флт_Збудлструктури2012.ppt
- Количество слайдов: 81



Збудливі тканини : нервова, м'язова, залозиста
 г — це наука про життєві процеси, діяльність окремих органів")
Фізіолоія(грец. φυσιολογία — природознавство) г — це наука про життєві процеси, діяльність окремих органів та їх системи і в цілому всього організму. Основним у фізіології є експериментальний метод дослідження. Виникнення фізіології як науки пов'язане з ім'ям англійського вченого Вільяма Гарвея, який відкрив замкнений кровообіг (1628 р. ) і опублікував працю “Анатомічнідані про рух серця і крові тварин” у.
 —- відносна сталістьскладу та властивостей внутрішнього")
Гомеостаз(давн. -грец — однаковий, подібнийта στάσις—. стан, нерухомість) —- відносна сталістьскладу та властивостей внутрішнього середовища біологічних систем різних рівнів організації. В основі гомеостазу лежать динамічні процеси, оскільки сталість внутрішнього середовища безперервно порушується і так само безперервно відновлюється. Теоретичне обґрунтування поняття “гомеостаз” зробили французький фізіолог Клод Бернар та американськийфізіолог Уолтер Кеннон Організм як тварини, так і людини є відкритоюсистемою, що досить сильно пов'язна з середовищем. Існує таке поняття, як саморегуляція, для якої було запропоновано назву "gomios stasis" лат. подібний стан. Гомеостаз - це постійністьвнутрішнього середовищаі фізіологічних функцій організму(сталістьхімічного складу та фізико-хімічних властивостей, він виражається наявністю ряду стійких показників (констант), характеризує нормальнийстан організма температуру, : тиск крові, кількістьу крові речовин( К, Na). Са,

Подразливість – здатність живого при зміні зовнішнього середовища активнопротидіяти змінам гомеостатичних показниківорганізму поза певні межі через зміни своєї структури та функцій

Збудження – це складний фізіологічний процес швидкої тимчасової зміни трансмембранної різниці потенціалів клітин, який проявляється спеціалізованою реакцією тканини (проведення нервового імпульсу, скорочення м’язу, виділення секрету тощо)

Мембранний потенціал спокою КЛІТИНИ ЗБУДЛИВИХ ТКАНИН, ЯК І КЛІТИНИ ІНШИХ ТКАНИН, ЗЗОВНІ ВКРИТІ КЛІТИННОЮ МЕМБРАНОЮ. ОДНІЄЮ З ХАРАКТЕРНИХ ОСОБЛИВОСТЕЙ ЦІЄЇ МЕМБРАНИ Є ТЕ, ЩО НАВІТЬ ЗА УМОВ ПОВНОГО СПОКОЮ (ВІДСУТНОСТІПОДРАЗНЕННЯ)МЕМБРАНА КЛІТИН ЗБУДЛИВИХ ТКАНИН ЕЛЕКТРИЧНО ПОЛЯРИЗОВАНА. ІНШИМИ СЛОВАМИ, ВОНА МАЄ РІЗНИЦЮ ЕЛЕКТРИЧНИХ ЗАРЯДІВ МІЖ ЗОВНІШНЬОЮ І ВНУТРІШНЬОЮ ПОВЕРХНЯМИ МЕМБРАНИ. ТАКУ РІЗНИЦЮ ЗАРЯДІВНАЗИВАЮТЬ МЕМБРАННИМ ПОТЕНЦІАЛОМ СПОКОЮ

