

Живая клетка.ppt
- Количество слайдов: 131
 ЖИВАЯ КЛЕТКА
ЖИВАЯ КЛЕТКА
 Иерархические уровни организации живой материи
Иерархические уровни организации живой материи
 КЛЕТКА • Клетка – элементарная структурная, функциональная и генетическая единица в составе всех живых организмов.
КЛЕТКА • Клетка – элементарная структурная, функциональная и генетическая единица в составе всех живых организмов.
 ВИДЫ КЛЕТОК В зависимости от способности синтезировать ДНК клетки подразделяют на 3 группы: 1. Статические — не делящиеся, ядросодержащие, необратимо дифференцированные клетки — нервные, мышечные, железистые, гранулоциты и макрофаги крови и тканей, жировые клетки. При увеличении нагрузки эти клетки гипертрофируются, а при чрезмерной нагрузке — погибают. 2. Лабильные — обновляющиеся, постоянно делящиеся клетки. К ним относятся стволовые клетки костного мозга, самоподдерживающая популяция клеток, способная дифференцироваться в разных направлениях, способна к пролиферации, имеет четко выраженный клеточный цикл развития. Тканевые клетки — самоподдерживающая популяция, способная дифференцироваться только в одном направлении — это клетки базального слоя эпидермиса, крипт кишечника и др. 3. Стабильные (покоящиеся) — способные к пролиферации клетки. В норме в базе покоя не синтезируют ДНК. При стимуляции начинают синтезировать ДНК и митотически делиться. Это клетки мерокринных желез — печени, почек, поджелудочной железы и хряща. Эти клетки способны к регенерации. Клетки, периодически входящие в митотический цикл только при воздействии индукторов-митогенов — лимфоциты и др.
ВИДЫ КЛЕТОК В зависимости от способности синтезировать ДНК клетки подразделяют на 3 группы: 1. Статические — не делящиеся, ядросодержащие, необратимо дифференцированные клетки — нервные, мышечные, железистые, гранулоциты и макрофаги крови и тканей, жировые клетки. При увеличении нагрузки эти клетки гипертрофируются, а при чрезмерной нагрузке — погибают. 2. Лабильные — обновляющиеся, постоянно делящиеся клетки. К ним относятся стволовые клетки костного мозга, самоподдерживающая популяция клеток, способная дифференцироваться в разных направлениях, способна к пролиферации, имеет четко выраженный клеточный цикл развития. Тканевые клетки — самоподдерживающая популяция, способная дифференцироваться только в одном направлении — это клетки базального слоя эпидермиса, крипт кишечника и др. 3. Стабильные (покоящиеся) — способные к пролиферации клетки. В норме в базе покоя не синтезируют ДНК. При стимуляции начинают синтезировать ДНК и митотически делиться. Это клетки мерокринных желез — печени, почек, поджелудочной железы и хряща. Эти клетки способны к регенерации. Клетки, периодически входящие в митотический цикл только при воздействии индукторов-митогенов — лимфоциты и др.
 ФОРМА КЛЕТОК
ФОРМА КЛЕТОК
 ФУНКЦИИ КЛЕТКИ 1. Специализация 2. Поддержание клеточного гомеостаза 3. Обмен информацией 4. Биотрансформирующая функция 5. Биологический синтез 6. Энергетический обмен 7. Хранение и передача наследственной информации
ФУНКЦИИ КЛЕТКИ 1. Специализация 2. Поддержание клеточного гомеостаза 3. Обмен информацией 4. Биотрансформирующая функция 5. Биологический синтез 6. Энергетический обмен 7. Хранение и передача наследственной информации
") ХИМИЧЕСКАЯ ОРГАНИЗАЦИЯ КЛЕТКИ 1. Неорганические вещества ( вода, кислоты, основания, положительные и отрицательные ионы) 2. Органические вещества (белки — 10 -20%, жиры или липиды — 1 -5%, углеводы — 0, 2 - 2%) 3. Нуклеиновые кислоты ( ДНК и РНК) 4. Ферменты, или энзимы
ХИМИЧЕСКАЯ ОРГАНИЗАЦИЯ КЛЕТКИ 1. Неорганические вещества ( вода, кислоты, основания, положительные и отрицательные ионы) 2. Органические вещества (белки — 10 -20%, жиры или липиды — 1 -5%, углеводы — 0, 2 - 2%) 3. Нуклеиновые кислоты ( ДНК и РНК) 4. Ферменты, или энзимы
 Компоненты клетки • клетки всех типов характеризуются сходством общей организации и строения важнейших компонентов. • каждая клетка эукариот состоит из двух основных компонентов – ядра и цитоплазмы
Компоненты клетки • клетки всех типов характеризуются сходством общей организации и строения важнейших компонентов. • каждая клетка эукариот состоит из двух основных компонентов – ядра и цитоплазмы
 Ядро • Ядро – важнейший компонент клетки, содержащий её генетический аппарат. • Функции ядра: • хранение генетической информации (в молекулах ДНК, находящихся в хромосомах); • реализация генетической информации, контролирующей различные процессы в клетке: транскрипция информационных, рибосомальных, транспортных РНК → синтетическая активность; апоптоз и т. д. ); • воспроизведение и передача генетической информации при делении клетки.
Ядро • Ядро – важнейший компонент клетки, содержащий её генетический аппарат. • Функции ядра: • хранение генетической информации (в молекулах ДНК, находящихся в хромосомах); • реализация генетической информации, контролирующей различные процессы в клетке: транскрипция информационных, рибосомальных, транспортных РНК → синтетическая активность; апоптоз и т. д. ); • воспроизведение и передача генетической информации при делении клетки.
 клетки выявляются следующие компоненты: ядерная") Компоненты ядра • • • В ядре неделящейся (интерфазной) клетки выявляются следующие компоненты: ядерная оболочка (кариолемма) хроматин ядрышко кариоплазма
Компоненты ядра • • • В ядре неделящейся (интерфазной) клетки выявляются следующие компоненты: ядерная оболочка (кариолемма) хроматин ядрышко кариоплазма
 на светооптическом уровне практически не определяется.") Ядерная оболочка • • Ядерная оболочка (кариолемма, нуклеолемма) на светооптическом уровне практически не определяется. Под электронном микроскопом обнаруживается, что она состоит из двух мембран – наружной и внутренней мембран, разделенных полостью шириной 15 -40 нм – перинуклеарной цистерной. Наружная мембрана составляет единое целое с мембранами гр. ЭПС: на её поверхности имеются рибосомы, а перинуклеарная цистерна сообщается с цистерной гр. ЭПС
Ядерная оболочка • • Ядерная оболочка (кариолемма, нуклеолемма) на светооптическом уровне практически не определяется. Под электронном микроскопом обнаруживается, что она состоит из двух мембран – наружной и внутренней мембран, разделенных полостью шириной 15 -40 нм – перинуклеарной цистерной. Наружная мембрана составляет единое целое с мембранами гр. ЭПС: на её поверхности имеются рибосомы, а перинуклеарная цистерна сообщается с цистерной гр. ЭПС
 • Внутренняя мембрана – гладкая, её интегральные белки связаны со слоем, состоящим из сети промежуточных филаментов (ламинов), - ламиной, или ядерной пластинкой. • Ламина играет большую роль в поддержании формы ядра, укладке хроматина и структурной организации поровых комплексов
• Внутренняя мембрана – гладкая, её интегральные белки связаны со слоем, состоящим из сети промежуточных филаментов (ламинов), - ламиной, или ядерной пластинкой. • Ламина играет большую роль в поддержании формы ядра, укладке хроматина и структурной организации поровых комплексов
 • В определенных точках наружная и внутренняя мембрана смыкаются, образуя ядерные поры. • Ядерная пора образована двумя параллельными кольцами диаметром 80 нм, содержащих по 8 белковых гранул, от которых к центру поры тянутся фибриллы, формирующие диафрагму толщиной около 5 нм. В середине диафрагмы лежит центральная гранула. Белковые гранулы ядерной поры структурно связаны с белками ядерной ламины. • Совокупность компонентов, входящих в состав ядерной поры, называется комплексом ядерной поры.
• В определенных точках наружная и внутренняя мембрана смыкаются, образуя ядерные поры. • Ядерная пора образована двумя параллельными кольцами диаметром 80 нм, содержащих по 8 белковых гранул, от которых к центру поры тянутся фибриллы, формирующие диафрагму толщиной около 5 нм. В середине диафрагмы лежит центральная гранула. Белковые гранулы ядерной поры структурно связаны с белками ядерной ламины. • Совокупность компонентов, входящих в состав ядерной поры, называется комплексом ядерной поры.
 • Ядерная оболочка клетки содержит 20004000 поровых комплексов. • Число поровых комплексов возрастает с увеличением функциональной активности: в клетках с высокой синтетической активностью ядерные поры занимают до 35% поверхности кариолеммы.
• Ядерная оболочка клетки содержит 20004000 поровых комплексов. • Число поровых комплексов возрастает с увеличением функциональной активности: в клетках с высокой синтетической активностью ядерные поры занимают до 35% поверхности кариолеммы.
 Комплекс ядерной поры • • Комплекс ядерной поры обеспечивает избирательный транспорт веществ между цитоплазмой и ядром. По каналу, образованному поровым комплексом, движутся мелкие водорастворимые молекулы и ионы; активно переносятся в ядро белки, синтезируемые в цитоплазме (белки с маркировкой в виде с особой последовательности аминокислот – NLS, распознаваемой рецепторами NLS в комплексе поры); из ядра в цитоплазму переносятся субъединицы рибосом.
Комплекс ядерной поры • • Комплекс ядерной поры обеспечивает избирательный транспорт веществ между цитоплазмой и ядром. По каналу, образованному поровым комплексом, движутся мелкие водорастворимые молекулы и ионы; активно переносятся в ядро белки, синтезируемые в цитоплазме (белки с маркировкой в виде с особой последовательности аминокислот – NLS, распознаваемой рецепторами NLS в комплексе поры); из ядра в цитоплазму переносятся субъединицы рибосом.
 клетке соответствует хромосомам и состоит из") • • • Хроматин в интерфазной (неделящейся) клетке соответствует хромосомам и состоит из комплекса ДНК и белка. Выраженность спирализации каждой из хромосом неодинакова по длине. Соответственно, различают два вида хроматина: эухроматин и гетерохроматин. Эухроматин соответствует участкам хромосом, которые деспирализованы и открыты для транскрипции. Эти участки не окрашиваются и не видны в световой микроскоп. Гетерохроматин соответствует конденсированным сегментам хромосом, что делает их недоступными для транскрипции. Гетерохроматин интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид мелких гранул и глыбок.
• • • Хроматин в интерфазной (неделящейся) клетке соответствует хромосомам и состоит из комплекса ДНК и белка. Выраженность спирализации каждой из хромосом неодинакова по длине. Соответственно, различают два вида хроматина: эухроматин и гетерохроматин. Эухроматин соответствует участкам хромосом, которые деспирализованы и открыты для транскрипции. Эти участки не окрашиваются и не видны в световой микроскоп. Гетерохроматин соответствует конденсированным сегментам хромосом, что делает их недоступными для транскрипции. Гетерохроматин интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид мелких гранул и глыбок.
 Структура ядер
Структура ядер
 эухроматин-гетерохроматин • по соотношению эу- и гетерохроматина в ядре можно оценить активность процессов транскрипции, и, следовательно, синтетической функции клетки. • При её повышении это соотношение изменяется в пользу эухроматина, при снижении – нарастает содержание гетерохроматина. • Соотношение эухроматин-гетерохроматин может, например, служить основой для дифференциальной диагностики доброкачественных и злокачественных опухолевых клеток. • При полном подавлении функции ядра в поврежденных и гибнущих клетках, оно уменьшается в размерах и содержит только гетерохроматин. Такое явление называется кариопикнозом.
эухроматин-гетерохроматин • по соотношению эу- и гетерохроматина в ядре можно оценить активность процессов транскрипции, и, следовательно, синтетической функции клетки. • При её повышении это соотношение изменяется в пользу эухроматина, при снижении – нарастает содержание гетерохроматина. • Соотношение эухроматин-гетерохроматин может, например, служить основой для дифференциальной диагностики доброкачественных и злокачественных опухолевых клеток. • При полном подавлении функции ядра в поврежденных и гибнущих клетках, оно уменьшается в размерах и содержит только гетерохроматин. Такое явление называется кариопикнозом.
