23_09_16_LAURA.ppt
- Количество слайдов: 153



Зат алмасуға кіріспе. Тамақтану биохимиясы. Энергетикалық алмасу. Мақсаты: Тамақтанудың метаболизмі мен биохимиясы және энергети калық алмасумен катаболи зімнің жалпы жолдары тура лы түсінік қалыптастыру.

ЖОСПАРЫ: -Зат алмасуға кіріспе. -Тамақтану биохимиясы. -Энергетикалық алмасу.

Жануарлар ағзасының жасушалары үшін энергетикалық субстраттар - көмірсулар, липид тер , белоктар қажет 1: 1: 4 .

Состав пищи человека

Тәуелдік калориялықтың үлесі. Белок - 14% липидтер - 30% , көмірсулар - 56%

Толық құнарлы рацион 5 -компонент Энегияның қайнар көзі Көмірсулар Липидтер Ақуыздар

Аустырырмайтың амин қышқылдар арг, гис, изо, лей, лиз, мет, фен, тре, три, вал

Ауыстырылмай тың май қышқылдар. Линоль қ-лы, линолен қ-лы
4 -СН=СН-СН 2 -СН=СН(СН 2)7 -СООН")
Аустырылмайтың май қышқылдар Линоль қышқылы СН 3 -(СН 2)4 -СН=СН-СН 2 -СН=СН(СН 2)7 -СООН Ол арахидон қышқыл дың және арығарай простагландиндердің синтезіне қажет.

Дәрумендер Майда еритін А, Д, Е, К В 1, В 2, В 3, Н , РР , В 6, В 12, С, Р, фоль қ -лы және т. б. Суда еритін

Гиповитаминоздар Келесі дәрумендердің жетіспеушілігі жиі кездеседі : тиамин, ниацин, дәр. С, рибофлавин фоль қ-лы

Дәрумендердің Жиі кездесетін ауруларға әкеледі. Мыс. В 1 гиповитаминозда пелагра дамиді.


Сірек кездесетін гиповитаминоздар Пантотен қ-лы, пиридок син, биотин, дәрумендер В 12, Д, Е, К жетіспеушілігінде байқалады

Неорганикалық элементер • Мышьяк • кальций • хлор, хром, мыс • йод, темір

Неорганикалық элементер магний, марганец, никель, молибден, фосфор, калий, селен , кремний, натрий, ванадий, цинк және т. б.

Неорганикалық элементердің жетіспеушілігінің салдары темірдің – анемия йодтың - эндемикалық зоб кальций , фтордың – тістің аурулары және т. б.

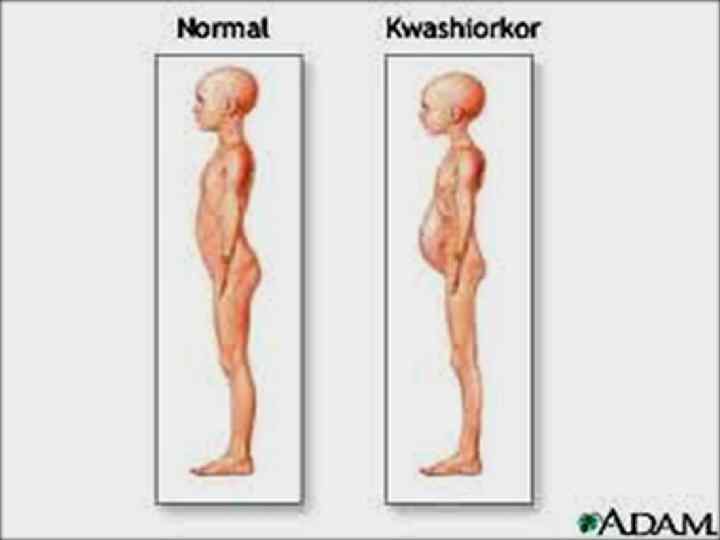
Тағамның жетіспеушілігінің салдары Ақуыздардың Квашиоркор