Будова клітинної мембрани n Схема біліпідного шару
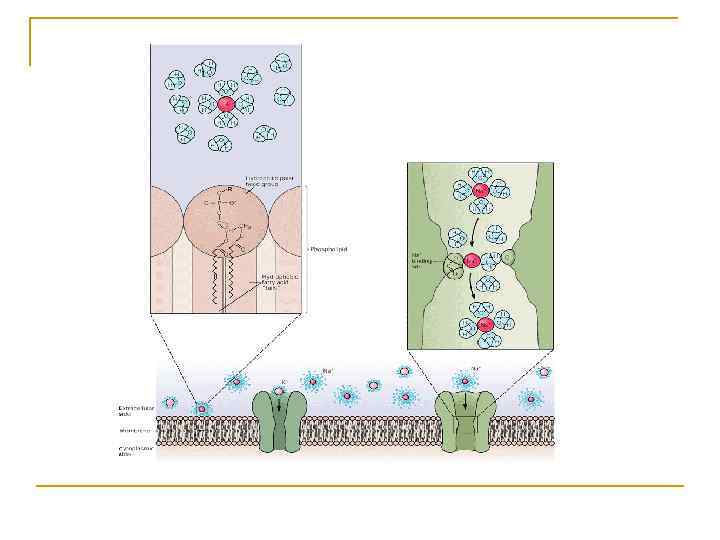

Методика вимірювання мембранного потенціалу спокою § Поза- та внутрішньоклітинна реєстрація

Вміст іонів всередині і ззовні аксона кальмара та нейрона

Розподіл іонів на мембрані нейрона

Типи каналів мембрани n Канали та насоси

Типи каналів мембрани

Загальна схема формування мембранної різниці зарядів на штучній напівпроникній мембрані, яка розділяє розчини з різною концентрацією іонів

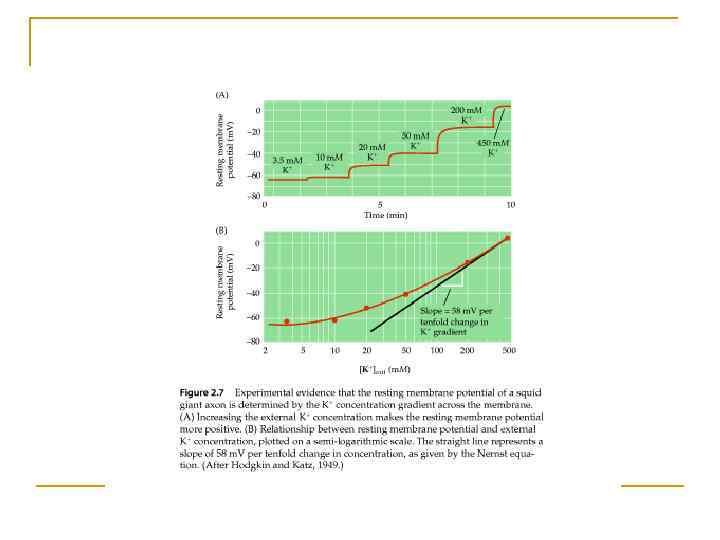
Формування МП іонами К+ n Концентраційний і електричний градієнти
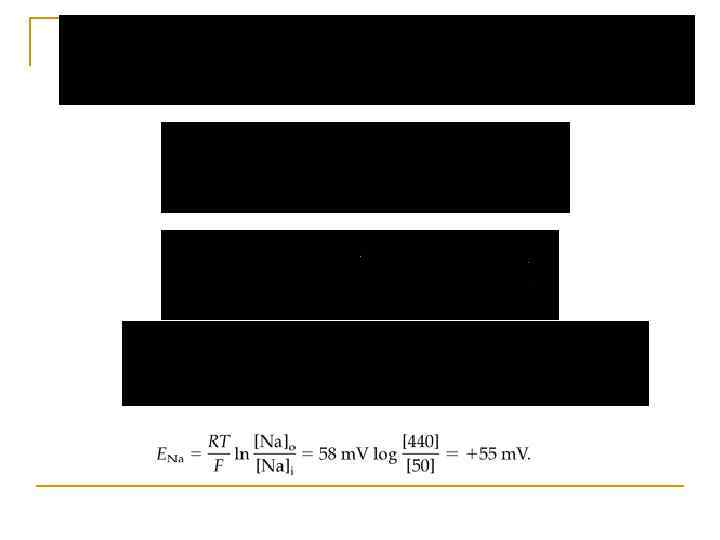