 – скопление гетерохроматина, соответствующее одной из пары Х-хромосом,") • Половой хроматин (тельце Барра) – скопление гетерохроматина, соответствующее одной из пары Х-хромосом, которая в интерфазе плотно скручена и неактивна. • • • Выявление полового хроматина используется как диагностический тест для определения генетического женского пола, что существенно при изучении генетических аномалий и, особенно, в спортивной медицине. Обычно анализируют эпителиальные клетки слизистой оболочки полости рта, где, как и в большинстве других клеток, половой хроматин выявляется как крупная глыбка гетерохроматина, лежащая рядом с ядерной оболочки. В нейтрофильных лейкоцитах крови половой хроматин имеет вид маленькой добавочной дольки ядра ( «барабанной палочки» ).
• Половой хроматин (тельце Барра) – скопление гетерохроматина, соответствующее одной из пары Х-хромосом, которая в интерфазе плотно скручена и неактивна. • • • Выявление полового хроматина используется как диагностический тест для определения генетического женского пола, что существенно при изучении генетических аномалий и, особенно, в спортивной медицине. Обычно анализируют эпителиальные клетки слизистой оболочки полости рта, где, как и в большинстве других клеток, половой хроматин выявляется как крупная глыбка гетерохроматина, лежащая рядом с ядерной оболочки. В нейтрофильных лейкоцитах крови половой хроматин имеет вид маленькой добавочной дольки ядра ( «барабанной палочки» ).
 ДНК,") Упаковка хроматина в ядре • В деконденсированном состоянии длина одной молекулы (двойной спирали) ДНК, образующей одну хромосому, составляет около 5 см, а общая длина молекул ДНК в ядре – более 2 м. Такие длинные нити ДНК компактно и упорядоченно упакованы в ядре диаметром всего 5 -10 мкм. • Компактная упаковка молекул ДНК осуществляется благодаря связи ДНК со специальными основными белками – гистонами.
Упаковка хроматина в ядре • В деконденсированном состоянии длина одной молекулы (двойной спирали) ДНК, образующей одну хромосому, составляет около 5 см, а общая длина молекул ДНК в ядре – более 2 м. Такие длинные нити ДНК компактно и упорядоченно упакованы в ядре диаметром всего 5 -10 мкм. • Компактная упаковка молекул ДНК осуществляется благодаря связи ДНК со специальными основными белками – гистонами.
 Упаковка хроматина • • • Начальный уровень упаковки хроматина – нуклеосома с диаметром 11 нм. Нуклеосома состоит из блока, образованного комплексом из 8 молекул гистонов, на который намотана двойная нить ДНК (цепочка из 166 пар нуклеотидов). Нуклеосомы разделены короткими участками свободной ДНК (48 пар оснований). Нуклеосомная нить имеет вид нитки с бусинами, где каждая бусина – нуклеосома. Второй уровень упаковки также обусловлен гистонами и приводит к скручиванию нуклеосомной нити (виток из 6 нуклеосом) с формированием хроматиновой фибриллы диаметром 30 нм. Хроматиновые фибриллы образуют петли диаметром 300 нм. При делении клетки в результате еще более компактной укладки и сверхспирализации ДНК появляются хромосомы (диаметр 700 нм), видимые под световым микроскопом. Компактная упаковка ДНК в ядре обеспечивает упорядоченное расположение очень длинных молекул ДНК в небольшом объеме ядра, а также функциональный контроль активности генов.
Упаковка хроматина • • • Начальный уровень упаковки хроматина – нуклеосома с диаметром 11 нм. Нуклеосома состоит из блока, образованного комплексом из 8 молекул гистонов, на который намотана двойная нить ДНК (цепочка из 166 пар нуклеотидов). Нуклеосомы разделены короткими участками свободной ДНК (48 пар оснований). Нуклеосомная нить имеет вид нитки с бусинами, где каждая бусина – нуклеосома. Второй уровень упаковки также обусловлен гистонами и приводит к скручиванию нуклеосомной нити (виток из 6 нуклеосом) с формированием хроматиновой фибриллы диаметром 30 нм. Хроматиновые фибриллы образуют петли диаметром 300 нм. При делении клетки в результате еще более компактной укладки и сверхспирализации ДНК появляются хромосомы (диаметр 700 нм), видимые под световым микроскопом. Компактная упаковка ДНК в ядре обеспечивает упорядоченное расположение очень длинных молекул ДНК в небольшом объеме ядра, а также функциональный контроль активности генов.
 Ядрышко • Ядрышко выявляется в интерфазном ядре на светооптическом уровне как мелкая (~ 1 мкм в диаметре), плотная сферическая структура, интенсивно окрашивающееся основными красителями
Ядрышко • Ядрышко выявляется в интерфазном ядре на светооптическом уровне как мелкая (~ 1 мкм в диаметре), плотная сферическая структура, интенсивно окрашивающееся основными красителями
 Ядрышко • Ядрышко образовано специализированными участками хромосом – ядрышковыми организаторами, на которых происходит синтез рибосомальной РНК, а также её сборка в предшественники рибосомальных субъединиц.
Ядрышко • Ядрышко образовано специализированными участками хромосом – ядрышковыми организаторами, на которых происходит синтез рибосомальной РНК, а также её сборка в предшественники рибосомальных субъединиц.
 Компоненты ядрышка • • Аморфный компонент, слабо окрашиваемый, представляет собой участки расположения ядрышковых организаторов: крупные петли ДНК, активно участвующих в транскрипции рибосомальной РНК; Фибриллярный компонент состоит из множества нитей диаметром 5 -8 нм, преимущественно во внутренней части ядрышка, и представляет собой длинные молекулы р. РНК (первичные транскрипты); Гранулярный компонент образован скоплением плотных мелких гранулярных частиц, представляющие собой зреющие субъединицы рибосом. Рибосомальные субъединицы образуется из р. РНК, синтезированной в ядрышке, и белков, синтезированных в цитоплазме. Фибриллярный и гранулярный компоненты ядрышка образуют ядрышковую нить – нуклеолонему, которая образует петлистую сеть, выделяющуюся большой плотностью на фоне менее плотного ядерного матрикса
Компоненты ядрышка • • Аморфный компонент, слабо окрашиваемый, представляет собой участки расположения ядрышковых организаторов: крупные петли ДНК, активно участвующих в транскрипции рибосомальной РНК; Фибриллярный компонент состоит из множества нитей диаметром 5 -8 нм, преимущественно во внутренней части ядрышка, и представляет собой длинные молекулы р. РНК (первичные транскрипты); Гранулярный компонент образован скоплением плотных мелких гранулярных частиц, представляющие собой зреющие субъединицы рибосом. Рибосомальные субъединицы образуется из р. РНК, синтезированной в ядрышке, и белков, синтезированных в цитоплазме. Фибриллярный и гранулярный компоненты ядрышка образуют ядрышковую нить – нуклеолонему, которая образует петлистую сеть, выделяющуюся большой плотностью на фоне менее плотного ядерного матрикса
 Ядрышко • Размеры и объем ядрышек увеличиваются при повышении функциональной активности клетки. Особенно крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также клеток быстрорастущих злокачественных опухолей. • Ядрышко исчезает в профазе митоза, в результате инактивации рибосомных генов при конденсации соответствующих хромосом, и вновь формируется в поздней телофазе.
Ядрышко • Размеры и объем ядрышек увеличиваются при повышении функциональной активности клетки. Особенно крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также клеток быстрорастущих злокачественных опухолей. • Ядрышко исчезает в профазе митоза, в результате инактивации рибосомных генов при конденсации соответствующих хромосом, и вновь формируется в поздней телофазе.
 Ядерный матрикс • Ядерный матрикс – компонент ядра, в котором располагаются хроматин и ядрышко. Ядерный матрикс образован кариоплазмой и кариоскелетом. • Кариоплазма – жидкий компонент ядра, содержащий РНК, ионы, ферменты, метаболиты, растворенные в воде. • Кариоскелет состоит из ламины и других фибриллярных белков.
Ядерный матрикс • Ядерный матрикс – компонент ядра, в котором располагаются хроматин и ядрышко. Ядерный матрикс образован кариоплазмой и кариоскелетом. • Кариоплазма – жидкий компонент ядра, содержащий РНК, ионы, ферменты, метаболиты, растворенные в воде. • Кариоскелет состоит из ламины и других фибриллярных белков.
 Цитоплазма • Цитоплазма отделена от внешней среды плазматической мембраной и содержит органеллы и включения, погруженные в клеточный матрикс (цитозоль, гиалоплазма).
Цитоплазма • Цитоплазма отделена от внешней среды плазматической мембраной и содержит органеллы и включения, погруженные в клеточный матрикс (цитозоль, гиалоплазма).
 • Органеллы – постоянные компоненты цитоплазмы, имеющие характерную структуру и специализированные на выполнении определенных функций в клетке. • Включения – непостоянные компоненты цитоплазмы, образованные в результате накопления продуктов метаболизма клеток.
• Органеллы – постоянные компоненты цитоплазмы, имеющие характерную структуру и специализированные на выполнении определенных функций в клетке. • Включения – непостоянные компоненты цитоплазмы, образованные в результате накопления продуктов метаболизма клеток.
 Органеллы • Органеллы – постоянно присутствующие в цитоплазме структуры, имеющие определенное строение и специализированные на выполнении определенных функций в клетке. • Они подразделяются на органеллы общего значения и специальные органеллы.
Органеллы • Органеллы – постоянно присутствующие в цитоплазме структуры, имеющие определенное строение и специализированные на выполнении определенных функций в клетке. • Они подразделяются на органеллы общего значения и специальные органеллы.
 Органеллы общего значения • Органеллы общего значения имеются во всех клетках и необходимы для обеспечения их жизнедеятельности. К ним относятся: • митохондрии, • рибосомы • эндоплазматическая сеть (ЭПС), • комплекс Гольджи • лизосомы • пероксисомы, • клеточный центр • компоненты цитоскелета.
Органеллы общего значения • Органеллы общего значения имеются во всех клетках и необходимы для обеспечения их жизнедеятельности. К ним относятся: • митохондрии, • рибосомы • эндоплазматическая сеть (ЭПС), • комплекс Гольджи • лизосомы • пероксисомы, • клеточный центр • компоненты цитоскелета.
 Специальные органеллы • • • Специальные органеллы содержатся лишь в некоторых специализированных клетках, где они обеспечивают выполнение специальных функций. К специальным органеллам относят реснички, жгутики, миофибриллы, акросома. Все специальные органеллы образуются при развитии клетки как производные органелл общего значения, так, например, акросома спермия является производным комплекса Гольджи, реснички и жгутики – микротрубочек цитоскелета и т. д
Специальные органеллы • • • Специальные органеллы содержатся лишь в некоторых специализированных клетках, где они обеспечивают выполнение специальных функций. К специальным органеллам относят реснички, жгутики, миофибриллы, акросома. Все специальные органеллы образуются при развитии клетки как производные органелл общего значения, так, например, акросома спермия является производным комплекса Гольджи, реснички и жгутики – микротрубочек цитоскелета и т. д
 Органеллы • В состав многих органелл входит элементарная биологическая мембрана, поэтому органеллы подразделяются также на мембранные и немембранные. • Мембранные органеллы: митохондрии, ЭПС, комплекс Гольджи, лизосомы, пероксисомы; • Немембранные органеллы: рибосомы, клеточный центр, компоненты цитоскелета, микроворсинки, реснички, жгутики
Органеллы • В состав многих органелл входит элементарная биологическая мембрана, поэтому органеллы подразделяются также на мембранные и немембранные. • Мембранные органеллы: митохондрии, ЭПС, комплекс Гольджи, лизосомы, пероксисомы; • Немембранные органеллы: рибосомы, клеточный центр, компоненты цитоскелета, микроворсинки, реснички, жгутики
 Внутриклеточные мембраны • Элементарная биологическая мембрана, входящая в состав клеточных органелл, по своему строению представляет собой бислой липидов со встроенными белками и сходна со строением плазмолеммы, но не идентична ей. • Толщина мембран внутриклеточных органелл несколько меньше (6 -7, 5 нм). • Мембраны различных органелл существенно различаются по своим функциональным свойствам, благодаря присутствию разных структурных белков, формирующих трансмембранные каналы или насосы, ферментов, рецепторов, а также липидов
Внутриклеточные мембраны • Элементарная биологическая мембрана, входящая в состав клеточных органелл, по своему строению представляет собой бислой липидов со встроенными белками и сходна со строением плазмолеммы, но не идентична ей. • Толщина мембран внутриклеточных органелл несколько меньше (6 -7, 5 нм). • Мембраны различных органелл существенно различаются по своим функциональным свойствам, благодаря присутствию разных структурных белков, формирующих трансмембранные каналы или насосы, ферментов, рецепторов, а также липидов
 СИНТЕТИЧЕСКИЙ АППАРАТ КЛЕТКИ • Синтетический аппарат клеток включает органеллы, участвующие в синтезе различных веществ. • К таким органеллам относятся рибосомы, эндоплазматическая сеть и комплекс Гольджи. • Деятельность синтетического аппарата клетки контролируется активностью генов, локализованных в ядре.
