соматических клеток, в результате которого из одной клетки образуются две одинаковых дочерних клетки. Митоз - это универсальный способ увеличения количества или замещения погибших клеток. Период между делениями клетки называется интерфазой. Интерфаза и митоз ( М) составляют клеточный цикл.
Митоз – непрямое деление соматических клеток, в результате которого из одной клетки образуются две одинаковых дочерних клетки. Митоз - это универсальный способ увеличения количества или замещения погибших клеток. Период между делениями клетки называется интерфазой. Интерфаза и митоз ( М) составляют клеточный цикл.
 Клеточный цикл состоит из четырех периодов: пресинтетический -G 1; синтетический - S; постсинтетический – G 2; митоз (М). Интерфаза включает три периода: пресинтетический -G 1; синтетический - S; постсинтетический – G 2.
Клеточный цикл состоит из четырех периодов: пресинтетический -G 1; синтетический - S; постсинтетический – G 2; митоз (М). Интерфаза включает три периода: пресинтетический -G 1; синтетический - S; постсинтетический – G 2.

 Интерфаза – период интенсивного синтеза и роста. В этот период синтезируется много веществ, необходимых для роста, происходит репликация ДНК. В пресинтетический период ( G 1) клетка растет, осуществляет синтез белков и РНК, накапливает богатые энергией вещества.
Интерфаза – период интенсивного синтеза и роста. В этот период синтезируется много веществ, необходимых для роста, происходит репликация ДНК. В пресинтетический период ( G 1) клетка растет, осуществляет синтез белков и РНК, накапливает богатые энергией вещества.
 Клеточный цикл
Клеточный цикл
 характеризуется удвоением материала. На этой стадии продолжается синтез белков и РНК.") Синтетический период (S) характеризуется удвоением материала. На этой стадии продолжается синтез белков и РНК. В постсинтетическом периоде (G 2 ) клетка готовится к делению, накапливая энергию и белки. В клетке увеличивается количество митохондрий. Делится центросома.
Синтетический период (S) характеризуется удвоением материала. На этой стадии продолжается синтез белков и РНК. В постсинтетическом периоде (G 2 ) клетка готовится к делению, накапливая энергию и белки. В клетке увеличивается количество митохондрий. Делится центросома.
 завершается репликация ДНК. В состав каждой хромосомы уже входят") В постсинтетической стадии (G 2) завершается репликация ДНК. В состав каждой хромосомы уже входят две двойных молекулы ДНК, которые являются точной копией исходной молекулы ДНК. Количество ДНК в клетке на постсинтетической стадии обозначается символом 4 с. Синтезируются вещества, необходимые для деления клетки. В конце интерфазы процессы синтеза прекращаются.
В постсинтетической стадии (G 2) завершается репликация ДНК. В состав каждой хромосомы уже входят две двойных молекулы ДНК, которые являются точной копией исходной молекулы ДНК. Количество ДНК в клетке на постсинтетической стадии обозначается символом 4 с. Синтезируются вещества, необходимые для деления клетки. В конце интерфазы процессы синтеза прекращаются.
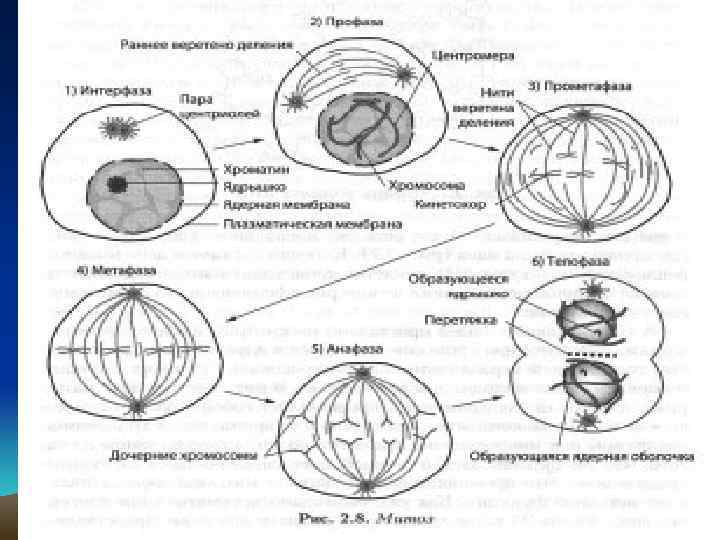
 В профазе хромосомы спирализуются, конденсируются, становятся видимыми в виде двух хроматид, образованных в процессе редупликации и соединенных центромерами. Редупликация происходит так, что в каждой хроматиде имеется одна «старая» и одна вновь синтезированная нить молекулы ДНК. Редупликация – удвоение, процесс самовоспроизведения молекулы нуклеиновых кислот.
В профазе хромосомы спирализуются, конденсируются, становятся видимыми в виде двух хроматид, образованных в процессе редупликации и соединенных центромерами. Редупликация происходит так, что в каждой хроматиде имеется одна «старая» и одна вновь синтезированная нить молекулы ДНК. Редупликация – удвоение, процесс самовоспроизведения молекулы нуклеиновых кислот.
 В профазе в цитоплазме формируется специальный аппарат деления – ахроматиновое веретено, состоящее из белков, которые накопились в период интерфазы. Постепенно вновь образованные хромосомы укорачиваются на 4% от первоначальной длины. В конце профазы исчезает ядрышко, ядерная мембрана растворяется, хромосомы оказываются в цитоплазме и начинают двигаться по направлению к экватору.
