Дыхательная цепь.ppt
- Количество слайдов: 47



ЖИРЫ БЕЛКИ УГЛЕВОДЫ Аминокислоты ГЛЮКОЗА ПВК ГЛИЦЕРИН ПВК ВЖК Ацетил-Ко. А КЕТОКИСЛОТЫ Ацетил-Ко. А Цикл Кребса

Биологическим окислением или тканевым дыханием называется процесс потребления кислорода Окислительное фосфорилирование – синтез АТФ, который использует энергию электрохимического потенциала переноса электронов мембраны митохондрий. Для этого пути синтеза АТФ необходим кислород. Общая реакция синтеза АТФ: АДФ+НАДН Н+О 2 +Н 3 РО 4 АТФ + НАД+ +Н 2 О

Дыхательная цепь это комплекс специфических белков, встроенных в мембрану митохондрий, способных транспортировать электроны от восстановленных эквивалентов на кислород. Электроны, поступающие в эту цепь переносчиков, богаты энергией, по мере продвижения их по цепи, от одного переносчика к другому, они теряют свободную энергию. Значительная часть этой энергии запасается в виде АТФ.
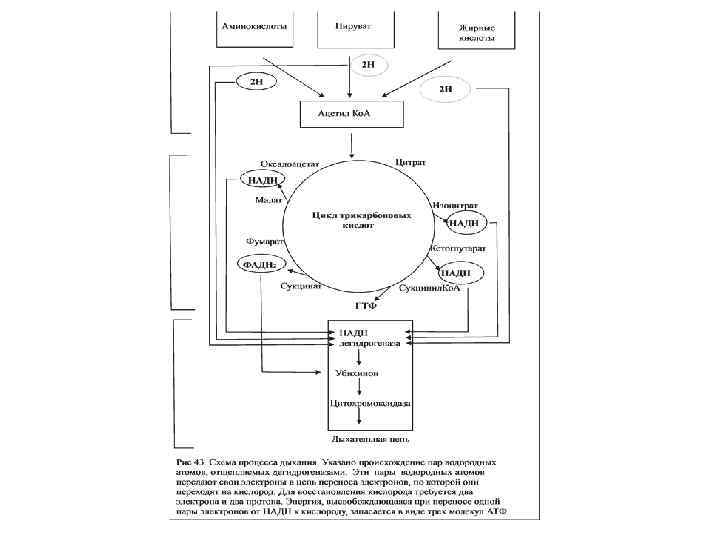

Дыхательная цепь состоит из: 1. Пиридинзависимых дегидрогеназ, которые в качестве коферментов содержат НАД либо НАДФ 2. Флавинзависимых дегидрогеназ, у которых роль простетической группы выполнят ФАД или ФМН. 3. Убихинона коэнзим Q 4. Цитохромов – белков содержащих гем.

Общая характеристика белков дыхательной цепи переносчики Простетическая Донор Акцептор группа электронов NADH дегидрогеназа, FMN, Fe. S комплекс I Сукцинатдегидрогеназа FAD, Fe. S Комплекс II Коэнзим Q, убихинон NADH QH 2 дегидрогеназа, Комплекс III Цитохром с NADH, цит bс1 Гем с QH 2 Цитохромоксидаза Комплекс IV Гем а Cu 2+ Ко. Q сукцинат Ко. Q NADH Комплекс III, Цит b c 1 Цитохром с комплекс Комплекс III IV Цито O 2 хром с

Окисленные и восстановленные формы НАД и ФАД

Убихинон Q является производным бензохинона с длинной боковой цепью, которая в большинстве тканей млекопитающих состоит из 10 изопреновых единиц. Восстановленная форма флавиновых ферментов в цепи дыхательных катализаторов передает атомы водорода Ко. Q. Как всякий хинон Ко. Q может существовать в окисленной и восстановленной формах Окисленная и восстановленная форма убихинона