СИНТЕТИЧЕСКИЙ АППАРАТ КЛЕТКИ • Синтетический аппарат клеток включает органеллы, участвующие в синтезе различных веществ. • К таким органеллам относятся рибосомы, эндоплазматическая сеть и комплекс Гольджи. • Деятельность синтетического аппарата клетки контролируется активностью генов, локализованных в ядре.
 Рибосомы • Рибосомы – мелкие, плотные немембранные органеллы, диаметром 15 -30 нм. • Функция рибосом – синтез белка путем соединения аминокислот в полипептидные цепочки • Каждая рибосома состоит из двух субъединиц: большой и малой. • Субъединицы образованы рибосомальными РНК (р. РНК) и особыми белками (около 80 видов). Соотношение р. РНК и белков равно 1: 1.
Рибосомы • Рибосомы – мелкие, плотные немембранные органеллы, диаметром 15 -30 нм. • Функция рибосом – синтез белка путем соединения аминокислот в полипептидные цепочки • Каждая рибосома состоит из двух субъединиц: большой и малой. • Субъединицы образованы рибосомальными РНК (р. РНК) и особыми белками (около 80 видов). Соотношение р. РНК и белков равно 1: 1.
 Рибосомы • Субъединицы собираются в ядре из р. РНК, которая образуется в ядрышке, и белков, которые синтезируются в цитоплазме и поступают в ядро. Затем субъединицы рибосом через ядерные поры перемещаются в цитоплазму, где они участвуют в синтезе белка.
Рибосомы • Субъединицы собираются в ядре из р. РНК, которая образуется в ядрышке, и белков, которые синтезируются в цитоплазме и поступают в ядро. Затем субъединицы рибосом через ядерные поры перемещаются в цитоплазму, где они участвуют в синтезе белка.
 • • Рибосомы могут встречаться в цитоплазме как отдельные гранулы (функционально неактивные, не транслирующие рибосомы), так и в форме скоплений – полирибосом (полисом) – активные рибосомы. Отдельные рибосомы полисом удерживаются вместе нитью информационной РНК. Информация, переносимая и. РНК, кодирует последовательность аминокислот в белке соответствующей последовательностью нуклеотидов. Рибосомы транслируют эту генетическую информацию в последовательность аминокислот в ходе белкового синтеза.
• • Рибосомы могут встречаться в цитоплазме как отдельные гранулы (функционально неактивные, не транслирующие рибосомы), так и в форме скоплений – полирибосом (полисом) – активные рибосомы. Отдельные рибосомы полисом удерживаются вместе нитью информационной РНК. Информация, переносимая и. РНК, кодирует последовательность аминокислот в белке соответствующей последовательностью нуклеотидов. Рибосомы транслируют эту генетическую информацию в последовательность аминокислот в ходе белкового синтеза.
 • • • Полисомы могут свободно располагаться в гиалоплазме, или быть прикрепленными к мембранам эндоплазматической сети (ЭПС). При этом белки, которые синтезируются на свободных полисомах, остаются в гиалоплазме и далее используются самой клеткой. Полисомы, которые своими большими субъединицами прикреплены к мембранам ЭПС, синтезируют белки, накапливающиеся в просвете цистерн ЭПС. В дальнейшем эти белки либо выводятся из клетки (например, пищеварительные ферменты, гормоны), либо остаются в клетке в структурах, ограниченных мембраной (например, лизосомы с набором лизосомальных ферментов)
• • • Полисомы могут свободно располагаться в гиалоплазме, или быть прикрепленными к мембранам эндоплазматической сети (ЭПС). При этом белки, которые синтезируются на свободных полисомах, остаются в гиалоплазме и далее используются самой клеткой. Полисомы, которые своими большими субъединицами прикреплены к мембранам ЭПС, синтезируют белки, накапливающиеся в просвете цистерн ЭПС. В дальнейшем эти белки либо выводятся из клетки (например, пищеварительные ферменты, гормоны), либо остаются в клетке в структурах, ограниченных мембраной (например, лизосомы с набором лизосомальных ферментов)
 Рибосомы • Рибосомы, в связи с наличием р. РНК, интенсивно окрашиваются основными красителями (гематоксилин, метиленовый синий). Присутствие значительного числа рибосом в цитоплазме клеток, активно синтезирующих белок, придает ей базофилию на светооптическом уровне.
Рибосомы • Рибосомы, в связи с наличием р. РНК, интенсивно окрашиваются основными красителями (гематоксилин, метиленовый синий). Присутствие значительного числа рибосом в цитоплазме клеток, активно синтезирующих белок, придает ей базофилию на светооптическом уровне.
 • • • ЭПС – система уплощенных, трубчатых, везикулярных структур, ограниченных") ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ (ЭПС) • • • ЭПС – система уплощенных, трубчатых, везикулярных структур, ограниченных мембраной. Название обусловлено тем, что её многочисленные элементы (цистерны, трубочки, пузырьки) образуют единую, непрерывную трехмерную сеть. Степень развития ЭПС варьирует в различных клетках, и даже в разных участках одной и той же клетки, и зависит от функциональной активности клеток. Различают две разновидности ЭПС: гранулярную ЭПС (гр. ЭПС) и гладкую, или агранулярную ЭПС (а. ЭПС), которые связаны между собой в переходной области.
ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ (ЭПС) • • • ЭПС – система уплощенных, трубчатых, везикулярных структур, ограниченных мембраной. Название обусловлено тем, что её многочисленные элементы (цистерны, трубочки, пузырьки) образуют единую, непрерывную трехмерную сеть. Степень развития ЭПС варьирует в различных клетках, и даже в разных участках одной и той же клетки, и зависит от функциональной активности клеток. Различают две разновидности ЭПС: гранулярную ЭПС (гр. ЭПС) и гладкую, или агранулярную ЭПС (а. ЭПС), которые связаны между собой в переходной области.
 • • Гранулярная ЭПС образована мембранными трубочками и уплощенными цистернами, на наружной (обращенной в сторону гиалоплазмы) поверхности которых расположены рибосомы. Прикрепление рибосом происходит благодаря интегральным рецепторным белкам мембран гр. ЭПС – рибофоринам. Эти же белки формируют гидрофобные каналы в мембране гр. ЭПС для проникновения синтезированной белковой цепочки в просвет цистерн.
• • Гранулярная ЭПС образована мембранными трубочками и уплощенными цистернами, на наружной (обращенной в сторону гиалоплазмы) поверхности которых расположены рибосомы. Прикрепление рибосом происходит благодаря интегральным рецепторным белкам мембран гр. ЭПС – рибофоринам. Эти же белки формируют гидрофобные каналы в мембране гр. ЭПС для проникновения синтезированной белковой цепочки в просвет цистерн.
 вновь синтезированных белковых молекул от") гр. ЭПС • Основная функция гр. ЭПС: сегрегация (отделение) вновь синтезированных белковых молекул от гиалоплазмы. • Таким образом, гр. ЭПС обеспечивает: • 1) биосинтез белков, предназначенных для экспорта из клетки; • 2) биосинтез ферментов лизосом • 2) биосинтез мембранных белков. • Белковые молекулы накапливаются внутри просвета цистерн, приобретают вторичную и третичную структуру, а также подвергаются начальным посттрансляционным изменениям – гидроксилированию, сульфатированию, фосфорилированию и гликозилированию (присоединение к белкам олигосахаридов с образованием гликопротеинов).
гр. ЭПС • Основная функция гр. ЭПС: сегрегация (отделение) вновь синтезированных белковых молекул от гиалоплазмы. • Таким образом, гр. ЭПС обеспечивает: • 1) биосинтез белков, предназначенных для экспорта из клетки; • 2) биосинтез ферментов лизосом • 2) биосинтез мембранных белков. • Белковые молекулы накапливаются внутри просвета цистерн, приобретают вторичную и третичную структуру, а также подвергаются начальным посттрансляционным изменениям – гидроксилированию, сульфатированию, фосфорилированию и гликозилированию (присоединение к белкам олигосахаридов с образованием гликопротеинов).
 гр. ЭПС • • • Гр. ЭПС присутствует во всех клетках, но наиболее развита в клетках, специализирующихся на белковом синтезе: в эпителиальных клетках поджелудочной железы, вырабатывающих пищеварительные ферменты; в фибробластах соединительной ткани, синтезирующих коллаген; в плазматических клетках, продуцирующих иммуноглобулины. В этих клетках элементы гр. ЭПС образуют параллельные скопления цистерн; при этом просвет цистерн часто расширен. Для всех этих клеток характерна выраженная базофилия цитоплазмы в области расположения элементов гр. ЭПС.
гр. ЭПС • • • Гр. ЭПС присутствует во всех клетках, но наиболее развита в клетках, специализирующихся на белковом синтезе: в эпителиальных клетках поджелудочной железы, вырабатывающих пищеварительные ферменты; в фибробластах соединительной ткани, синтезирующих коллаген; в плазматических клетках, продуцирующих иммуноглобулины. В этих клетках элементы гр. ЭПС образуют параллельные скопления цистерн; при этом просвет цистерн часто расширен. Для всех этих клеток характерна выраженная базофилия цитоплазмы в области расположения элементов гр. ЭПС.
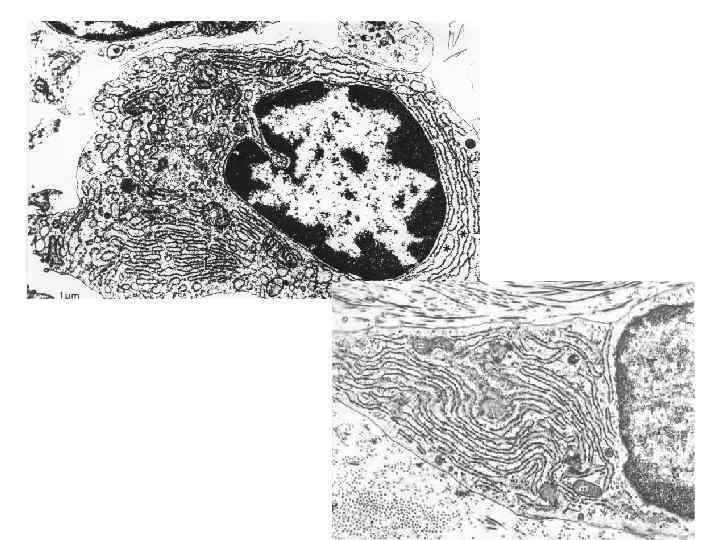
 Агранулярная ЭПС • агр. ЭПС представляет собой трехмерную сеть мембранных трубочек, канальцев, пузырьков, на поверхности которых рибосомы отсутствуют.
Агранулярная ЭПС • агр. ЭПС представляет собой трехмерную сеть мембранных трубочек, канальцев, пузырьков, на поверхности которых рибосомы отсутствуют.
 участие в синтезе липидов, в том числе мембранных, холестерина") Функции а. ЭПС • 1) участие в синтезе липидов, в том числе мембранных, холестерина и стероидов; • 2) метаболизм гликогена • 3) нейтрализация и детоксикация эндогенных и экзогенных токсичных веществ; • 4) накопление ионов Са (особенно в специализированной виде а. ЭПС – саркоплазматической сети мышечных клеток).