В профазе в цитоплазме формируется специальный аппарат деления – ахроматиновое веретено, состоящее из белков, которые накопились в период интерфазы. Постепенно вновь образованные хромосомы укорачиваются на 4% от первоначальной длины. В конце профазы исчезает ядрышко, ядерная мембрана растворяется, хромосомы оказываются в цитоплазме и начинают двигаться по направлению к экватору.
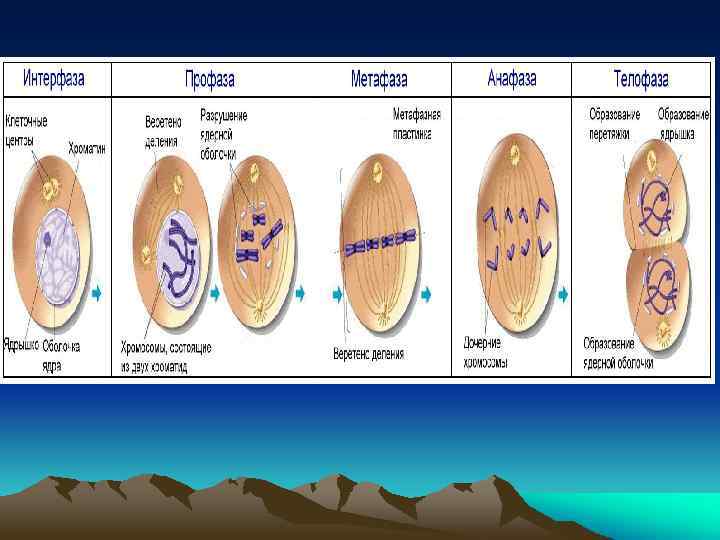
 Метафаза. Начало этой фазы называется прометафаза. В прометафазе хромосомы располагаются в цитоплазме довольно беспорядочно. Формируется митотический аппарат, в состав которого входит веретено деления и центриоли иные центры организации микротрубочек. Прометафаза завершается, как только центромеры всех хромосом оказываются в экваториальной плоскости.
Метафаза. Начало этой фазы называется прометафаза. В прометафазе хромосомы располагаются в цитоплазме довольно беспорядочно. Формируется митотический аппарат, в состав которого входит веретено деления и центриоли иные центры организации микротрубочек. Прометафаза завершается, как только центромеры всех хромосом оказываются в экваториальной плоскости.
 После формирования митотического аппарата хромосомы начинают перемещаться в экваториальную плоскость клетки; это движение хромосом называется метакинез. В метафазе хромосомы максимально спирализованы. Центромеры хромосом располагаются в экваториальной плоскости клетки независимо друг от друга. Полюсные нити веретена деления тянутся от полюсов клетки к хромосомам, а хромосомальные – от центромер (кинетохоров) – к полюсам. Совокупность хромосом в экваториальной плоскости клетки образует метафазную пластинку.
После формирования митотического аппарата хромосомы начинают перемещаться в экваториальную плоскость клетки; это движение хромосом называется метакинез. В метафазе хромосомы максимально спирализованы. Центромеры хромосом располагаются в экваториальной плоскости клетки независимо друг от друга. Полюсные нити веретена деления тянутся от полюсов клетки к хромосомам, а хромосомальные – от центромер (кинетохоров) – к полюсам. Совокупность хромосом в экваториальной плоскости клетки образует метафазную пластинку.
 В анафазе происходит разделение хромосом на хроматиды. С этого момента каждая хроматида становится самостоятельной однохроматидной хромосо мой, в основе которой лежит одна молекула ДНК. Однохроматидные хромосомы в составе анафазных групп расходятся к полюсам клетки. При расхождении хромосомальные микротрубочки укорачиваются, а полюсные – удлиняются. При этом полюсные и хромосомальные нити скользят вдоль друга.
В анафазе происходит разделение хромосом на хроматиды. С этого момента каждая хроматида становится самостоятельной однохроматидной хромосо мой, в основе которой лежит одна молекула ДНК. Однохроматидные хромосомы в составе анафазных групп расходятся к полюсам клетки. При расхождении хромосомальные микротрубочки укорачиваются, а полюсные – удлиняются. При этом полюсные и хромосомальные нити скользят вдоль друга.
 В телофазе веретено деления разрушается. Хромосомы у полюсов клетки деспирализуются, вокруг них формируются ядерные оболочки. В клетке образуются два ядра, генетически идентичные исходному ядру. Содержание ДНК в дочерних ядрах становится равным 2 c.
В телофазе веретено деления разрушается. Хромосомы у полюсов клетки деспирализуются, вокруг них формируются ядерные оболочки. В клетке образуются два ядра, генетически идентичные исходному ядру. Содержание ДНК в дочерних ядрах становится равным 2 c.
 В цитокинезе происходит разделение цитоплазмы и формирование мембран дочерних клеток. У животных цитокинез происходит путем перешнуровывания клетки. У растений цитокинез происходит иначе: в экваториальной плоскости образуются пузырьки, которые сливаются с образованием двух параллельных мембран. На этом митоз завершается, и наступает очередная интерфаза.