Рівняння. Гольдмана q В стані спокою PК : РNa : РCl становить 1 : 0, 04 : 0, 45.
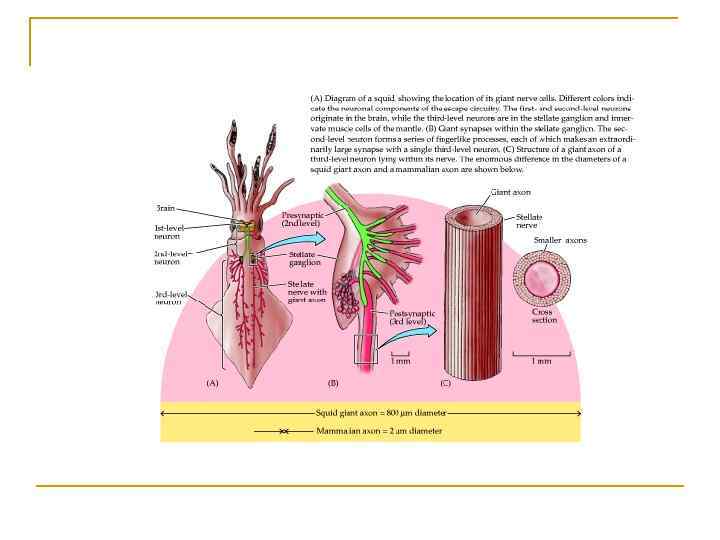
 ,")
Перша внутрішньоклітинна реєстрація ПД в гігантському аксоні кальмара( Ходжкін , Хакслі 1939) ,

Вигляд потенціалу дії при внутрішньоклітинній реєстрації n Фази потенціалу дії

Роль іонів Na+в формуванні ПД

Методика фіксації потенціалу для дослідження іонних струмів формування потенціалу дії

Трансмембранні струми n Струми при дії ТЕА і ТТХ

Загальні зміни натрієвої і калієвої провідності при формуванні ПД n Провідність Na+, K+

Зміни збудливості при формуванні ПД

Відкривання каналу через конформацію молекули каналу при деполяризації мембрани

Ворітний механізм активації і інактивації n Робота воріт

Дослідження роботи окремих каналів n Петч-клемп

Дослідження роботи певних видів каналів в ооцитахафриканської жаби

Молекулярна структура натрієвого каналу електричного органу вугра (1832 амінокислоти, 4 домени по 6 сегментів, пора знаходиться між 5 і 6 сегментом )
; Б – загальний вигляд")
Загальна схема будови калієвого каналу А-окремасубодиниця (домен); Б – загальний вигляд

Механізми роботи окремих каналів n Струми

Селективний фільтр натрієвого каналу
")
Дія постійного струму на мембрану (полярний закон Пфлюгера )

Зміни МП при подразненні струмом різної сили і напрямку

Електрична схема мембрани

Пасивні зміни МП з часом n Постійна часу мембрани
 зміни МП з відстанню n Постійна довжини мембрани")
Пасивні електротонічні ( ) зміни МП з відстанню n Постійна довжини мембрани

Формування мієлінових оболонок

Механізм проведення ПД по немієлінізованому нервовому волокну

Механізм проведення ПД по мієлінізованому нервовому волокну

Швидкість проведення збудження прямопропорційна сталійдовжинімембраниі оберненопропорційнасталійчасу .