Функции а. ЭПС • 1) участие в синтезе липидов, в том числе мембранных, холестерина и стероидов; • 2) метаболизм гликогена • 3) нейтрализация и детоксикация эндогенных и экзогенных токсичных веществ; • 4) накопление ионов Са (особенно в специализированной виде а. ЭПС – саркоплазматической сети мышечных клеток).
 • агр. ЭПС хорошо развита: • в клетках, активно продуцирующих стероидные гормоны – клетки коркового вещества надпочечников, интерстициальные гландулоциты яичка, клетки желтого тела яичника. • в клетках печени, где её ферменты участвуют в метаболизме гликогена, а также в процессах, которые обеспечивают нейтрализацию и детоксикацию эндогенных биологически активных веществ (гормонов) и экзогенных вредных веществ (алкоголя, лекарственных веществ и др. )
• агр. ЭПС хорошо развита: • в клетках, активно продуцирующих стероидные гормоны – клетки коркового вещества надпочечников, интерстициальные гландулоциты яичка, клетки желтого тела яичника. • в клетках печени, где её ферменты участвуют в метаболизме гликогена, а также в процессах, которые обеспечивают нейтрализацию и детоксикацию эндогенных биологически активных веществ (гормонов) и экзогенных вредных веществ (алкоголя, лекарственных веществ и др. )
") КОМПЛЕКС ГОЛЬДЖИ • Комплекс Гольджи – мембранная органелла, образованная тремя основными элементами: • 1) скопления уплощенных цистерн; • 2) мелкие пузырьки; • 3) конденсирующие вакуоли. • Комплекс этих элементов называется диктиосомой.
КОМПЛЕКС ГОЛЬДЖИ • Комплекс Гольджи – мембранная органелла, образованная тремя основными элементами: • 1) скопления уплощенных цистерн; • 2) мелкие пузырьки; • 3) конденсирующие вакуоли. • Комплекс этих элементов называется диктиосомой.
 Комплекс Гольджи • • Цистерны имеют вид изогнутых дисков с несколько расширенными периферическими отделами. Цистерны образуют группу в виде стопки из 3 -30 элементов. Выпуклая сторона этой группы обращена обычно к ядру, вогнутая – к плазмолемме. От периферических расширений цистерн отщепляются пузырьки и вакуоли. Пузырьки – мелкие (диаметр 40 -80 нм), окруженные мембраной сферические элементы с содержимым умеренной электронной плотности. Вакуоли – крупные (диаметр 0. 1 -1. 0 мкм), сферические образования, отделяющиеся от зрелой поверхности комплекса Гольджи в некоторых железистых клетках. Вакуоли содержат секреторный продукт, находящийся в процессе конденсации.
Комплекс Гольджи • • Цистерны имеют вид изогнутых дисков с несколько расширенными периферическими отделами. Цистерны образуют группу в виде стопки из 3 -30 элементов. Выпуклая сторона этой группы обращена обычно к ядру, вогнутая – к плазмолемме. От периферических расширений цистерн отщепляются пузырьки и вакуоли. Пузырьки – мелкие (диаметр 40 -80 нм), окруженные мембраной сферические элементы с содержимым умеренной электронной плотности. Вакуоли – крупные (диаметр 0. 1 -1. 0 мкм), сферические образования, отделяющиеся от зрелой поверхности комплекса Гольджи в некоторых железистых клетках. Вакуоли содержат секреторный продукт, находящийся в процессе конденсации.
 Комплекс Гольджи • Комплекс Гольджи обладает полярностью: • в каждой диктиосоме выделяют две поверхности: формирующуюся (незрелую, или цисповерхность) и зрелую (транс-поверхность).
Комплекс Гольджи • Комплекс Гольджи обладает полярностью: • в каждой диктиосоме выделяют две поверхности: формирующуюся (незрелую, или цисповерхность) и зрелую (транс-поверхность).
 Комплекс Гольджи • Цис-поверхность выпуклой формы обращена в сторону ЭПС и связана с ней системой мелких транспортных пузырьков, отщепляющих от ЭПС. • Таким образом, белки в транспортных пузырьках проникают через цисповерхность.
Комплекс Гольджи • Цис-поверхность выпуклой формы обращена в сторону ЭПС и связана с ней системой мелких транспортных пузырьков, отщепляющих от ЭПС. • Таким образом, белки в транспортных пузырьках проникают через цисповерхность.
 Комплекс Гольджи • Каждая группа медиальных цистерн внутри стопки отличается особым составом ферментов, и для каждой группы характерны свои реакции обработки белков. • Обработанные вещества выходят в вакуолях с вогнутой транс-поверхности.
Комплекс Гольджи • Каждая группа медиальных цистерн внутри стопки отличается особым составом ферментов, и для каждой группы характерны свои реакции обработки белков. • Обработанные вещества выходят в вакуолях с вогнутой транс-поверхности.
; обработка белковых молекул") Функции комплекса Гольджи • • синтез полисахаридов и гликопротеинов (гликокаликса, слизи); обработка белковых молекул (терминальное гликозилирование – включение углеводных компонентов; фосфорилирование – добавление фосфатных групп; ацилирование – добавление жирных кислот; сульфатирование – добавление сульфатных остатков и т. д. ; сортировка белков на трансповерхности; упаковка секреторных продуктов в мембранные структуры
Функции комплекса Гольджи • • синтез полисахаридов и гликопротеинов (гликокаликса, слизи); обработка белковых молекул (терминальное гликозилирование – включение углеводных компонентов; фосфорилирование – добавление фосфатных групп; ацилирование – добавление жирных кислот; сульфатирование – добавление сульфатных остатков и т. д. ; сортировка белков на трансповерхности; упаковка секреторных продуктов в мембранные структуры
 • Секреторные продукты, обработанные в комплексе Гольджи, оказываются далее в секреторных гранулах, которые : выделяются путем экзоцитоза или остаются в клетке в виде: • в первичных лизосомах • в окаймленных пузырьках, в которых интегральные белки транспортируются в плазмолемму. • специфических гранул зернистых лейкоцитов
• Секреторные продукты, обработанные в комплексе Гольджи, оказываются далее в секреторных гранулах, которые : выделяются путем экзоцитоза или остаются в клетке в виде: • в первичных лизосомах • в окаймленных пузырьках, в которых интегральные белки транспортируются в плазмолемму. • специфических гранул зернистых лейкоцитов
 МИТОХОНДРИИ • Митохондрии – мембранные органеллы, присутствующих во всех эукариотических клетках, и представляющие собой энергетический аппарат клетки.
МИТОХОНДРИИ • Митохондрии – мембранные органеллы, присутствующих во всех эукариотических клетках, и представляющие собой энергетический аппарат клетки.
 Функции митохондрий: • основная – обеспечение клетки легко доступной энергией, которая образуется благодаря окислению метаболитов, и запасается частично в виде высоко-энергетических фосфатных связей АТФ; • участие в биосинтезе стероидов; • участие в окислении жирных кислот.
Функции митохондрий: • основная – обеспечение клетки легко доступной энергией, которая образуется благодаря окислению метаболитов, и запасается частично в виде высоко-энергетических фосфатных связей АТФ; • участие в биосинтезе стероидов; • участие в окислении жирных кислот.
 Митохондрии • Митохондрии могут иметь эллиптическую, палочковидную или нитевидную форму. Их размеры составляют 0. 2 -2 мкм в ширину и до 10 мкм в длину. • Число митохондрий в разных клетках и их распределение в пределах клетки варьирует.
Митохондрии • Митохондрии могут иметь эллиптическую, палочковидную или нитевидную форму. Их размеры составляют 0. 2 -2 мкм в ширину и до 10 мкм в длину. • Число митохондрий в разных клетках и их распределение в пределах клетки варьирует.
 Митохондрии • • Много митохондрий встречается в клетках с активным метаболизмом, требующим высоких энергетических затрат: кардиомиоцитах, клетках почечных канальцев, париетальных клетках желез дна желудка и т. д. В цитоплазме митохондрии могут распределяться диффузно, но имеют тенденцию аккумулироваться в участках максимального потребления энергии, например в апикальной части реснитчатых клеток, в связующем отделе сперматозоидов, или вблизи ионных насосов (зона базальной исчерченности в проксимальных канальцах почек, в исчерченных протоках слюнных желез и др. ).
Митохондрии • • Много митохондрий встречается в клетках с активным метаболизмом, требующим высоких энергетических затрат: кардиомиоцитах, клетках почечных канальцев, париетальных клетках желез дна желудка и т. д. В цитоплазме митохондрии могут распределяться диффузно, но имеют тенденцию аккумулироваться в участках максимального потребления энергии, например в апикальной части реснитчатых клеток, в связующем отделе сперматозоидов, или вблизи ионных насосов (зона базальной исчерченности в проксимальных канальцах почек, в исчерченных протоках слюнных желез и др. ).
 Митохондрии • • • Под электронным микроскопом митохондрии имеют характерную структуру. Каждая митохондрия состоит из наружной и внутренней мембран, между которыми находится межмембранное пространство. Внутренняя мембрана образует складки - кристы, обращенные внутрь митохондрии. Пространство, ограниченное внутренней мембраной, заполнено митохондриальным матриксом, - мелкозернистым материалом различной электронной плотности.
Митохондрии • • • Под электронным микроскопом митохондрии имеют характерную структуру. Каждая митохондрия состоит из наружной и внутренней мембран, между которыми находится межмембранное пространство. Внутренняя мембрана образует складки - кристы, обращенные внутрь митохондрии. Пространство, ограниченное внутренней мембраной, заполнено митохондриальным матриксом, - мелкозернистым материалом различной электронной плотности.
, что") Митохондрии • Наружная мембрана митохондрий содержит много молекул специализированных транспортных белков (например, порин), что обеспечивает её высокую проницаемость, а также белки-рецепторы, распознающие белки, которые переносятся через обе мембраны митохондрий в особых точках их контакта – зонах слипания.
Митохондрии • Наружная мембрана митохондрий содержит много молекул специализированных транспортных белков (например, порин), что обеспечивает её высокую проницаемость, а также белки-рецепторы, распознающие белки, которые переносятся через обе мембраны митохондрий в особых точках их контакта – зонах слипания.
 • • • Внутренняя мембрана митохондрий образует складки – кристы, благодаря чему значительно увеличивается внутренняя поверхность митохондрий. В состав внутренней мембраны входят транспортные белки; ферменты дыхательной цепи и сукцинатдегидрогеназа; комплекс АТФ-синтетазы. На кристах имеются элементарные частицы (оксисомы, или F 1 -частицы), состоящие из округлой головки (9 нм) и цилиндрической ножки. Именно на них происходит сопряжение процессов окисления и фосфорилирования (АДФ → АТФ).
• • • Внутренняя мембрана митохондрий образует складки – кристы, благодаря чему значительно увеличивается внутренняя поверхность митохондрий. В состав внутренней мембраны входят транспортные белки; ферменты дыхательной цепи и сукцинатдегидрогеназа; комплекс АТФ-синтетазы. На кристах имеются элементарные частицы (оксисомы, или F 1 -частицы), состоящие из округлой головки (9 нм) и цилиндрической ножки. Именно на них происходит сопряжение процессов окисления и фосфорилирования (АДФ → АТФ).
") Митохондрии • Чаще всего кристы располагаются перпендикулярно длинной оси митохондрий и имеют пластинчатую (ламеллярную) форму.
Митохондрии • Чаще всего кристы располагаются перпендикулярно длинной оси митохондрий и имеют пластинчатую (ламеллярную) форму.
 Митохондрии • В клетках, синтезирующих стероидные гормоны, кристы имеют вид трубочек или пузырьков - тубулярновезикулярные кристы. В этих клетках ферменты стероидного синтеза частично локализуются на внутренней мембране митохондрий
Митохондрии • В клетках, синтезирующих стероидные гормоны, кристы имеют вид трубочек или пузырьков - тубулярновезикулярные кристы. В этих клетках ферменты стероидного синтеза частично локализуются на внутренней мембране митохондрий
 Митохондрии • Число и площадь крист отражает функциональную активность клеток: наибольшая площадь крист характерна, например, для митохондрий клеток сердечной мышцы, где потребность в энергии постоянно очень велика.