В цитокинезе происходит разделение цитоплазмы и формирование мембран дочерних клеток. У животных цитокинез происходит путем перешнуровывания клетки. У растений цитокинез происходит иначе: в экваториальной плоскости образуются пузырьки, которые сливаются с образованием двух параллельных мембран. На этом митоз завершается, и наступает очередная интерфаза.
 Цитокинез на завершающей стадии в животной клетке.
Цитокинез на завершающей стадии в животной клетке.
 Мейозом называется особый способ деления эукариотических клеток, при котором исходное число хромосом уменьшается в 2 раза (от древнегреч. «мейон» – меньше – и от «мейозис» – уменьшение). Часто уменьшение числа хромосом называется редукцией. Исходное число хромосом в мейоцитах (клетках, вступающих в мейоз) в результате мейоза, называется гаплоидным хромосомным числом (n).
Мейозом называется особый способ деления эукариотических клеток, при котором исходное число хромосом уменьшается в 2 раза (от древнегреч. «мейон» – меньше – и от «мейозис» – уменьшение). Часто уменьшение числа хромосом называется редукцией. Исходное число хромосом в мейоцитах (клетках, вступающих в мейоз) в результате мейоза, называется гаплоидным хромосомным числом (n).
 гомологичных хромосом с последующим расхождением их в разные") Главной особенностью мейоза является конъюгация (спаривание) гомологичных хромосом с последующим расхождением их в разные клетки. Мейотическое распределение хромосом по дочерним клеткам называется сегрегацией хромосом.
Главной особенностью мейоза является конъюгация (спаривание) гомологичных хромосом с последующим расхождением их в разные клетки. Мейотическое распределение хромосом по дочерним клеткам называется сегрегацией хромосом.
 Мейоз подразделяется на профазу, метафазу, анафазу, телофазу. Эти стадии имеются в первом делении мейоза, затем еще раз повторяются во втором. Интерфаза, предшествующая мейозу, аналогична митотической и включает удвоение хромосом в S-периоде. Первое клеточное деление мейоза – мейоз I, начинается с профазы I. Профаза I состоит из пяти подстадий: лептотены, зиготены, пахитены, диплотены, диакинеза.
Мейоз подразделяется на профазу, метафазу, анафазу, телофазу. Эти стадии имеются в первом делении мейоза, затем еще раз повторяются во втором. Интерфаза, предшествующая мейозу, аналогична митотической и включает удвоение хромосом в S-периоде. Первое клеточное деление мейоза – мейоз I, начинается с профазы I. Профаза I состоит из пяти подстадий: лептотены, зиготены, пахитены, диплотены, диакинеза.
 На стадии лептотены происходит спирализация и уплотнение хромосом, как в процессе митоза, а именно, как в профазе митоза. Нити хромосом в этот период в основном одиночные, но иногда на концах могут иметь раздвоение.
На стадии лептотены происходит спирализация и уплотнение хромосом, как в процессе митоза, а именно, как в профазе митоза. Нити хромосом в этот период в основном одиночные, но иногда на концах могут иметь раздвоение.
 сначала отдельных участков гомологичных хромосом, а затем они") На стадии зиготены происходит соединение (конъюгация) сначала отдельных участков гомологичных хромосом, а затем они соединяются по всей длине. Две сцепленных таким образом хромосомы называются бивалентом, который состоит из четырех хроматид. В это время происходит очень важный процесс обмена участками хроматид между гомологичными хромосомами, который называется кроссинговер.
На стадии зиготены происходит соединение (конъюгация) сначала отдельных участков гомологичных хромосом, а затем они соединяются по всей длине. Две сцепленных таким образом хромосомы называются бивалентом, который состоит из четырех хроматид. В это время происходит очень важный процесс обмена участками хроматид между гомологичными хромосомами, который называется кроссинговер.
 Согласно теории Янссенса–Дарлингтона, при развитии кроссинговера гомологичные хромосомы с гаплотипами хроматид АВ и ab образуют биваленты. В одной из хроматид в первой хромосоме происходит разрыв на участке А–В, тогда как в прилежащей хроматиде второй хромосомы происходит разрыв на участке a–b. Клетка стремится исправить повреждение с помощью ферментов репарации–рекомбинации и присоединить фрагменты хроматид. Однако при этом возможно присоединение крест–накрест (кроссинговер), и образуются рекомбинантные гаплотипы (хроматиды) Ab и а. В.
Согласно теории Янссенса–Дарлингтона, при развитии кроссинговера гомологичные хромосомы с гаплотипами хроматид АВ и ab образуют биваленты. В одной из хроматид в первой хромосоме происходит разрыв на участке А–В, тогда как в прилежащей хроматиде второй хромосомы происходит разрыв на участке a–b. Клетка стремится исправить повреждение с помощью ферментов репарации–рекомбинации и присоединить фрагменты хроматид. Однако при этом возможно присоединение крест–накрест (кроссинговер), и образуются рекомбинантные гаплотипы (хроматиды) Ab и а. В.
 Рекомбинантные хроматиды, которые образовались в ходе кроссинговера, несут новые сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются кроссоверными, с их участием разовьются кроссоверные гаметы, зиготы и особи.
Рекомбинантные хроматиды, которые образовались в ходе кроссинговера, несут новые сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются кроссоверными, с их участием разовьются кроссоверные гаметы, зиготы и особи.
") Хроматиды, которые не участвовали в кроссинговере, сохраняют исходные сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются некроссоверными; с их участием разовьются некроссоверные гаметы, зиготы и особи.