Будова електричного синапсу

Тонка будова щільного контакту та доказ відсутності синаптичної затримки
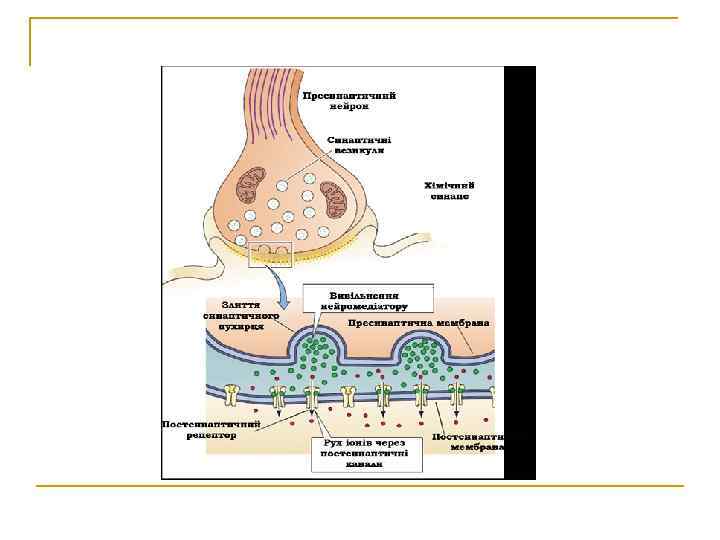

Етапи передач збдженняв хімічному і синапсі

Передача нервового імпульсу у хімічному синапсі

Аксоннийтранспорт

Аксоннийтранспорт

Механізмаксонноготранспорту

Загальна схема будови джгутика а поперечний розріз джгутика: - центральні поодинокі 1 мікротрубочки 2 - центральна , оболонка, 3 - дінеїновіручки, 4 - радіальна спиця, - субфібрила. А, 6 - субфібрила. В, 7 5 нексиновиймісток, 8 – мембрана джгутика; схема, яка показує взаємне ковзання б периферичних дублетів мікротрубочок (дублетк ковзає вздовж дублета завдяки дії п динеїнових ручок)

Будова м'язового волокна і саркомера скелетного м'яза

Загальна будова м'язового волокна

Загальна схема будови посмугованого м'язового волокна

Загальна схема будови молекули міозину

Схема м'язової системи кісткових риб 1 - міомери 2 - міосепти ;


Моторна одиниця
 та повільні (І) АТФази")
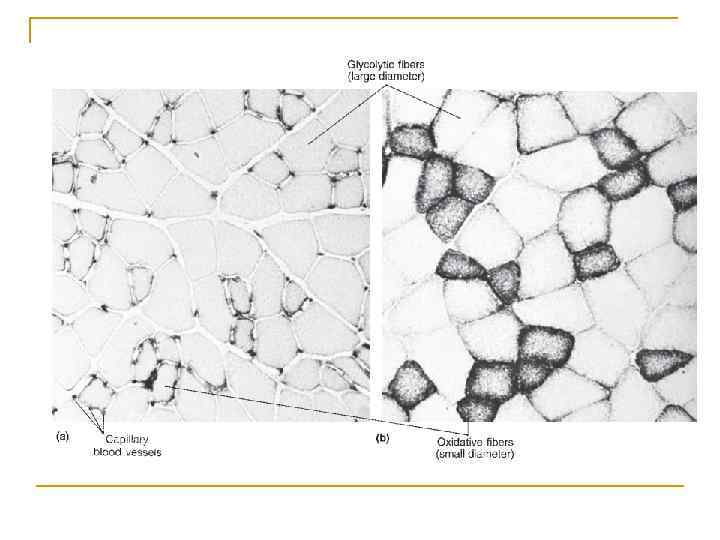
Біопсія скелетного м'язу: швидкі та повільн м'язові волокна (швидкі (ІІ) та повільні (І) АТФази )
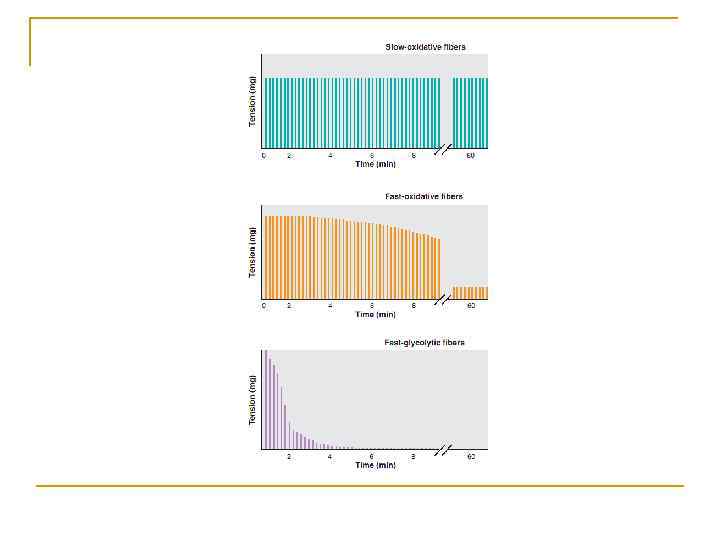