Митохондрии • Число и площадь крист отражает функциональную активность клеток: наибольшая площадь крист характерна, например, для митохондрий клеток сердечной мышцы, где потребность в энергии постоянно очень велика.
 Митохондриальный матрикс • Митохондриальный матрикс – мелкозернистое вещество, заполняющее полость митохондрии. • Матрикс содержит несколько сотен ферментов: ферменты цикла Кребса, окисления жирных кислот, белкового синтеза. • Здесь иногда встречаются митохондриальные гранулы, а также локализуются митохондриальные ДНК, и. РНК, т. РНК, р. РНК и митохондриальные рибосомы.
Митохондриальный матрикс • Митохондриальный матрикс – мелкозернистое вещество, заполняющее полость митохондрии. • Матрикс содержит несколько сотен ферментов: ферменты цикла Кребса, окисления жирных кислот, белкового синтеза. • Здесь иногда встречаются митохондриальные гранулы, а также локализуются митохондриальные ДНК, и. РНК, т. РНК, р. РНК и митохондриальные рибосомы.
 Митохондриальные гранулы • Митохондриальные гранулы – частицы высокой электронной плотности диаметром 20 -50 нм, содержащие ионы Са и Мg.
Митохондриальные гранулы • Митохондриальные гранулы – частицы высокой электронной плотности диаметром 20 -50 нм, содержащие ионы Са и Мg.
 Митохондриальные цитопатологии • Повреждения митохондриальной ДНК в результате мутаций могут привести к развитию ряда патологий митохондриальных цитопатий (синдромы Барта, Патерсона, МERRF (красных разорванных волокон) и др.
Митохондриальные цитопатологии • Повреждения митохондриальной ДНК в результате мутаций могут привести к развитию ряда патологий митохондриальных цитопатий (синдромы Барта, Патерсона, МERRF (красных разорванных волокон) и др.
 макромолекул внеклеточного и") ЛИЗОСОМЫ • Лизосомы – мембранные органеллы, которые обеспечивают внутриклеточное переваривание (расщепление) макромолекул внеклеточного и внутриклеточного происхождения, и обновление компонентов клетки.
ЛИЗОСОМЫ • Лизосомы – мембранные органеллы, которые обеспечивают внутриклеточное переваривание (расщепление) макромолекул внеклеточного и внутриклеточного происхождения, и обновление компонентов клетки.
 Лизосомы • Морфологически лизосомы представляют собой округлые пузырьки, ограниченные мембраной и содержащие большое количество различных гидролаз (более 60 ферментов).
Лизосомы • Морфологически лизосомы представляют собой округлые пузырьки, ограниченные мембраной и содержащие большое количество различных гидролаз (более 60 ферментов).
, протеазы, нуклеазы, сульфатазы,") • Наиболее характерными ферментами лизосом являются: кислая фосфатаза (маркёр лизосом), протеазы, нуклеазы, сульфатазы, липазы, гликозидазы. • Все литические ферменты лизосом представляют собой кислые гидролазы, т. е. оптимум их активности проявляется при р. Н≈5
• Наиболее характерными ферментами лизосом являются: кислая фосфатаза (маркёр лизосом), протеазы, нуклеазы, сульфатазы, липазы, гликозидазы. • Все литические ферменты лизосом представляют собой кислые гидролазы, т. е. оптимум их активности проявляется при р. Н≈5
 обладает протонным насосом, вызывающим закисление среды") Лизосомы • Мембрана лизосом (около 6 нм толщиной) обладает протонным насосом, вызывающим закисление среды внутри органелл, обеспечивает диффузию низкомолекулярных продуктов переваривания макромолекул в гиалоплазму и препятствует утечке литических ферментов в гиалоплазму. • Повреждение мембраны приводит к разрушению клетки вследствие самопереваривания.
Лизосомы • Мембрана лизосом (около 6 нм толщиной) обладает протонным насосом, вызывающим закисление среды внутри органелл, обеспечивает диффузию низкомолекулярных продуктов переваривания макромолекул в гиалоплазму и препятствует утечке литических ферментов в гиалоплазму. • Повреждение мембраны приводит к разрушению клетки вследствие самопереваривания.
 • Лизосомы присутствуют во всех клетках. Особенно много лизосом в тех клетках, где активно протекают процессы фагоцитоза с последующим перевариванием захваченного материала (например, в нейтрофильных гранулоцитах, макрофагах, остеокластах).
• Лизосомы присутствуют во всех клетках. Особенно много лизосом в тех клетках, где активно протекают процессы фагоцитоза с последующим перевариванием захваченного материала (например, в нейтрофильных гранулоцитах, макрофагах, остеокластах).
 Фаголизосомы • • Фаголизосома формируется путем слияния первичной лизосомы с фагосомой мембранным пузырьком, содержащим материал, захваченный клеткой извне. Процесс разрушения этого материала называется гетерофагией. Гетерофагия играет важную роль в функции всех клеток. Особое значение гетерофагия имеет для клеток, осуществляющих защитную функцию, таких как макрофаги и нейтрофильные лейкоциты, которые захватывают и переваривают болезнетворные микроорганизмы.
Фаголизосомы • • Фаголизосома формируется путем слияния первичной лизосомы с фагосомой мембранным пузырьком, содержащим материал, захваченный клеткой извне. Процесс разрушения этого материала называется гетерофагией. Гетерофагия играет важную роль в функции всех клеток. Особое значение гетерофагия имеет для клеток, осуществляющих защитную функцию, таких как макрофаги и нейтрофильные лейкоциты, которые захватывают и переваривают болезнетворные микроорганизмы.
 • Аутофаголизосома образуется при слиянии первичной лизосомы с аутофагосомой мембранным пузырьком, содержащим собственные компоненты клетки, которые подлежат разрушению. • Процесс переваривания внутриклеточного материала называется аутофагией. • Аутофагия обеспечивает постоянное обновление клеточных структур благодаря перевариванию митохондрий, полисом, фрагментов мембран.
• Аутофаголизосома образуется при слиянии первичной лизосомы с аутофагосомой мембранным пузырьком, содержащим собственные компоненты клетки, которые подлежат разрушению. • Процесс переваривания внутриклеточного материала называется аутофагией. • Аутофагия обеспечивает постоянное обновление клеточных структур благодаря перевариванию митохондрий, полисом, фрагментов мембран.
 Остаточные тельца • Остаточные тельца – лизосомы, содержащие непереваренный материал, которые могут находиться в цитоплазме длительное время. • В некоторых долгоживущих клетках (нейроны, кардиомиоциты, гепатоциты) в остаточных тельцах накапливается коричневый эндогенный пигмент липофусцин – «пигмент старения» .
Остаточные тельца • Остаточные тельца – лизосомы, содержащие непереваренный материал, которые могут находиться в цитоплазме длительное время. • В некоторых долгоживущих клетках (нейроны, кардиомиоциты, гепатоциты) в остаточных тельцах накапливается коричневый эндогенный пигмент липофусцин – «пигмент старения» .
 Лизосомные болезни • • • Дефицит лизосомальных ферментов может приводить к развитию ряда заболеваний (болезни накопления), вызванных накоплением в клетках непереваренных веществ, которые нарушают функцию клеток. Примерами могут служить: болезнь Хюрлера, при которой из-за отсутствия α-L-идуронидазы фибробласты и остеобласты накапливают дерматан сульфат, а у больных отмечаются множественные дефекты хондро- и остеогенеза и умственное отставание, болезнь Тэя-Сакса (из-за недостаточности гексозаминидазы А происходит накопление гликолипидов в нервных клетках и поражается нервная система), болезнь Гоше (вследствие наследственного дефекта глюкоцереброзидазы гликолипиды накапливаются в макрофагах и поражаются печень и селезенка) и другие.
Лизосомные болезни • • • Дефицит лизосомальных ферментов может приводить к развитию ряда заболеваний (болезни накопления), вызванных накоплением в клетках непереваренных веществ, которые нарушают функцию клеток. Примерами могут служить: болезнь Хюрлера, при которой из-за отсутствия α-L-идуронидазы фибробласты и остеобласты накапливают дерматан сульфат, а у больных отмечаются множественные дефекты хондро- и остеогенеза и умственное отставание, болезнь Тэя-Сакса (из-за недостаточности гексозаминидазы А происходит накопление гликолипидов в нервных клетках и поражается нервная система), болезнь Гоше (вследствие наследственного дефекта глюкоцереброзидазы гликолипиды накапливаются в макрофагах и поражаются печень и селезенка) и другие.
 ПЕРОКСИСОМЫ • • Пероксисомы – сферические мембранные органеллы диаметром 0. 05 – 1. 5 мкм, с умеренно плотным гомогенным или мелкозернистым матриксом. Мелкие пероксисомы встречаются во всех клетках, а крупные пероксисомы – в гепатоцитах, макрофагах, в клетках канальцев почки. Матрикс пероксисом содержит до 50 различных ферментов, важнейшие из которых: каталаза (маркёр пероксисом), пероксидаза, оксидазы аминокислот, уратоксидаза.
ПЕРОКСИСОМЫ • • Пероксисомы – сферические мембранные органеллы диаметром 0. 05 – 1. 5 мкм, с умеренно плотным гомогенным или мелкозернистым матриксом. Мелкие пероксисомы встречаются во всех клетках, а крупные пероксисомы – в гепатоцитах, макрофагах, в клетках канальцев почки. Матрикс пероксисом содержит до 50 различных ферментов, важнейшие из которых: каталаза (маркёр пероксисом), пероксидаза, оксидазы аминокислот, уратоксидаза.
 Пероксисомы • У некоторых видов животных в пероксисомах выявляется более плотная кристаллическая сердцевина – нуклеоид, состоящая из уратоксидазы. • В пероксисомах клеток человека нуклеотида нет, поскольку отсутствует способность метаболизировать ураты.
Пероксисомы • У некоторых видов животных в пероксисомах выявляется более плотная кристаллическая сердцевина – нуклеоид, состоящая из уратоксидазы. • В пероксисомах клеток человека нуклеотида нет, поскольку отсутствует способность метаболизировать ураты.
 Функции пероксисом: • окисление аминокислот и других субстратов; • защита клетки от действия перекиси водорода, сильного окислителя, образующегося в результате окисления органических соединений, и оказывающего повреждающий эффект на клетку. При этом каталаза пероксисом разлагает перекись водорода на воду и кислород. • участие в расщеплении жирных кислот; • участие в обезвреживании ряда веществ (спирт и др. ).
Функции пероксисом: • окисление аминокислот и других субстратов; • защита клетки от действия перекиси водорода, сильного окислителя, образующегося в результате окисления органических соединений, и оказывающего повреждающий эффект на клетку. При этом каталаза пероксисом разлагает перекись водорода на воду и кислород. • участие в расщеплении жирных кислот; • участие в обезвреживании ряда веществ (спирт и др. ).
 Пероксисомные болезни • Нарушения активности пероксисом вызывает ряд наследственных заболеваний – пероксисомных болезней с тяжелыми нарушениями нервной системы (синдром Целльвегера и др. )
Пероксисомные болезни • Нарушения активности пероксисом вызывает ряд наследственных заболеваний – пероксисомных болезней с тяжелыми нарушениями нервной системы (синдром Целльвегера и др. )
 ЦИТОСКЕЛЕТ • Цитоскелет – сложная трехмерная сеть немембранных органелл: • микротрубочек • микрофиламентов • промежуточных филаментов.
ЦИТОСКЕЛЕТ • Цитоскелет – сложная трехмерная сеть немембранных органелл: • микротрубочек • микрофиламентов • промежуточных филаментов.
 Цитоскелет • Основная функция цитоскелета – опорно-двигательная: • поддержание и изменение формы клеток; • перемещение компонентов внутри клетки; • транспорт веществ внутрь клетки и из клетки; • обеспечение подвижности клетки
Цитоскелет • Основная функция цитоскелета – опорно-двигательная: • поддержание и изменение формы клеток; • перемещение компонентов внутри клетки; • транспорт веществ внутрь клетки и из клетки; • обеспечение подвижности клетки
 Микротрубочки • Микротрубочки – наиболее крупные компоненты цитоскелета. Микротрубочки – полые цилиндрические образования различной длины, с диаметром 2425 нм, с толщиной стенки 5 нм.