Хроматиды, которые не участвовали в кроссинговере, сохраняют исходные сочетания аллелей. Такие хроматиды (однохроматидные хромосомы) называются некроссоверными; с их участием разовьются некроссоверные гаметы, зиготы и особи.

 Кроссинговер делает возможным образование новых комбинаций отцовских и материнских генов в половых клетках. Гомологичные хромосомы после кроссинговера не расходятся, поэтому сестринские хроматиды обеих хромосом остаются прочно связанными вплоть до анафазы.
Кроссинговер делает возможным образование новых комбинаций отцовских и материнских генов в половых клетках. Гомологичные хромосомы после кроссинговера не расходятся, поэтому сестринские хроматиды обеих хромосом остаются прочно связанными вплоть до анафазы.
 Кроссинговер – это сложный физиологобиохимический процесс, который находится под генетическим контролем клетки и подвержен влиянию факторов внешней среды. При этом обнаружены гены, выполняющие функции запирателей кроссинговера, но однако есть также гены, повышающие его частоту
Кроссинговер – это сложный физиологобиохимический процесс, который находится под генетическим контролем клетки и подвержен влиянию факторов внешней среды. При этом обнаружены гены, выполняющие функции запирателей кроссинговера, но однако есть также гены, повышающие его частоту
 Обнаружено, что на частоту кроссинговера влияют возраст организма, а также экзогенные факторы: температура, радиация, концентрация солей, химические мутагены, лекарства, гормоны. При большинстве указанных воздействий частота кроссинговера повышается.
Обнаружено, что на частоту кроссинговера влияют возраст организма, а также экзогенные факторы: температура, радиация, концентрация солей, химические мутагены, лекарства, гормоны. При большинстве указанных воздействий частота кроссинговера повышается.
 может служить мерой") Частота различных типов рекомбинаций (мейотический, митотический кроссинговер и сестринские хроматидные обмены) может служить мерой действия мутагенов, канцерогенов, антибиотиков и др.
Частота различных типов рекомбинаций (мейотический, митотический кроссинговер и сестринские хроматидные обмены) может служить мерой действия мутагенов, канцерогенов, антибиотиков и др.
 В целом кроссинговер представляет собой один из регулярных генетических процессов, контролируемых многими генами как непосредственно, так и через физиологическое состояние мейотических или митотических клеток.
В целом кроссинговер представляет собой один из регулярных генетических процессов, контролируемых многими генами как непосредственно, так и через физиологическое состояние мейотических или митотических клеток.
 Благодаря сцепленному наследованию удачные сочетания аллелей оказываются относительно устойчивыми. В результате образуются группы генов, каждая из которых функционирует как единый суперген, контролирующий несколько признаков.
Благодаря сцепленному наследованию удачные сочетания аллелей оказываются относительно устойчивыми. В результате образуются группы генов, каждая из которых функционирует как единый суперген, контролирующий несколько признаков.
 В то же время, в ходе кроссинговера возникают рекомбинации – т. е. новые комбинации аллелей. Таким образом, кроссинговер повышает комбинативную изменчивость организмов.
В то же время, в ходе кроссинговера возникают рекомбинации – т. е. новые комбинации аллелей. Таким образом, кроссинговер повышает комбинативную изменчивость организмов.
 Стадия пахитены наступает к концу конъюгации и характеризуется гаплоидным числом утолщенных и укороченных бивалентов. На стадии пахитены можно видеть ядрышки, прикрепленные к определенным участкам хромосом в области вторичных перетяжек.
Стадия пахитены наступает к концу конъюгации и характеризуется гаплоидным числом утолщенных и укороченных бивалентов. На стадии пахитены можно видеть ядрышки, прикрепленные к определенным участкам хромосом в области вторичных перетяжек.
 В конце стадии пахитены появляется продольная щель, и на стадиии диплотены гомологичные хромосомы почти расходятся друг от друга. На этой стадии каждая хромосома состоит из двух хроматид, которые сцеплены в некоторых участках. Эти сцепления называются хиазмами. Каждый бивалент содержит две или три хиазмы, хотя длинные хромосомы могут иметь их больше, чем короткие.
В конце стадии пахитены появляется продольная щель, и на стадиии диплотены гомологичные хромосомы почти расходятся друг от друга. На этой стадии каждая хромосома состоит из двух хроматид, которые сцеплены в некоторых участках. Эти сцепления называются хиазмами. Каждый бивалент содержит две или три хиазмы, хотя длинные хромосомы могут иметь их больше, чем короткие.
 В стадию диакинеза хиазмы начинают смещаться к концам хроматид и постепенно исчезать. Биваленты смещаются в экваториальную плоскость. Ядерная мембрана и ядрышки растворяются, формируется веретено деление, заканчивается профаза I.
В стадию диакинеза хиазмы начинают смещаться к концам хроматид и постепенно исчезать. Биваленты смещаются в экваториальную плоскость. Ядерная мембрана и ядрышки растворяются, формируется веретено деление, заканчивается профаза I.
 Во время метафазы I хромосомы располагаются в экваториальной области, образуя метафазную пластинку. Нити веретена направляются только к одному полюсу, прикрепляются к центромерам каждого бивалента. Гомологичные хромосомы отделяются друг от друга и растягиваются к разным полюсам. В редуцированный гаплоидный набор попадает по одной гомологичной хромосоме из каждого бивалента.