Цитохромы Дальнейший перенос электронов от Ко. Q на кислород осуществляет система цитохромов. Цитохромы это гемопротеины 5 типов b, c, c 1, a, a 3, Внутри комплекса III электроны передаются от цитохромов b на железосерные центры, потом на цитохром с1, а затем на цитохром с. Цитохромы а и а 3 (цитохромоксидаза) образуют IV комплекс переносчиков электронов. Особенность химического строения цитохромоксидазы заключается в наличии ионов меди, меняющих свою валентность. Цитохромоксидаза непосредственно реагирует с молекулярным кислородом.
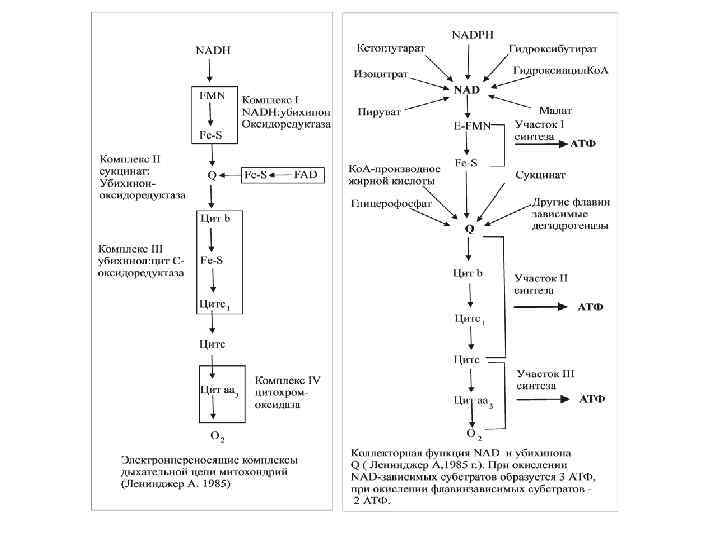

Последовательность белков определяется их окислительно-восстановительными потенциалами

• В каждом из 3 пунктов: • 1 -й пункт - это НАДН-дегидрогеназный комплекс; • 2 -й пункт - QН 2 -цитохром -с-редуктазный комплекс; • 3 -й пункт - цитохром-с-оксидазный комплекс • при переносе пары электронов от НАДН 2 одновременно на наружную поверхность внутренней мембраны Мх, т. е. в межмембранное пространство выносится по паре протонов.
 и фосфорилирования (синтеза АТФ) Каким же образом энергия электронов")
Теории сопряжения окисления (переноса электронов) и фосфорилирования (синтеза АТФ) Каким же образом энергия электронов запасается в виде АТФ? Самыми первыми гипотезами сопряжения окисления и фосфорилирования были химическая и механохимическая гипотезы, предполагавшие наличие некого промежуточного, высокоэнергетического вещества преобразующего энергию электронов в энергию АТФ. В настоящее время эти гипотезы представляют исторический интерес. Современной теорией сопряжения является хемиосмотическая теория английского биохимика Питера Митчелла (1961 г. ). Суть этой теории заключается в том, что при прохождении электронов по дыхательной цепи высвобождается энергия, которая используется для перемещения протонов из матрикса в межмембранное пространство. Особое значение в транспорте водорода в межмембранное пространство отводится Ко. Q, который является липофильным веществом и может легко перемещаться по мембране
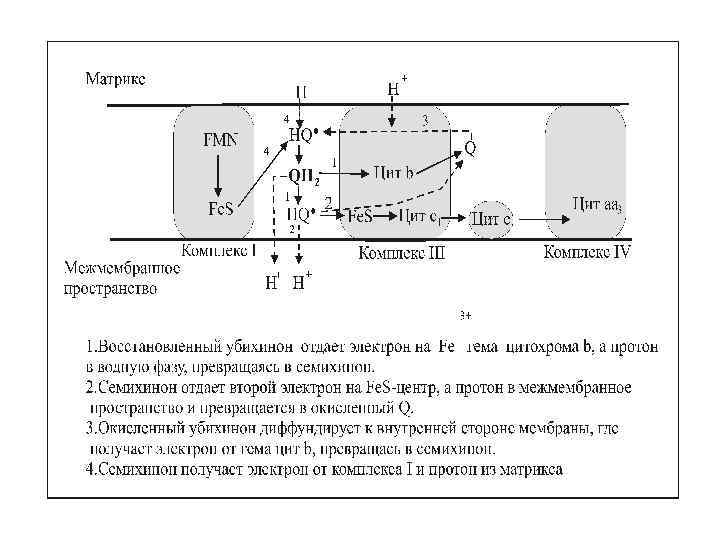