Микротрубочки • Микротрубочки – наиболее крупные компоненты цитоскелета. Микротрубочки – полые цилиндрические образования различной длины, с диаметром 2425 нм, с толщиной стенки 5 нм.
 Микротрубочки • Стенка микротрубочки состоит из спирально расположенных нитей – протофиламентов, образованных димерами из глобулярных белковых молекул – α- и βтубулина. • Стенка микротрубочки образована 13 субъединицамипротофиламентами.
Микротрубочки • Стенка микротрубочки состоит из спирально расположенных нитей – протофиламентов, образованных димерами из глобулярных белковых молекул – α- и βтубулина. • Стенка микротрубочки образована 13 субъединицамипротофиламентами.
 Микротрубочки • Микротрубочки могут располагаться в цитоплазме в виде отдельных элементов, в виде пучков, где они связаны тонкими поперечными мостиками, или могут частично сливаться друг с другом, образуя дуплеты (в аксонеме ресничек и жгутиков) и триплеты (в базальном тельце и центриолях
Микротрубочки • Микротрубочки могут располагаться в цитоплазме в виде отдельных элементов, в виде пучков, где они связаны тонкими поперечными мостиками, или могут частично сливаться друг с другом, образуя дуплеты (в аксонеме ресничек и жгутиков) и триплеты (в базальном тельце и центриолях
 Микротрубочки • Микротрубочки представляют собой лабильную систему, в которой сохраняется равновесие между их постоянной сборкой и диссоциацией.
Микротрубочки • Микротрубочки представляют собой лабильную систему, в которой сохраняется равновесие между их постоянной сборкой и диссоциацией.
 Функции микротрубочек – поддержание стабильной формы клеток, и порядка распределения её компонентов; – обеспечение внутриклеточного транспорта, в том числе органелл, пузырьков, секреторных гранул (благодаря некоторым белкам, ассоциированным с микротрубочками); – образование основы центриолей и ахроматинового веретена деления и обеспечение движения хромосом в процессе митоза; – образование основы ресничек и жгутиков, а также обеспечение их движения.
Функции микротрубочек – поддержание стабильной формы клеток, и порядка распределения её компонентов; – обеспечение внутриклеточного транспорта, в том числе органелл, пузырьков, секреторных гранул (благодаря некоторым белкам, ассоциированным с микротрубочками); – образование основы центриолей и ахроматинового веретена деления и обеспечение движения хромосом в процессе митоза; – образование основы ресничек и жгутиков, а также обеспечение их движения.
") Микротрубочки • Угнетение самосборки микротрубочек при действии на клетку блокаторов (колхицин и др. ) вызывает гибель быстроделящихся клеток вследствие отсутствия митотического веретена деления, нарушения транспортных процессов в клетке (аксонный транспорт в нейронах, секреция), изменения форм клетки, дезорганизацию органелл в клетке (в частности, цистерн ЭПС)
Микротрубочки • Угнетение самосборки микротрубочек при действии на клетку блокаторов (колхицин и др. ) вызывает гибель быстроделящихся клеток вследствие отсутствия митотического веретена деления, нарушения транспортных процессов в клетке (аксонный транспорт в нейронах, секреция), изменения форм клетки, дезорганизацию органелл в клетке (в частности, цистерн ЭПС)
 Клеточный центр • Клеточный центр образован двумя полыми цилиндрическими структурами центриолями, которые расположены под прямым углом друг к другу.
Клеточный центр • Клеточный центр образован двумя полыми цилиндрическими структурами центриолями, которые расположены под прямым углом друг к другу.
 Центриоли • • Каждая центриоль представляет собой короткий цилиндр длиной ~ 0, 5 мкм и диаметром ~ 0, 2 мкм, состоящий из 9 триплетов частично слившихся трубочек (А, В и С), связанных поперечными белковыми мостиками. Формула строения центриоли описывается как (9 × 3) + 0, так как в центральной части микротрубочки отсутствуют. Каждый триплет центриоли связан с глобулярными белковыми тельцами – сателлитами, от которых отходят микротрубочки, образующие центросферу.
Центриоли • • Каждая центриоль представляет собой короткий цилиндр длиной ~ 0, 5 мкм и диаметром ~ 0, 2 мкм, состоящий из 9 триплетов частично слившихся трубочек (А, В и С), связанных поперечными белковыми мостиками. Формула строения центриоли описывается как (9 × 3) + 0, так как в центральной части микротрубочки отсутствуют. Каждый триплет центриоли связан с глобулярными белковыми тельцами – сателлитами, от которых отходят микротрубочки, образующие центросферу.
 Центриоли • • В неделящейся клетке выявляется одна пара центриолей – диплосома, которая располагается обычно вблизи ядра. Перед делением клетки в Sпериоде интерфазы происходит дупликация центриолей: под прямым углом к каждой зрелой (материнской) центриоли пары образуется новая (дочерняя) центриоль. В ранней профазе митоза пары центриолей расходятся к полюсам клетки и служат центрами образования микротрубочек ахроматинового веретена деления.
Центриоли • • В неделящейся клетке выявляется одна пара центриолей – диплосома, которая располагается обычно вблизи ядра. Перед делением клетки в Sпериоде интерфазы происходит дупликация центриолей: под прямым углом к каждой зрелой (материнской) центриоли пары образуется новая (дочерняя) центриоль. В ранней профазе митоза пары центриолей расходятся к полюсам клетки и служат центрами образования микротрубочек ахроматинового веретена деления.
 Реснички и жгутики • Реснички и жгутики являются выростами цитоплазмы, обладающие подвижностью. Основу ресничек и жгутиков составляет каркас из микротрубочек, называемый аксонемой. • Длина ресничек равна 2 -10 мкм, а их количество на поверхности одной клетки может составлять до нескольких сотен. • В организме человека жгутик есть только в одном типе клеток – сперматозоидах. При этом один сперматозоид имеет один жгутик длиной 50 -70 мкм.
Реснички и жгутики • Реснички и жгутики являются выростами цитоплазмы, обладающие подвижностью. Основу ресничек и жгутиков составляет каркас из микротрубочек, называемый аксонемой. • Длина ресничек равна 2 -10 мкм, а их количество на поверхности одной клетки может составлять до нескольких сотен. • В организме человека жгутик есть только в одном типе клеток – сперматозоидах. При этом один сперматозоид имеет один жгутик длиной 50 -70 мкм.
 и") Аксонема • • Аксонема образована 9 периферическими парами микротрубочек (микротрубочки А и В) и одной центрально расположенной парой; такое строение описывается формулой (9 × 2) + 2. Центральная пара микротрубочек окружена центральной оболочкой, от которой к периферическим дуплетам расходятся радиальные спицы. Периферические дублеты связаны друг с другом мостиками белка нексина, а от микротрубочки А к микротрубочке В соседнего дублета отходят “ручки” из белка динеина, который обладает АТФ-азной активностью, что необходимо для скольжения соседних дублетов в аксонеме, вызывающих движение (биение) ресничек и жгутиков
Аксонема • • Аксонема образована 9 периферическими парами микротрубочек (микротрубочки А и В) и одной центрально расположенной парой; такое строение описывается формулой (9 × 2) + 2. Центральная пара микротрубочек окружена центральной оболочкой, от которой к периферическим дуплетам расходятся радиальные спицы. Периферические дублеты связаны друг с другом мостиками белка нексина, а от микротрубочки А к микротрубочке В соседнего дублета отходят “ручки” из белка динеина, который обладает АТФ-азной активностью, что необходимо для скольжения соседних дублетов в аксонеме, вызывающих движение (биение) ресничек и жгутиков
 Микрофиламенты • Микрофиламенты – тонкие белковые нити диаметром 57 нм, расположенные в цитоплазме поодиночке, в виде сетей или упорядоченными пучками (в скелетной и сердечной мышцах). • Основной белок микрофиламентов – актин – встречается в клетках как в мономерной форме (глобулярный G-актин), так и в виде полимерного фибриллярного F-актина
Микрофиламенты • Микрофиламенты – тонкие белковые нити диаметром 57 нм, расположенные в цитоплазме поодиночке, в виде сетей или упорядоченными пучками (в скелетной и сердечной мышцах). • Основной белок микрофиламентов – актин – встречается в клетках как в мономерной форме (глобулярный G-актин), так и в виде полимерного фибриллярного F-актина
 Функции микрофиламентов • • • в мышечных волокнах и клетках актиновые микрофиламенты образуют упорядоченные пучки и при взаимодействии с миозиновыми филаментами обеспечивают их сокращение. в немышечных клетках микрофиламенты образуют кортикальную (терминальную) сеть, в которой микрофиламенты сшиты с помощью особых белков (филамин и др. ). Кортикальная сеть, с одной стороны, обеспечивает поддержание формы клетки, а с другой - способствует изменениям формы плазмолеммы, обеспечивая, таким образом, функции эндо- и экзоцитоза, миграции клеток, образования псевдоподий. микрофиламенты тесно связаны с органеллами, транспортными пузырьками, секреторными гранулами и играют важную роль в их перемещении внутри цитоплазмы. микрофиламенты формируют сократимую перетяжку (срединное тельце) при цитотомии, завершающей клеточное деление. микрофиламенты участвуют в организации структуры межклеточных соединений (zonula adherens – поясок сцепления). микрофиламенты являются основой специальных выростов цитоплазмы – микроворсинок и стереоцилий.
Функции микрофиламентов • • • в мышечных волокнах и клетках актиновые микрофиламенты образуют упорядоченные пучки и при взаимодействии с миозиновыми филаментами обеспечивают их сокращение. в немышечных клетках микрофиламенты образуют кортикальную (терминальную) сеть, в которой микрофиламенты сшиты с помощью особых белков (филамин и др. ). Кортикальная сеть, с одной стороны, обеспечивает поддержание формы клетки, а с другой - способствует изменениям формы плазмолеммы, обеспечивая, таким образом, функции эндо- и экзоцитоза, миграции клеток, образования псевдоподий. микрофиламенты тесно связаны с органеллами, транспортными пузырьками, секреторными гранулами и играют важную роль в их перемещении внутри цитоплазмы. микрофиламенты формируют сократимую перетяжку (срединное тельце) при цитотомии, завершающей клеточное деление. микрофиламенты участвуют в организации структуры межклеточных соединений (zonula adherens – поясок сцепления). микрофиламенты являются основой специальных выростов цитоплазмы – микроворсинок и стереоцилий.
 Микроворсинки • Микроворсинки – пальцевидные выросты цитоплазмы клетки диаметром 0. 1 мкм и длиной 1 мкм, основу которых образуют актиновые микрофиламенты. • Микроворсинки обеспечивают многократное увеличение площади поверхности клетки. На апикальной поверхности некоторых клеток, активно участвующих в процессах расщепления и всасывания веществ, имеется до несколько тысяч микроворсинок, образующих в совокупности щёточную каемку (эпителий тонкой кишки и почечных канальцев).
Микроворсинки • Микроворсинки – пальцевидные выросты цитоплазмы клетки диаметром 0. 1 мкм и длиной 1 мкм, основу которых образуют актиновые микрофиламенты. • Микроворсинки обеспечивают многократное увеличение площади поверхности клетки. На апикальной поверхности некоторых клеток, активно участвующих в процессах расщепления и всасывания веществ, имеется до несколько тысяч микроворсинок, образующих в совокупности щёточную каемку (эпителий тонкой кишки и почечных канальцев).
 Микроворсинка • Основа каждой микроворсинки – пучок, содержащий около 40 микрофиламентов, расположенных вдоль её длинной оси. Микрофиламенты имеют поперечные сшивки из белков (фимбрин, виллин), и прикреплены к плазмолемме особыми белковыми мостиками (минимиозин). • У основания микроворсинки микрофиламенты пучка вплетаются в терминальную сеть
Микроворсинка • Основа каждой микроворсинки – пучок, содержащий около 40 микрофиламентов, расположенных вдоль её длинной оси. Микрофиламенты имеют поперечные сшивки из белков (фимбрин, виллин), и прикреплены к плазмолемме особыми белковыми мостиками (минимиозин). • У основания микроворсинки микрофиламенты пучка вплетаются в терминальную сеть
 Стереоцилии • Стереоцилии – длинные, иногда ветвящиеся микроворсинки, имеющие каркас из микрофиламентов. Они встречаются редко (в главных клетках эпителия протока придатка семенника).