Во время метафазы I хромосомы располагаются в экваториальной области, образуя метафазную пластинку. Нити веретена направляются только к одному полюсу, прикрепляются к центромерам каждого бивалента. Гомологичные хромосомы отделяются друг от друга и растягиваются к разным полюсам. В редуцированный гаплоидный набор попадает по одной гомологичной хромосоме из каждого бивалента.
 Анафаза I характеризуется окончательным разделением гомологичных хромосом. Нити веретена оттягивают по паре хроматид к противоположным полюсам клетки. Хромосомы разделяются на два гаплоидных набора, попадающих в дочерние клетки.
Анафаза I характеризуется окончательным разделением гомологичных хромосом. Нити веретена оттягивают по паре хроматид к противоположным полюсам клетки. Хромосомы разделяются на два гаплоидных набора, попадающих в дочерние клетки.
 После завершения перемещения гомологичных центромер к противоположным полюсам начинается короткая по времени телофаза I, что означает завершение первого деления мейоза.
После завершения перемещения гомологичных центромер к противоположным полюсам начинается короткая по времени телофаза I, что означает завершение первого деления мейоза.
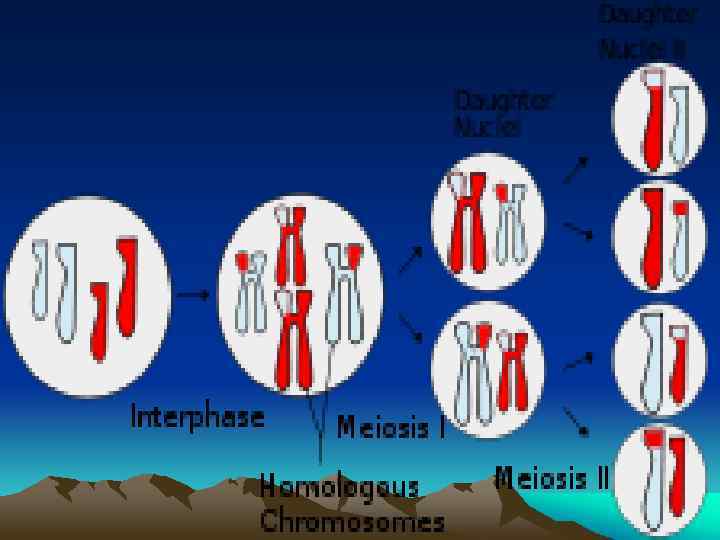
 К началу второго деления мейоза хромосомы уже удвоены, состоят из 2 -х хроматид, соединенных в области центромеры. При этом каждая хроматида содержит гаплоидный набор хромосом. В профазе II хромосомы спирализуются, исчезают ядерная оболочка и ядрышко, формируется ахроматиновое волокно, центриоли перемещаются к полюсам. В период метафазы II хромосомы прикрепляются к нитям веретена деления в области центромеры.
К началу второго деления мейоза хромосомы уже удвоены, состоят из 2 -х хроматид, соединенных в области центромеры. При этом каждая хроматида содержит гаплоидный набор хромосом. В профазе II хромосомы спирализуются, исчезают ядерная оболочка и ядрышко, формируется ахроматиновое волокно, центриоли перемещаются к полюсам. В период метафазы II хромосомы прикрепляются к нитям веретена деления в области центромеры.
 В анафазе II после продольного деления центромеры одна хроматида идет к одному полюсу, другая – к другому и превращается таким образом в хромосомы. В телофазе II образуются 4 гаплоидных ядра. В клетке происходит цитокинез, в результате которого из 2 -х предшествующих клеток получаются четыре. Каждая клетка содержит гаплоидный набор хромосом, структура которых состоит из одной хроматиды.
В анафазе II после продольного деления центромеры одна хроматида идет к одному полюсу, другая – к другому и превращается таким образом в хромосомы. В телофазе II образуются 4 гаплоидных ядра. В клетке происходит цитокинез, в результате которого из 2 -х предшествующих клеток получаются четыре. Каждая клетка содержит гаплоидный набор хромосом, структура которых состоит из одной хроматиды.
 Значение мейоза: I. Половое размножение: 1. В результате мейоза образуются четыре дочерние клетки, каждая из которых содержит половинное число хромосом по сравнению с родительской клеткой. 2. Этот процесс обеспечивает постоянство числа хромосом в ряду поколений, размножающихся половым путем организмов.
Значение мейоза: I. Половое размножение: 1. В результате мейоза образуются четыре дочерние клетки, каждая из которых содержит половинное число хромосом по сравнению с родительской клеткой. 2. Этот процесс обеспечивает постоянство числа хромосом в ряду поколений, размножающихся половым путем организмов.
 3. При данном типе такого оплодотворения для формирования нового организма необходимо оплодотворение, то есть слияние двух половых клеток (гамет), которые образуют зиготу, содержащую уже постоянное для каждого вида число хромосом, то есть диплоидный набор 2 n. 4. Если бы не происходила редукция (уменьшение) наследственного материала, то в каждом поколении число хромосом бы удваивалось.
3. При данном типе такого оплодотворения для формирования нового организма необходимо оплодотворение, то есть слияние двух половых клеток (гамет), которые образуют зиготу, содержащую уже постоянное для каждого вида число хромосом, то есть диплоидный набор 2 n. 4. Если бы не происходила редукция (уменьшение) наследственного материала, то в каждом поколении число хромосом бы удваивалось.