В результате между двумя сторонами внутренней мембраны создается электрохимический протонный градиент: н = + z р Н, - мембранный потенциал (разница зарядов наружной и внутренней, отрицательной поверхности мембраны) z р Н – градиент р. Н ( формируется за счет более кислой среды межмембранного пространства)

Электрохимический потенциал заставляет протоны двигаться в обратном направлении – с наружной поверхности внутрь. Однако внутренняя мембрана непроницаема для них, за исключением специальных пор – протонных каналов. В области этих каналов на внутренней поверхности внутренней мембраны располагается АТФ синтетаза. Энергия обратного тока электронов по этому градиенту используется АТФ синтетазой, катализирующей образование АТФ по следующей реакции: АДФ + Н 3 РО 4 АТФ + Н 2 О

АТФ синтетаза – интегральный белок внутренней мембраны митохондрий. Он расположен в непосредственной близости к дыхательной цепи. АТФ синтетаза состоит из 2 белковых комплексов, обозначаемых как F 0 и F 1. Гидрофобный комплекс F 0 погружен в мембрану. Он служит основанием, которое фиксирует АТФ синтетазу в мембране. Комплекс F 0 состоит из нескольких субъединиц, образующих канал, по которому протоны переносятся в матрикс.

Комплекс F 1 выступает в митохондриальный матрикс. Он состоит из 9 субъединиц, которые образуют сферический вырост. Повышение концентрации протонов в межмембранном пространстве активирует АТФ синтетазу. Электрохимический потенциал заставляет протоны двигаться по каналу АТФ синтетазы в матрикс. Параллельно под действием электрохимического потенциала происходит конформационное изменение субъединиц F 1 комплекса, в результате чего из АДФ и неорганического фосфата образуется АТФ. Образующаяся АТФ, при участии фермента адениннуклеотидтранслоказы с абсолютной специфичностью, транспортируется из матрикс на наружную сторону мембраны и попадает в цитозоль. АДФ транспортируется за счет антипорта АТФ. Фосфат транспортируется по типу симпорта с ионами водорода при участии фермента фосфаттранслоказы.
 Ингибиторы")
ИНГИБИТОРЫ ДЫХАТЕЛЬНОЙ ЦЕПИ И ОКИСЛИТЕЛЬНОГО ФОСФОРИЛИРОВАНИЯ Ингибиторы можно разделить на 3 группы: 1) Ингибиторы собственно дыхательной цепи; 2)Ингибиторы окислительного фосфорилирования; 3)Разобщители окислительного фосфорилирования.
 Ингибиторы, блокирующие дыхательную цепь, могут действовать в 3 местах, соответствующих трем комплексам дыхательной")
1) Ингибиторы, блокирующие дыхательную цепь, могут действовать в 3 местах, соответствующих трем комплексам дыхательной цепи: В первом комплексе дыхательная цепь ингибируется барбитуратами (например, амобарбиталом), а также ротеноном (высокотоксичное в-во, добываемое из растений, которое использовалось американскими индейцами в качестве яда при ловле рыб). Эти ингибиторы препятствуют НАД-зависимому окислению субстратов.

• Второй участок, где может блокироваться дыхательная цепь это участок между цитохромом b и цитохромом с, т. е. во втором комплексе. Таким ингибитором является антибиотик антимицин А. • Третий участок это цитохромоксидаза. Он ингибируется классическими ядами - сернистым водородом, окисью углерода и цианидом.
 К ингибиторам, подавляющим стадию фосфорилирования относится антибиотик олигомицин. Он полностью блокирует")
• 2) К ингибиторам, подавляющим стадию фосфорилирования относится антибиотик олигомицин. Он полностью блокирует окисление и фосфорилирование в интактных митохондриях за счет того, что не пропускает протоны через канал F 0. • Атрактилозид также ингибирует окислительное фосфорилирование. Он ингибирует фермент - транслоказу, который производит поступление АДФ в митохондрию и выход АТФ из митохондрий.