Стереоцилии • Стереоцилии – длинные, иногда ветвящиеся микроворсинки, имеющие каркас из микрофиламентов. Они встречаются редко (в главных клетках эпителия протока придатка семенника).
 Промежуточные филаменты • Промежуточные филаменты в клетках различных типов различаются по своей химической природе и молекулярному весу. • Выделяются 6 основных классов промежуточных филаментов
Промежуточные филаменты • Промежуточные филаменты в клетках различных типов различаются по своей химической природе и молекулярному весу. • Выделяются 6 основных классов промежуточных филаментов
 • Цитокератины – промежуточные филаменты, характерные для клеток эпителия • Этот класс включает около 20 близких полипептидов (тонофиламентов). Кератиновые филаменты входят в состав десмосом и полудесмосом, участвуют в образовании рогового вещества в эпителии кожи и являются главным компонентом волос и ногтей.
• Цитокератины – промежуточные филаменты, характерные для клеток эпителия • Этот класс включает около 20 близких полипептидов (тонофиламентов). Кератиновые филаменты входят в состав десмосом и полудесмосом, участвуют в образовании рогового вещества в эпителии кожи и являются главным компонентом волос и ногтей.
. Десмины играют") Десмины • Десмины – промежуточные филаменты мышечных тканей (за исключением миоцитов сосудов). Десмины играют важную роль в организации миофибрилл в мышечной ткани и обеспечении сократительной функции.
Десмины • Десмины – промежуточные филаменты мышечных тканей (за исключением миоцитов сосудов). Десмины играют важную роль в организации миофибрилл в мышечной ткани и обеспечении сократительной функции.
 Промежуточные филаменты • Нейрофиламенты – промежуточные филаменты нейронов, которые играют важную роль в поддержании формы отростков нервных клеток. • Глиальные клетки содержат глиальный фибриллярный кислый белок и встречаются только в клетках глии (астроциты, олигодендроциты).
Промежуточные филаменты • Нейрофиламенты – промежуточные филаменты нейронов, которые играют важную роль в поддержании формы отростков нервных клеток. • Глиальные клетки содержат глиальный фибриллярный кислый белок и встречаются только в клетках глии (астроциты, олигодендроциты).
 Промежуточные филаменты в диагностике • • Идентификация классов промежуточных филаментов (методами иммуноцитохимии с антителами к данному типу промежуточных филаментов) имеет большое значение в диагностике опухолей, и, следовательно, в прогнозе и выборе противоопухолевого лечения. Так, выявление различных форм кератинов свидетельствует о недифференцированных опухолях эпителиального происхождения, карциномах, аденокарциномах. Десмин является маркёром опухолей мышечного происхождения, а глиальный фибриллярный кислый белок – маркёр опухолей глиального происхождения.
Промежуточные филаменты в диагностике • • Идентификация классов промежуточных филаментов (методами иммуноцитохимии с антителами к данному типу промежуточных филаментов) имеет большое значение в диагностике опухолей, и, следовательно, в прогнозе и выборе противоопухолевого лечения. Так, выявление различных форм кератинов свидетельствует о недифференцированных опухолях эпителиального происхождения, карциномах, аденокарциномах. Десмин является маркёром опухолей мышечного происхождения, а глиальный фибриллярный кислый белок – маркёр опухолей глиального происхождения.
 КЛЕТОЧНЫЙ ЦИКЛ • Клеточный цикл – совокупность процессов, происходящих в клетке между двумя последовательными делениями или между её образованием и гибелью. • Клеточный цикл включает в себя собственно митотическое деление и интерфазу – промежуток между делениями.
КЛЕТОЧНЫЙ ЦИКЛ • Клеточный цикл – совокупность процессов, происходящих в клетке между двумя последовательными делениями или между её образованием и гибелью. • Клеточный цикл включает в себя собственно митотическое деление и интерфазу – промежуток между делениями.
 ИНТЕРФАЗА • Интерфаза занимает около 90% всего времени клеточного цикла и подразделяется на три периода: • пресинтетический или постмитотический – G 1 (от англ. gap – промежуток); • синтетический – S; • постсинтетический или премитотический - G 2.
ИНТЕРФАЗА • Интерфаза занимает около 90% всего времени клеточного цикла и подразделяется на три периода: • пресинтетический или постмитотический – G 1 (от англ. gap – промежуток); • синтетический – S; • постсинтетический или премитотический - G 2.
 Пресинтетический период – G 1 • Пресинтетический период – G 1 – характеризуется активным ростом клетки, синтезом белка и РНК, благодаря чему клетка восстанавливает необходимый набор органелл и достигает нормальных размеров. G 1 период длится от нескольких часов до нескольких дней. • В течение этого периода синтезируются особые «запускающие» белки – активаторы S периода. Они обеспечивают достижение клеткой точки R (точки ограничения), после которого она вступает в S-период.
Пресинтетический период – G 1 • Пресинтетический период – G 1 – характеризуется активным ростом клетки, синтезом белка и РНК, благодаря чему клетка восстанавливает необходимый набор органелл и достигает нормальных размеров. G 1 период длится от нескольких часов до нескольких дней. • В течение этого периода синтезируются особые «запускающие» белки – активаторы S периода. Они обеспечивают достижение клеткой точки R (точки ограничения), после которого она вступает в S-период.
 Период репродуктивного покоя • • • Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G 0). Клетки некоторых тканей под влиянием определенных факторов способны возвращаться из периода G 0 в клеточный цикл, клетки других тканей утрачивают эту способность по мере дифференцировки. Абсолютное большинство дифференцированных клеток организма, выполняющих свои специфические функции, не делятся.
Период репродуктивного покоя • • • Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G 0). Клетки некоторых тканей под влиянием определенных факторов способны возвращаться из периода G 0 в клеточный цикл, клетки других тканей утрачивают эту способность по мере дифференцировки. Абсолютное большинство дифференцированных клеток организма, выполняющих свои специфические функции, не делятся.
 ДНК, синтезом гистонов и других") Синтетический период • Синтетический период –Sхарактеризуется репликацией (удвоением содержания) ДНК, синтезом гистонов и других белков. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. • S-период длится у большинства клеток 8 -12 часов.
Синтетический период • Синтетический период –Sхарактеризуется репликацией (удвоением содержания) ДНК, синтезом гистонов и других белков. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. • S-период длится у большинства клеток 8 -12 часов.
 Постсинтетический период • Постсинтетический период – G 2 длится 2 -4 часа и продолжается вплоть до митоза. • В течение этого периода запасается энергия, и синтезируются белки, в частности тубулины, необходимые для процесса деления.
Постсинтетический период • Постсинтетический период – G 2 длится 2 -4 часа и продолжается вплоть до митоза. • В течение этого периода запасается энергия, и синтезируются белки, в частности тубулины, необходимые для процесса деления.
 Регуляция клеточного цикла • По уровню обновления ткани организма подразделяются на три группы – три типа клеточных популяций: • (1) Обновляющиеся клеточные популяции характеризуются постоянным обновлением. Естественная убыль дифференцированных клеток, специализированных к выполнению определенных функций и неспособных к делению уравновешена образованием новых клеток в результате деления малодифференцированных камбиальных клеток и последующей дифференцировки (физиологическая регенерация). • К таким популяциям относят клетки костного мозга и крови, эпителий кишки, эпидермис кожи.
Регуляция клеточного цикла • По уровню обновления ткани организма подразделяются на три группы – три типа клеточных популяций: • (1) Обновляющиеся клеточные популяции характеризуются постоянным обновлением. Естественная убыль дифференцированных клеток, специализированных к выполнению определенных функций и неспособных к делению уравновешена образованием новых клеток в результате деления малодифференцированных камбиальных клеток и последующей дифференцировки (физиологическая регенерация). • К таким популяциям относят клетки костного мозга и крови, эпителий кишки, эпидермис кожи.
 Растущие клеточные популяции способны к увеличению массы ткани за") Растущие клеточные популяции • (2) Растущие клеточные популяции способны к увеличению массы ткани за счет нарастания числа клеток и их полиплоидизации. • Их долгоживущие клетки выполняют специализированные функции, но сохраняют способность при стимуляции, под действием некоторых факторов вновь вступать в клеточный цикл, чтобы восстановить свою нормальную численность. • К растущим популяциям относят эпителий почек, различных желез, печени.
Растущие клеточные популяции • (2) Растущие клеточные популяции способны к увеличению массы ткани за счет нарастания числа клеток и их полиплоидизации. • Их долгоживущие клетки выполняют специализированные функции, но сохраняют способность при стимуляции, под действием некоторых факторов вновь вступать в клеточный цикл, чтобы восстановить свою нормальную численность. • К растущим популяциям относят эпителий почек, различных желез, печени.
 Стабильные клеточные популяции состоят из высокоспециализированных клеток с полной") Стабильные клеточные популяции • (3) Стабильные клеточные популяции состоят из высокоспециализированных клеток с полной потерей способности к делению. К таким популяциям относятся нейроны, кардиомиоциты.
Стабильные клеточные популяции • (3) Стабильные клеточные популяции состоят из высокоспециализированных клеток с полной потерей способности к делению. К таким популяциям относятся нейроны, кардиомиоциты.
 Регуляция клеточного цикла • Регуляция клеточного цикла в различных тканях организма осуществляется сложной системой механизмов, стимулирующих или ингибирующих клеточное деление. • Протоонкогены (около 50) – группа генов-активаторов, контролирующих клеточное деление и дифференцировку. Изменения структуры и усиление активности экспрессии протоонкогенов вызывает развитие опухолей. Повышение активности протоонкогенов может быть связано с изменениями строениями ДНК (в результате мутаций), увеличением количества генов (генной амплификации) или их перегруппировкой, при которой гены размещаются вблизи активного промотора (т. e. , участка ДНК, ответственного за инициацию транскрипции). • Злокачественная трансформация клетки может возникнуть не только вследствие повышения активности протоонкогенов, но и в результате снижения активности другой группы генов, называемых антионкогенами.
Регуляция клеточного цикла • Регуляция клеточного цикла в различных тканях организма осуществляется сложной системой механизмов, стимулирующих или ингибирующих клеточное деление. • Протоонкогены (около 50) – группа генов-активаторов, контролирующих клеточное деление и дифференцировку. Изменения структуры и усиление активности экспрессии протоонкогенов вызывает развитие опухолей. Повышение активности протоонкогенов может быть связано с изменениями строениями ДНК (в результате мутаций), увеличением количества генов (генной амплификации) или их перегруппировкой, при которой гены размещаются вблизи активного промотора (т. e. , участка ДНК, ответственного за инициацию транскрипции). • Злокачественная трансформация клетки может возникнуть не только вследствие повышения активности протоонкогенов, но и в результате снижения активности другой группы генов, называемых антионкогенами.
 является универсальным механизмом деления соматических клеток. • Во время митоза") Митоз • Митоз (кариокинез) является универсальным механизмом деления соматических клеток. • Во время митоза родительская клетка делится, и каждая из дочерних клеток получает набор хромосом идентичный родительскому, и, таким образом, происходит равномерное распределение генетического материала. Продолжительность митоза – 1 -3 часа.
Митоз • Митоз (кариокинез) является универсальным механизмом деления соматических клеток. • Во время митоза родительская клетка делится, и каждая из дочерних клеток получает набор хромосом идентичный родительскому, и, таким образом, происходит равномерное распределение генетического материала. Продолжительность митоза – 1 -3 часа.
 Митоз • Митоз включает 4 основные фазы: • профазу • метафазу, • анафазу и • телофазу.
Митоз • Митоз включает 4 основные фазы: • профазу • метафазу, • анафазу и • телофазу.
 Профаза • • Профаза начинается с конденсации хромосом, которые становятся видимыми в световой микроскоп как нитевидные структуры. Каждая хромосома состоит из двух параллельно лежащих сестринских хроматид, связанных в области центромеры. Ядерная оболочка распадается на мембранные пузырьки и исчезает к концу профазы, так же как и ядрышко. Кариоплазма смешивается с цитоплазмой. Пары центриолей расходятся к противоположным полюсам клетки и дают начало микротрубочкам митотического веретена.