 II. Генетическая изменчивость: Мейоз создает возможности для возникновения в клетках новых генных комбинаций. Это ведет к изменению в фенотипе потомства, получаемого в результате слияния гамет.
II. Генетическая изменчивость: Мейоз создает возможности для возникновения в клетках новых генных комбинаций. Это ведет к изменению в фенотипе потомства, получаемого в результате слияния гамет.
 Механизмы мейоза, участвующие в создании изменчивости: 1. Уменьшение числа хромосом от диплоидного до гаплоидного, что сопровождается разделением аллелей, так что каждая гамета несет только один аллель по данному локусу ( месту);
Механизмы мейоза, участвующие в создании изменчивости: 1. Уменьшение числа хромосом от диплоидного до гаплоидного, что сопровождается разделением аллелей, так что каждая гамета несет только один аллель по данному локусу ( месту);
 2. Расположение бивалентов в экваториальной пластинке веретена в метафазе I и метафазе II определяется случайным образом. Последующее их разделение в анафазах I и II создает новые комбинации аллелей в гаметах. Этот процесс называется независимым распределением аллелей и лежит в основе второго закона Менделя.
2. Расположение бивалентов в экваториальной пластинке веретена в метафазе I и метафазе II определяется случайным образом. Последующее их разделение в анафазах I и II создает новые комбинации аллелей в гаметах. Этот процесс называется независимым распределением аллелей и лежит в основе второго закона Менделя.
 3. В результате образование хиазм между гомологичными хромосомами в профазе I часто происходит кроссинговер, ведущий к возникновению новых комбинаций аллелей в хромосомах половых клеток. Ранее существовавшие группы сцепления распадаются, возникают новые группы сцепления.
3. В результате образование хиазм между гомологичными хромосомами в профазе I часто происходит кроссинговер, ведущий к возникновению новых комбинаций аллелей в хромосомах половых клеток. Ранее существовавшие группы сцепления распадаются, возникают новые группы сцепления.
 4. В процессе мейоза I в формирующихся клетках с равной вероятностью оказываются отцовские и материнские хромосомы от предков организма, в котором происходит мейоз. Таким образом, происходит перемешивание генетической информации, полученной от предков.
4. В процессе мейоза I в формирующихся клетках с равной вероятностью оказываются отцовские и материнские хромосомы от предков организма, в котором происходит мейоз. Таким образом, происходит перемешивание генетической информации, полученной от предков.
. Мейоз I: 1, 2, 3. 4. 5") Схема мейоза (показана одна пара гомологичных хромосом). Мейоз I: 1, 2, 3. 4. 5 — профаза; 6 —метафаза; 7 — анафаза; 8 — телофаза; 9 — интеркинез. Мейоз II; 10 —метафаза; II —анафаза; 12 — дочерние клетки. Схема мейоза (показана одна пара гомологичных хромосом). Мейоз I: 1, 2, 3. 4. 5 — профаза; 6 —метафаза; 7 — анафаза; 8 — телофаза; 9 — интеркинез. Мейоз II;
Схема мейоза (показана одна пара гомологичных хромосом). Мейоз I: 1, 2, 3. 4. 5 — профаза; 6 —метафаза; 7 — анафаза; 8 — телофаза; 9 — интеркинез. Мейоз II; 10 —метафаза; II —анафаза; 12 — дочерние клетки. Схема мейоза (показана одна пара гомологичных хромосом). Мейоз I: 1, 2, 3. 4. 5 — профаза; 6 —метафаза; 7 — анафаза; 8 — телофаза; 9 — интеркинез. Мейоз II;
 ГАМЕТОГЕНЕЗ У ЧЕЛОВЕКА
ГАМЕТОГЕНЕЗ У ЧЕЛОВЕКА
 Первичные половые клетки выделяются из желточного мешка на 27 -й день развития и мигрирует на место формирования половых органов, на 46 -й день дифференцируются и становятся либо яичниками, либо семенниками. В этот период в мужских гонадах первичные клетки становятся сперматогониями, которые делятся путем митоза до периода половой зрелости.
Первичные половые клетки выделяются из желточного мешка на 27 -й день развития и мигрирует на место формирования половых органов, на 46 -й день дифференцируются и становятся либо яичниками, либо семенниками. В этот период в мужских гонадах первичные клетки становятся сперматогониями, которые делятся путем митоза до периода половой зрелости.
.") В 15 -16 лет сперматогонии вступают в мейоз и переходят в состояние сперматозоидов (спермиев). Процесс образования спермиев из сперматогоний занимает 70 дней.
В 15 -16 лет сперматогонии вступают в мейоз и переходят в состояние сперматозоидов (спермиев). Процесс образования спермиев из сперматогоний занимает 70 дней.
 проходят ряд последовательных митотических") Клетки зачаточного эпителия в мужских и женских половых железах (гонадах) проходят ряд последовательных митотических и мейотических делений, называемых гаметогенезом, в результате чего образуются зрелые мужские гаметы ( сперматогенез) и женские гаметы (оогенез).
Клетки зачаточного эпителия в мужских и женских половых железах (гонадах) проходят ряд последовательных митотических и мейотических делений, называемых гаметогенезом, в результате чего образуются зрелые мужские гаметы ( сперматогенез) и женские гаметы (оогенез).