Разобщители отличаются тем, что они нарушают систему сопряжения процессов окисления в дыхательной цепи и фосфорилирования. В этих условиях процесс дыхания происходит неконтролируемым образом. Т. е. концентрации АДФ и Рi в этом случае не являются лимитирующими. • Различают разные типы разобщения

Механизм разобщения: Снижение электрохимического градиента н. Устранение мембранного потенциала и/или протонного градиента. Динитрофенол - градиент протонов Валиномицин – перенос ионов калия Грамицидин – перенос ионов калия, натрия

Разобщители, как правило легко растворимы в липидном слое мембран, этим и объясняется их действие как разобщителей. • Среди разобщителей надо назвать некоторые лекарственные вещества: строфантин, папаверин, салицилаты. • Разобщителями могут быть и собственные соединения организма, его естественные метаболиты, например, жирные кислоты. Это имеет большое значение для организма на

Снижение температуры возбуждает терморецепторы, это ведет к повышению выделения норадреналина. Затем увеличивается липолиз, повышается содержание ВЖК, увеличивается концентрация ацетил-Ко. А. Это ведет к повышению активности цикла Кребса, повышению содержания НАДН и ФАДН 2. Увеличивается активность тканевого дыхания, возрастает электрохимический потенциал.

• ВЖК снижают электрохимический потенциал, синтез АТФ снижается, но н будет расходоваться на согревание. При этом на холоде человек дрожит. Рефлекторно возникает возбуждение терморецепторов, снижается содержание АТФ, увеличивается АДФ. Активируется ЦТК. Увеличивается концентрация НАДН, повышается электрохимический потенциал - человек согревается.
 железами в очень малых концентрациях (нмоль/л,")
Гормоны это химические соединения, синтезируемые специализированными (эндокринными) железами в очень малых концентрациях (нмоль/л, пмоль/л), оказывающие действие вдали от места синтеза (дистантность), обладающие высокой биологической активностью (эффект усиления).

Гипоталамус либерины и статины Тиролиберин Тиреотропный кортиколиберин гонадолиберин Гипофиз тропные гормоны Гонадотропный АКТГ гормон пролактин Периферические железы Щитовидная железа надпочечники Половые железы Молочные железы

Классификация и механизм действия гормонов Существует несколько видов классификации гормонов: 1. По железам внутренней секреции разделяют: гормоны гипоталамуса, гипофиза, щитовидной, паращитовидной, поджелудочной желез и т. д. 2. По биологическому действию можно выделить гормоны, влияющие на углеводный, белковый, липидный обмен, минеральный, водно солевой обмены.
 Белки и пептиды– к ним относятся,")
3. По химической природе гормоны делят на: а) Белки и пептиды– к ним относятся, например, гормоны гипоталамуса, гипофиза, инсулин, глюкагон б) Гликопротеины – тиреотропный гормон, ФСГ, ЛГ в) Производные аминокислот – адреналин, тироксин, трийодтиронин г) Стероидные: гормоны коры надпочечников, половые гормоны

По механизму действия гормоны подразделяют на: 1. мембранный механизм действия – для гормонов, влияющих на структуру и проницаемость мембраны клеток, например белковые гормоны, адреналин, инсулин. 2. внутриклеточный механизм действия для гидрофобных гормонов, которые могут приникать внутрь клетки (стероиды и гормоны щитовидной железы)

Классификация является условной, т. к. практически очень трудно выявить механизм действия гормонов. В последнее время в молекулярной биологии используется классификация гормонов по физико-химическим свойствам (полярности). Все гормоны подразделяются на две неравные группы: I. полярные, или гидрофильные гормоны — белки, пептиды и производные аминокислот (кроме тиреоидных гормонов). II. неполярные, или гидрофобные гормоны — стероиды (половые гормоны и гормоны коры надпочечников. ) и тиреоидные гормоны.