Співвідношення між ПД і скороченням

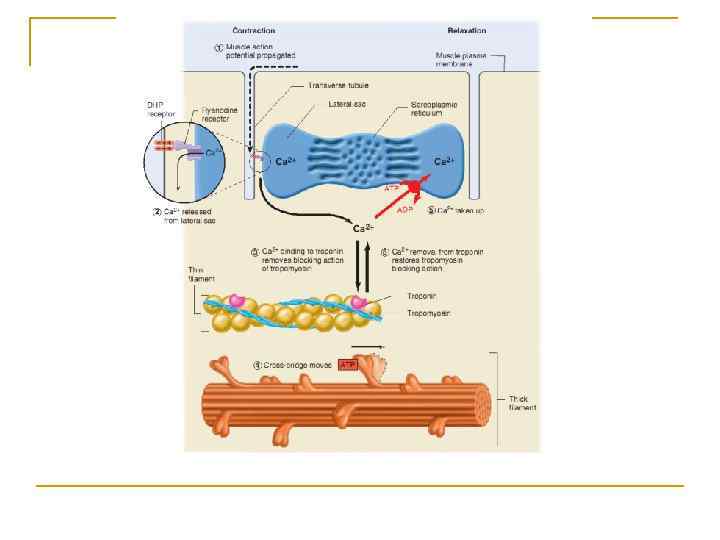
Електромеханічне спряження
")
Одночасна реєстрація потенціалу дії і скорочення фазного м'язового волокна щура (методика подвійного сахарозного містка) а). Схематичнезображення послідовності процесу збудження-скорочення посмугованому в м'язовомуволокні (б - г): а: 1 скорочення, 2 - потенціал дії; б - м'язове волокно в розслабленомустані, поверхнева (1) і тубулярна (2) мембрани поляризовані, потенціалспокоюдорівнює-90 м. В, внутрішньоклітинна концентрація іонів Са 2+ - 10 -7 М, АТФ - 5 10 -3 М, 3, 4 - термінальніцистерни і повздовжні трубочки саркоплазматичного ретикулюму відповідно; С - саркомер в - під час , , потенціалудії різниця потенціалівна поверхневійі тубулярнихмембранахреверсує і робитьсявсерединіпозитивною (+30 м. В), відбувається стимуляція виділенняйонів Са 2+ із цистерн саркоплазматичного ретикулюму г - в кінці потенціалудії , внутрішньоклітинна концентрація іонів Са 2+ досягає 10– 5 М, потенціал спокою майже відновлюєтьсядо початкової величини (-80 м. В).

Будова актоміозинового комплексу

Механіка скорочення

Молекулярний механізм скорочення

Можливий механізм м'язового скорочення

Залежність скорочення м'язу від частоти подразнення

Типовий вигляд шару гладких м'язів

Схема скорочення гладком'язової клітини

Гладкі м'язи

Схема іннервації гладких мязів
 гладких м'язів")
Схема побудови унітарних (вісцеральних) гладких м'язів

Молекулярний механізм скорочення гладких і посмугованих м'язів

Гладком'язові пейсмекери
 та сечоводу (В)")
Електрична та скоротлива активність гладких м'язів шлунка (А) та сечоводу (В)

Вплив різних чинників на зміни МП гладких м'язів

Дякую за увагу!
флт_Збудлструктури2012.ppt