 В обоих случаях процесс делится на три фазы – фазу размножения, фазу роста и фазу созревания. Фаза размножения включает многократные митотические процессы, в результате чего формируется множество сперматогоний. Затем каждая клетка растет и готовится к первому делению мейоза. Затем начинается фаза созревания, во время которой происходит первое и второе деление мейоза с последующей дифференцировкой гаплоидных клеток и формированием зрелых гамет.
В обоих случаях процесс делится на три фазы – фазу размножения, фазу роста и фазу созревания. Фаза размножения включает многократные митотические процессы, в результате чего формируется множество сперматогоний. Затем каждая клетка растет и готовится к первому делению мейоза. Затем начинается фаза созревания, во время которой происходит первое и второе деление мейоза с последующей дифференцировкой гаплоидных клеток и формированием зрелых гамет.
 В фазе размножения происходят многократные митотические деления, приводящие к образованию множества сперматогоний, проходящих период роста в порядке подготовки к первому делению мейоза и последующему цитогенезу.
В фазе размножения происходят многократные митотические деления, приводящие к образованию множества сперматогоний, проходящих период роста в порядке подготовки к первому делению мейоза и последующему цитогенезу.

 В процессе сперматогенеза сперматогонии увеличиваются в размере, превращаются в сперматоциты I порядка, содержат 46 удвоенных хромосом, затем вступают в первое деление мейоза, переходят в сперматоциты II порядка. Сперматоциты II порядка содержат по 2 хроматиды, в каждой из которых имеется по 23 хромосомы.
В процессе сперматогенеза сперматогонии увеличиваются в размере, превращаются в сперматоциты I порядка, содержат 46 удвоенных хромосом, затем вступают в первое деление мейоза, переходят в сперматоциты II порядка. Сперматоциты II порядка содержат по 2 хроматиды, в каждой из которых имеется по 23 хромосомы.
 После второго деления мейоза из сперматоцитов II порядка образуются сперматиды, содержащие по 23 хромосомы в единственной хроматиде. Сперматиды затем изменяются: меняется состояние клеточной оболочки, появляются шейка и хвост, формируется сперматозоид.
После второго деления мейоза из сперматоцитов II порядка образуются сперматиды, содержащие по 23 хромосомы в единственной хроматиде. Сперматиды затем изменяются: меняется состояние клеточной оболочки, появляются шейка и хвост, формируется сперматозоид.

 Образование и выделение гамет у мужчин начинается с наступления половой зрелости, продолжается в течение всей жизни
Образование и выделение гамет у мужчин начинается с наступления половой зрелости, продолжается в течение всей жизни
 У женщин процесс формирования половых клеток начинается после первичной половой дифференцировки плода, когда формируются в заложенных половых органах оогонии. Они претерпевают несколько митотических делений и со второго месяца внутриутробного развития до седьмого месяца у женского плода.
У женщин процесс формирования половых клеток начинается после первичной половой дифференцировки плода, когда формируются в заложенных половых органах оогонии. Они претерпевают несколько митотических делений и со второго месяца внутриутробного развития до седьмого месяца у женского плода.
 В этот период ряд оогоний вступает в первое деление мейоза и превращаются в ооциты I порядка. Деление ооцитов I порядка останавливается на стадии профазы I сразу после диплотены на седьмом месяце внутриутробного развития и не происходит до периода полового созревания.
В этот период ряд оогоний вступает в первое деление мейоза и превращаются в ооциты I порядка. Деление ооцитов I порядка останавливается на стадии профазы I сразу после диплотены на седьмом месяце внутриутробного развития и не происходит до периода полового созревания.
 Плод женского пола перед рождением содержит около 2 млн оогоний I порядка, но лишь 4 -5 часть из них достигает состояния оогоний II порядка и выходят из яичника (овуляция). У взрослой женщины овариальный цикл начинается с развития нескольких первичных фолликулов (содержащих ооциты I порядка) под воздействием фолликулостимулирующего гормона (вырабатывается передней долей гипофиза).
Плод женского пола перед рождением содержит около 2 млн оогоний I порядка, но лишь 4 -5 часть из них достигает состояния оогоний II порядка и выходят из яичника (овуляция). У взрослой женщины овариальный цикл начинается с развития нескольких первичных фолликулов (содержащих ооциты I порядка) под воздействием фолликулостимулирующего гормона (вырабатывается передней долей гипофиза).
.") После рождения девочки из этих фолликулов растет только один, а остальные разрушаются (атрезия фолликулов). В период половой зрелости каждый месяц ооцит I порядка заканчивает первое деление мейоза, после которого образуются две разных клетки: одна большая, которая содержит практически всю цитоплазму от ооцита I порядка. Она получила название ооцит II порядка.
После рождения девочки из этих фолликулов растет только один, а остальные разрушаются (атрезия фолликулов). В период половой зрелости каждый месяц ооцит I порядка заканчивает первое деление мейоза, после которого образуются две разных клетки: одна большая, которая содержит практически всю цитоплазму от ооцита I порядка. Она получила название ооцит II порядка.
 Помимо ооцита II порядка формируется маленькая клетка только с ядром с 23 хромосомами, которая получила название первое полярное тельце. Второе деление мейоза доходит до стадии метафазы, останавливается до тех пор, пока яйцеклетка не сольется со сперматозоидом. При оплодотворении ооцита II порядка совершает второе деление мейоза и образует крупную клетку – яйцо и второе полярное тельце.