Механизм действия гормонов Подразделение гормонов на гидрофильные и гидрофобные имеет принципиальное значение, т. к. связано с механизмом действия. а) Гидрофильные гормоны не способны проникать через плазмолемму. Должен существовать специальный механизм для восприятия сигнала и передачи его на эффекторные структуры. б) Гидрофобные же гормоны проходят через мембраны клетки и действуют внутриклеточно (в «сопровождении» специального рецепторного белка, достигают определенных областей хромосом).
 гормона на по верхности клеток")
Общая схема действия гидрофильных гормонов Для каждого гидрофильного (полярного) гормона на по верхности клеток мишеней имеются белки-рецепторы. Возбуждение рецептора гормоном ведет к изменению концентрации в клетке определенного внутриклеточного медиатора. Такими медиаторами (вторичным посредником) могут быть ц. АМФ, ц. ГМФ, Инозитолтрифосфат оксид азота (N 0), эйкозаноиды, и т. д.

ц. АМФ Гормоны, взаимодействие которых с рецепторами клетки мишени приводит к образованию ц. АМФ, действуют через трехкомпонентную систему. Эта система (еще ее называют аденилатциклазной) включает белок рецептор, G белок и фермент аденилатциклазу. G белок состоит из 3 субъединиц , , . В отсутствии гормонального сигнала эти субъединицы связаны между собой, а субъединица еще связана с ГМФ. Субъединица бывает двух типов стимулирующего ( s) и ингибирующего ( I). G белок контактирует с рецептором, воспринимающим внешний сигнал.

Связывание гормона с рецептором приводит к изменению структуры G белка: 1. вначале субъединица теряет сродство к ГМФ и заменяет его на ГТФ 2. В связанном с ГТФ состоянии субъединица высвобождается из комплекса, диффундирует по внутренней поверхности и активирует (если это s) или ингибирует ( если I) аденилатциклазу 3. через некоторое время ГТФ гидролизуется до ГМФ, в результате чего субъединица отделяется от АЦ и связывается с другими субъединицами G белка (т. е. возвращается в исходное состояние) 4. Аденилатциклаза синтезирует из АТФ циклический аденозинмонофосфат (ц. АМФ) 5. ц. АМФ активирует протеинкиназу
 — специальные регуляторные ферменты, способные фосфорилировать (за счет фосфатных групп АТФ) строго")
Протеинкиназы (ПК) — специальные регуляторные ферменты, способные фосфорилировать (за счет фосфатных групп АТФ) строго определенные белки, причем по строго определенным аминокислотным остаткам серина, треонина или тирозина (все эти аминокислоты содержат в радикале гидроксильную группу). Фосфорилирование (и обратное ему дефосфорилирование под действием протеинфосфатаз) — один из наиболее универсальных способов регуляции активности белков — как структурных, так и ферментов (см. регуляцию активности ферментов). Такая химическая модификация белка меняет его конфигурацию. В итоге в одних случаях активность белка в результате фосфорилирования повышается, в других — напротив, снижается. ц. АМФ гидролизуется фосфодиэстеразой. Ингибиторы фосфодиэстеразы (кофеин, например), увеличивая внутриклеточный уровень ц. АМФ, усиливают действие гормонов.

Инозитолтрифосфат является вторичным посредником для гормонов, связывающихся с рецептором подобному 2 адренорецептору, который активирует G белок, последний, в свою очередь, стимулирует фосфолипазу С. Основным субстратом этого фермента является мембранный липид – фосфатидилинозитолдифосфат (см химию липидов). При действии гормона в мембране клеток мишеней увеличивается активность фосфолипазы С, которая расщепляет фосфатидилинозитол 4 5 дифосфат на инозитол-1, 4, 5 трифосфат (ИТФ) и диацилглицерол (ДАГ). ИТФ как полярное вещество легко диффундирует внутри клетки, связываясь с кальцевыми каналами плазмолеммы и эндоплазматического ретикулума. В результате чего повышается концентрация кальция. Кальций активирует белки ферменты, кальмодулин. Кальций зависимыми ферментами являются, например, киназа фосфорилазы, пируваткарбоксилаза, дегидрогеназы, киназа гликогенсинтетазы.