Профаза • • Профаза начинается с конденсации хромосом, которые становятся видимыми в световой микроскоп как нитевидные структуры. Каждая хромосома состоит из двух параллельно лежащих сестринских хроматид, связанных в области центромеры. Ядерная оболочка распадается на мембранные пузырьки и исчезает к концу профазы, так же как и ядрышко. Кариоплазма смешивается с цитоплазмой. Пары центриолей расходятся к противоположным полюсам клетки и дают начало микротрубочкам митотического веретена.
 метафаза • В метафазе хромосомы выстраиваются в области экватора митотического веретена (в равной удаленности от центриолей противоположных полюсов), и образуют картину экваториальной (метафазной) пластинки (вид сбоку) или материнской звезды (вид со сторону полюсов). • Сестринские хроматиды к концу этой фазы разделяются щелью, однако удерживаются в области центромеры.
метафаза • В метафазе хромосомы выстраиваются в области экватора митотического веретена (в равной удаленности от центриолей противоположных полюсов), и образуют картину экваториальной (метафазной) пластинки (вид сбоку) или материнской звезды (вид со сторону полюсов). • Сестринские хроматиды к концу этой фазы разделяются щелью, однако удерживаются в области центромеры.
 Анафаза • • • Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды (в области центромеры) и движения дочерних хромосом к противоположным полюсам клеток, происходящего вдоль микротрубочек. Анафаза завершается скоплением на полюсах клетки двух идентичных наборов хромосом, которые образуют картину звезд (стадия дочерних звезд). В конце анафазы начинает образовываться клеточная перетяжка, благодаря сокращению актиновых микрофиламентов, концентрирующихся по окружности клетки.
Анафаза • • • Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды (в области центромеры) и движения дочерних хромосом к противоположным полюсам клеток, происходящего вдоль микротрубочек. Анафаза завершается скоплением на полюсах клетки двух идентичных наборов хромосом, которые образуют картину звезд (стадия дочерних звезд). В конце анафазы начинает образовываться клеточная перетяжка, благодаря сокращению актиновых микрофиламентов, концентрирующихся по окружности клетки.
 Телофаза • • Телофаза характеризуется реконструкцией ядер дочерних клеток и завершением их разделения. Ядерная оболочка восстанавливается, хромосомы постепенно деспирализуются, замещаясь картиной хроматина интерфазного ядра, а в конце телофазы вновь появляется ядрышко. Углубление клеточной перетяжки завершается полной цитотомией с формированием двух дочерних клеток. При этом происходит распределение органелл между дочерними клетками
Телофаза • • Телофаза характеризуется реконструкцией ядер дочерних клеток и завершением их разделения. Ядерная оболочка восстанавливается, хромосомы постепенно деспирализуются, замещаясь картиной хроматина интерфазного ядра, а в конце телофазы вновь появляется ядрышко. Углубление клеточной перетяжки завершается полной цитотомией с формированием двух дочерних клеток. При этом происходит распределение органелл между дочерними клетками
 Эндомитоз и полиплоидизация • • • Эндомитоз – процесс увеличения числа хромосом внутри ядерной оболочки без последующего деления клетки, что приводит к повышенному содержанию ДНК в ядре – полиплоидии. Полиплоидные ядра имеют больший объем. Полиплоидные клетки могут также возникнуть вследствие митотического деления без последующей цитотомией. При таком делении образуются двуядерные клетки с увеличенным вдвое набором хромосом. Основной смысл развития полиплоидии заключается в усилении функциональной активности клеток. Наличие полиплоидных – тетра- (4 n, если 1 n – гаплоидный набор хромосом) и октаплоидных (8 n) клеток – нормальное явление для гепатоцитов (клеток печени), переходного эпителия мочевого пузыря, секреторных клеток поджелудочной и слюнных желез. Уровень полиплоидизации мегакариоцитов красного костного мозга достигает – 16 -32 n.
Эндомитоз и полиплоидизация • • • Эндомитоз – процесс увеличения числа хромосом внутри ядерной оболочки без последующего деления клетки, что приводит к повышенному содержанию ДНК в ядре – полиплоидии. Полиплоидные ядра имеют больший объем. Полиплоидные клетки могут также возникнуть вследствие митотического деления без последующей цитотомией. При таком делении образуются двуядерные клетки с увеличенным вдвое набором хромосом. Основной смысл развития полиплоидии заключается в усилении функциональной активности клеток. Наличие полиплоидных – тетра- (4 n, если 1 n – гаплоидный набор хромосом) и октаплоидных (8 n) клеток – нормальное явление для гепатоцитов (клеток печени), переходного эпителия мочевого пузыря, секреторных клеток поджелудочной и слюнных желез. Уровень полиплоидизации мегакариоцитов красного костного мозга достигает – 16 -32 n.
 Антионкогены • Антионкогены – гены, которые продуцируют супрессоры опухолевого роста, угнетающие митотическую активность клеток. • Пример антионкогенов – ген р53. Ген р53 обеспечивает поддержание стабильности генетического аппарата и контролирует клеточный цикл. Его экспрессия резко усиливается при повреждении ДНК. Активация гена р53 приводит к остановке клеточного цикла (выходу в G 0 ) для репарации ДНК, а при тяжелых повреждениях запускает программу апоптоза (клеточной гибели). • Выявлена четкая связь между утратой функции гена р53 (в результате мутации или делеции) и развитием более 50 видов злокачественных опухолей у человека.
Антионкогены • Антионкогены – гены, которые продуцируют супрессоры опухолевого роста, угнетающие митотическую активность клеток. • Пример антионкогенов – ген р53. Ген р53 обеспечивает поддержание стабильности генетического аппарата и контролирует клеточный цикл. Его экспрессия резко усиливается при повреждении ДНК. Активация гена р53 приводит к остановке клеточного цикла (выходу в G 0 ) для репарации ДНК, а при тяжелых повреждениях запускает программу апоптоза (клеточной гибели). • Выявлена четкая связь между утратой функции гена р53 (в результате мутации или делеции) и развитием более 50 видов злокачественных опухолей у человека.
 Факторы роста • Факторы роста представляют собой гликопептиды, продуцируемые клетками различных тканей, усиливающие митотическую активность в определенных клетках-мишенях, имеющих специфические рецепторы на плазмолемме. • К ним относятся фактор роста нервов, инсулиноподобные факторы роста, колониестимулирующие факторы, интерлейкины и другие цитокины.
Факторы роста • Факторы роста представляют собой гликопептиды, продуцируемые клетками различных тканей, усиливающие митотическую активность в определенных клетках-мишенях, имеющих специфические рецепторы на плазмолемме. • К ним относятся фактор роста нервов, инсулиноподобные факторы роста, колониестимулирующие факторы, интерлейкины и другие цитокины.
 Кейлоны • Кейлоны, напротив, подавляют клеточное деление. Кейлоны образуются всеми зрелыми дифференцированными клетками и локально воздействуют на камбиальные элементы (стволовые и полустволовые клетки) этой же ткани. • Они обеспечивают стабильную численность клеточной популяции, а их выделение контролируется механизмом отрицательной обратной связи. • При уменьшении численности зрелых клеток данной популяции (например, потеря лейкоцитов при кровотечении или эпидермиса при ранении) продукция кейлонов снижается, что приводит к усилению митотической активности клеток, способных к делению, - репаративной регенерации
Кейлоны • Кейлоны, напротив, подавляют клеточное деление. Кейлоны образуются всеми зрелыми дифференцированными клетками и локально воздействуют на камбиальные элементы (стволовые и полустволовые клетки) этой же ткани. • Они обеспечивают стабильную численность клеточной популяции, а их выделение контролируется механизмом отрицательной обратной связи. • При уменьшении численности зрелых клеток данной популяции (например, потеря лейкоцитов при кровотечении или эпидермиса при ранении) продукция кейлонов снижается, что приводит к усилению митотической активности клеток, способных к делению, - репаративной регенерации
 Клеточная гибель • Число клеток в организме, органах и тканях регулируется гомеостатическими механизмами и динамическим равновесием между образованием клеток и их гибелью. • Гибель клеток, наряду с их размножением и дифференцировкой, является одним из ключевых процессов в обеспечении нормальной жизнедеятельности различных тканей
Клеточная гибель • Число клеток в организме, органах и тканях регулируется гомеостатическими механизмами и динамическим равновесием между образованием клеток и их гибелью. • Гибель клеток, наряду с их размножением и дифференцировкой, является одним из ключевых процессов в обеспечении нормальной жизнедеятельности различных тканей
 Гибель клеток • При гибели клеток могут наблюдаться два вида морфологических изменений, которые соответствуют различным механизмам её развития: • некроз и • апоптоз
Гибель клеток • При гибели клеток могут наблюдаться два вида морфологических изменений, которые соответствуют различным механизмам её развития: • некроз и • апоптоз
 • переохлаждения") Некроз • Возникает под действием резко выраженных повреждающих факторов: • перегревания (гипертермии) • переохлаждения (гипотермии) • недостатка кислорода (гипоксии) • нарушения кровоснабжения (ишемии) • метаболических ядов • механической травмы и др. • Некроз – «смерть в результате несчастного случая»
Некроз • Возникает под действием резко выраженных повреждающих факторов: • перегревания (гипертермии) • переохлаждения (гипотермии) • недостатка кислорода (гипоксии) • нарушения кровоснабжения (ишемии) • метаболических ядов • механической травмы и др. • Некроз – «смерть в результате несчастного случая»
 Некроз Поздние явления при некрозе включают: • разрыв ядерной оболочки, плазмолеммы и мембран органелл, • разрушение и растворение ядра (кариолизис) • исчезновение клеточных границ и распад клетки
Некроз Поздние явления при некрозе включают: • разрыв ядерной оболочки, плазмолеммы и мембран органелл, • разрушение и растворение ядра (кариолизис) • исчезновение клеточных границ и распад клетки
 гибель клеток – «смерть клетки в результате самоубийства (самоуничтожения)» -") Апоптоз • Физиологическая (запрограммированная) гибель клеток – «смерть клетки в результате самоубийства (самоуничтожения)» - активный, генетически контролируемый процесс, регулируемый внутренней программой, которая запускается внешними факторами (воздействие разнообразных повреждающих физических и химических факторов, умеренные по интенсивности; некоторые инфекции (вирусные); воздействие физиологических активаторов (индукторов) апоптоза; дефицит стимулирующих факторов, потеря контакта с другими клетками и др.
Апоптоз • Физиологическая (запрограммированная) гибель клеток – «смерть клетки в результате самоубийства (самоуничтожения)» - активный, генетически контролируемый процесс, регулируемый внутренней программой, которая запускается внешними факторами (воздействие разнообразных повреждающих физических и химических факторов, умеренные по интенсивности; некоторые инфекции (вирусные); воздействие физиологических активаторов (индукторов) апоптоза; дефицит стимулирующих факторов, потеря контакта с другими клетками и др.
") Апоптоз • Развитие апоптоза индуцируется особыми генами (киллерными генами – р53 и др. ) • Это энергоёмкий процесс и сопровождается активацией сигнальных систем в клетке.
Апоптоз • Развитие апоптоза индуцируется особыми генами (киллерными генами – р53 и др. ) • Это энергоёмкий процесс и сопровождается активацией сигнальных систем в клетке.
 Апоптоз • • Развитие апоптоза морфологически на светооптическом уровне проявляется уплотнением ядра (кариопикноз и кариорексис без разрушения кариолеммы), конденсацией цитоплазмы, которая уплотняется, сморщивается и уменьшается в размерах, органеллы при этом сохраняют свою целостность. При прогрессировании апоптоза изменяется форма клетки – образуются многочисленные крупные вздутия и выросты на поверхности – и происходит распад клетки на фрагменты – апоптозные тела.
Апоптоз • • Развитие апоптоза морфологически на светооптическом уровне проявляется уплотнением ядра (кариопикноз и кариорексис без разрушения кариолеммы), конденсацией цитоплазмы, которая уплотняется, сморщивается и уменьшается в размерах, органеллы при этом сохраняют свою целостность. При прогрессировании апоптоза изменяется форма клетки – образуются многочисленные крупные вздутия и выросты на поверхности – и происходит распад клетки на фрагменты – апоптозные тела.
 Т киллер индуцирует апоптоз раковой клетки
Т киллер индуцирует апоптоз раковой клетки