Помимо ооцита II порядка формируется маленькая клетка только с ядром с 23 хромосомами, которая получила название первое полярное тельце. Второе деление мейоза доходит до стадии метафазы, останавливается до тех пор, пока яйцеклетка не сольется со сперматозоидом. При оплодотворении ооцита II порядка совершает второе деление мейоза и образует крупную клетку – яйцо и второе полярное тельце.
 В оогенезе мейоз II заканчивается в момент оплодотворения, после которого в течение нескольких часов мужские и женские гаплоидные наборы хромосом не сливаются, хотя окружаются сразу общей ядерной мембраной. После слияния хромосом формируется клетка с диплоидным набором хромосом, которая называется зигота, в результате митотического деления которой формируется бластоциста.
В оогенезе мейоз II заканчивается в момент оплодотворения, после которого в течение нескольких часов мужские и женские гаплоидные наборы хромосом не сливаются, хотя окружаются сразу общей ядерной мембраной. После слияния хромосом формируется клетка с диплоидным набором хромосом, которая называется зигота, в результате митотического деления которой формируется бластоциста.
 Через 8 дней после овуляции бластоциста погружается в стенку матки, наружные клетки бластоцисты формируют трофобласт. Формируется новый организм.
Через 8 дней после овуляции бластоциста погружается в стенку матки, наружные клетки бластоцисты формируют трофобласт. Формируется новый организм.

 Патология мейоза обычно является отражением сбоя распределения хромосом в гаметах. Отсюда выделяют простое, последовательное и двойное нерасхождение хромосом. При простом нерасхождении происходит неправильное распределение хромосом или в первом или во втором делении мейоза.
Патология мейоза обычно является отражением сбоя распределения хромосом в гаметах. Отсюда выделяют простое, последовательное и двойное нерасхождение хромосом. При простом нерасхождении происходит неправильное распределение хромосом или в первом или во втором делении мейоза.
 Если затрагивается первое деление мейоза, то все гаметы будут иметь патологический набор хромосом – анеуплодию. Если затрагивается второе деление мейоза, то изменяется количество хромосом только в какой то части гамет.
Если затрагивается первое деление мейоза, то все гаметы будут иметь патологический набор хромосом – анеуплодию. Если затрагивается второе деление мейоза, то изменяется количество хромосом только в какой то части гамет.
 Последовательное нерасхождение затрагивает оба деления мейоза. В этом случае нормальные гаметы не образуются. Если мейоз повреждается у обоих родителей, то формируется двойное нерасхождение. Кроме указанных могут быть: Первичное нарушение, когда нарушения происходят у людей с изначально нормальным набором хромосом.
Последовательное нерасхождение затрагивает оба деления мейоза. В этом случае нормальные гаметы не образуются. Если мейоз повреждается у обоих родителей, то формируется двойное нерасхождение. Кроме указанных могут быть: Первичное нарушение, когда нарушения происходят у людей с изначально нормальным набором хромосом.
 Вторичное нарушение происходит у людей с изначально патологическим набором хромосом. Третичное происходит у людей, являющихся носителями сбалансированной перестройки хромосом.
Вторичное нарушение происходит у людей с изначально патологическим набором хромосом. Третичное происходит у людей, являющихся носителями сбалансированной перестройки хромосом.
 Современные методы анализа хромосом: 1. Цитогенетический метод – микроскопическое изучение хромосом человека, для чего применяют методы культивирования изучаемых клеток с последующим окрашиванием хромосом (простое, дифференциальное).
Современные методы анализа хромосом: 1. Цитогенетический метод – микроскопическое изучение хромосом человека, для чего применяют методы культивирования изучаемых клеток с последующим окрашиванием хромосом (простое, дифференциальное).
 2. Метод генетики соматических клеток- культивирование отдельных клеток с получением из них клонов. Клоны при этом могут сливаться, давать гибридное потомство, легко подвергаются селекции на питательных средах.
2. Метод генетики соматических клеток- культивирование отдельных клеток с получением из них клонов. Клоны при этом могут сливаться, давать гибридное потомство, легко подвергаются селекции на питательных средах.
 3. Биохимический метод позволяет определять метаболиты, специфические для какого либо наследственного заболевания. Современные биохимические методы – электрофорез, хроматография, спектрография. Биохимические методы используются у больных с наследственным нарушением обмена веществ.
3. Биохимический метод позволяет определять метаболиты, специфические для какого либо наследственного заболевания. Современные биохимические методы – электрофорез, хроматография, спектрография. Биохимические методы используются у больных с наследственным нарушением обмена веществ.
 4. Молекулярно-генетические методы позволяют проводить анализ особенностей ДНК с помощью разрезания молекулы ДНК специальными биологическими «ножницами» – рестриктазами ( ферментами). В результате образуются фрагменты разного размера, которые специфичны для той или иной рестриктазы. Интересным и современным методом является полимеразная цепная реакция (ПЦР), которая позволяет обнаруживать и многократно копировать небольшие участки ДНК.
4. Молекулярно-генетические методы позволяют проводить анализ особенностей ДНК с помощью разрезания молекулы ДНК специальными биологическими «ножницами» – рестриктазами ( ферментами). В результате образуются фрагменты разного размера, которые специфичны для той или иной рестриктазы. Интересным и современным методом является полимеразная цепная реакция (ПЦР), которая позволяет обнаруживать и многократно копировать небольшие участки ДНК.