Диацилглицерол неполярное гидрофобное вещество, которое способно лишь к латеральной диффузии в составе плазмолеммы. Он активирует еще один мембраносвязанный регуляторный фермент – протеинкиназу С (ПК С). У ПК С множество белков мишеней, что и определяет физиологический ответ.
, а каскад из двух или")
В регуляторной цепочке нередко имеется не одна ПК (протеинкиназа), а каскад из двух или даже более протеинкиназ. Первая из них (которая непосредственно активируется вторичным посредником фосфорилирует другую ПК, а эта вторая ПК действует уже на непосредственные объекты регуляции. В качестве последних могут быть ключевые ферменты метаболизма, структурные белки, факторы транскрипции или трансляции. Фосфорилирование или дефосфорилирование каких то из этих белков и вызывает тот конечный эффект, который «требовал» от клетки действующий на нее гормональный сигнал: • изменяется активность соответствующих ферментов или структурных белков, • изменяется активность соответствующих генов и скорость синтеза ферментов или структурных белков.
 гормонов мембранные рецепторы не требуются: как")
Общая схема действия гидрофобных гормонов Для гидрофобных (неполярных) гормонов мембранные рецепторы не требуются: как уже отмечалось, эти гормоны диффундируют через плазмолемму клетки мишени. Правда, отсюда следует, что они могут проникать и в любые другие клетки. Но в цитоплазме (или в ядрах) клеток мишеней содержатся специфические рецепторные белки, связывающие соответствующие гормоны. Благодаря этому, диффузия гормона в клетку мишень оказывается значительно более интенсивной, чем в прочие клетки.
 проникает в клеточное ядро, достигая тех")
Комплекс рецептор гормон (если он образуется в цитоплазме) проникает в клеточное ядро, достигая тех или иных генов, причем активность одних генов может возрастать, а других (в той же клетке) — уменьшаться.
 к соответствующим участкам ДНК")
Комплекс рецептор гормон влияет на сродство определенных транскрипционных факторах (ТЕ) к соответствующим участкам ДНК (энхансерам). В каких то случаях это сродство повышается и тогда РНК полимераза с большей скоростью связывается с промотором регулируемого гена, начинает считывать информацию, т. е. стимулируется транскрипция, а с ней и синтез определенного белка. В других случаях, напротив, сродство ТР к энхансеру снижается и синтез белка тормозится. Не исключено также, что комплекс рецептор гормон и сам может выступать в роли транскрипционного фактора.
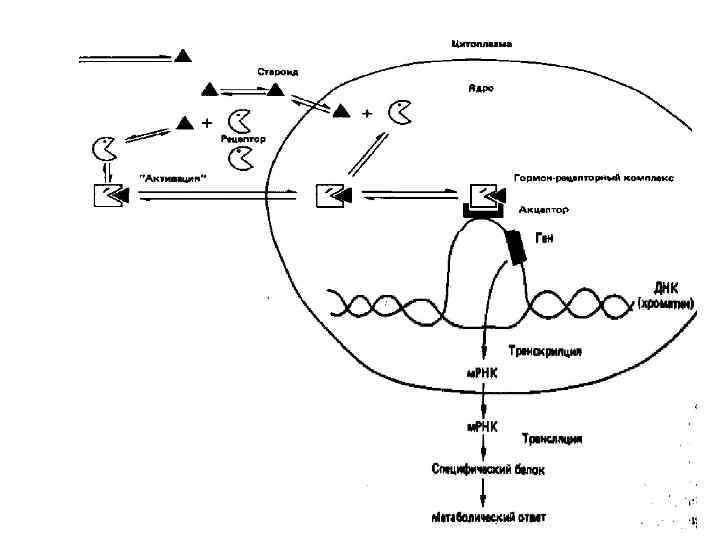

Гормон проникает через мембрану, связывается с рецептором, активирует экспрессию определённого гена
, так и на")
Итак, если гидрофильные гормоны могут влиять как на активность ферментов (белков), так и на их синтез, то гидрофобные гормоны лишь на синтез белков, реализуемое на генном уровне.
Дыхательная цепь.ppt