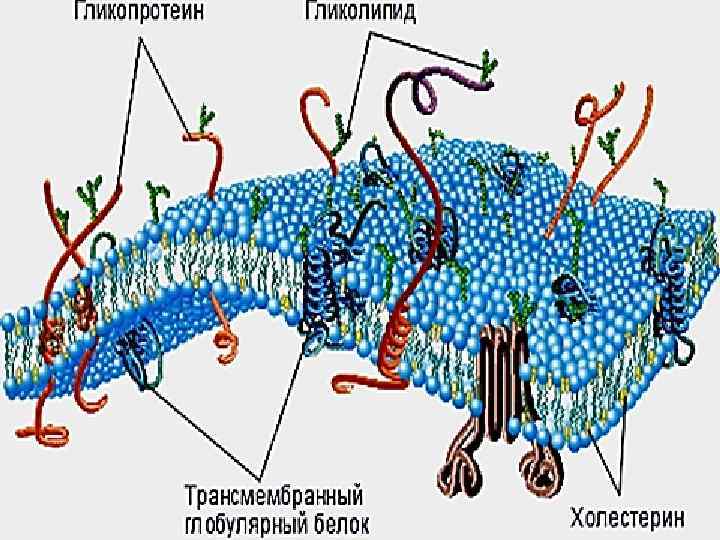
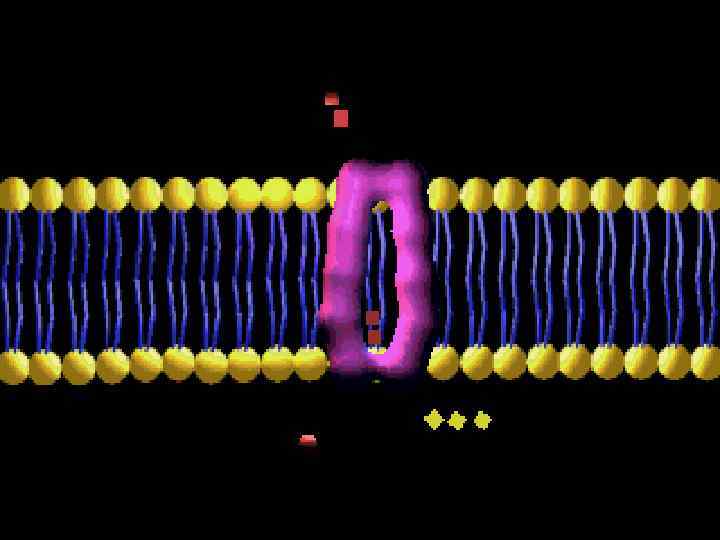
МЕМБРАНАЛАР

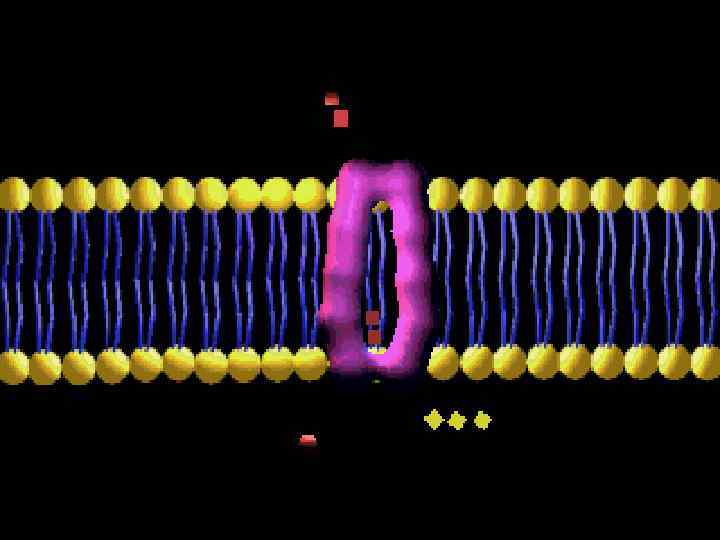
ЛИПИДТЫ БИҚАБАТ




МАКРОЭРГИЯЛЫҚ ҚОСЫЛЫСТАР

МАКРОЭРГИЯЛЫҚ ҚОСЫЛЫСТАР- ГИДРОЛИТИКА ЛЫҚ ЖОЛМЕН ЫДЫРАҒАНДА БОС ЭНЕРГИЯ

Арығарай бос энергия белгілі жұ мысқа пайдалана ды / биосинтез, зат тарды мембрана ар қылы белсенді та сымалдау және т. б. /

Ағза жасушала рында энергетикалық субстраттар ретін де көмірсулар, ли пидтер, аминқыш қылдар жатады.

Энергетикалық субстраттар дан бос энергияның болінуі катаболитикалық процестерде жүреді.

Бөліген бос энергия кейбір фосфорорга ниқалық қосылыс тардың макроэргиялық бай ланысынтарында жинақталу мүмкін.

Макроэрг. қос-ға АТФ , ГТФ, УТФ, ЦТФ, креатинфосфат, 1, 3 - дифосфоглицерат, фосфоенолпируват жатады. АТФ НЕГІЗГІ МАКРОЭРГИЯЛЫҚ ҚОСЫЛЫС

АТФ катаболизмді энергияны бөлетің анаболизммен энергияны пайдаланатың

Энергия бөліну үшін АТФ гидролизденеді. Екі жол арқылы- ортофосфатты , пирофосфатты

АТф гидролизі ортофосфатты - бос энергия бөлінеді АТФ + Н 2 О АДФ + Н 3 РО 4 + бос энергия ол жұмысқа қолданылады +

АТФ – өзінің фосфорильді тобын әртүрлі акцепторға бере алады. Бұл реакциялар киназалармен катализде неді. Фосфорланған қосылыстың энергиясы молаяды.

Осының себебінен бұл заттар ары қарай басқа ферментативті реакцияларға қатынаса алады. глюкозаның фосфорлануына Мыс. :

АТФ + глюкоза белсенді емес гексокиназа глюкоза -6 - фосфат + АДФ белсенді

АТФ-тың энергиясы актин және миозин жіпшелерінің өзара жылжуына пайдаланады. Бұл процесте: АТФ + Н 2 О АДФ + Н 2 РО 4 + + бос энергия бұлшық еттің жиырылуына

АТФ Бос энергия
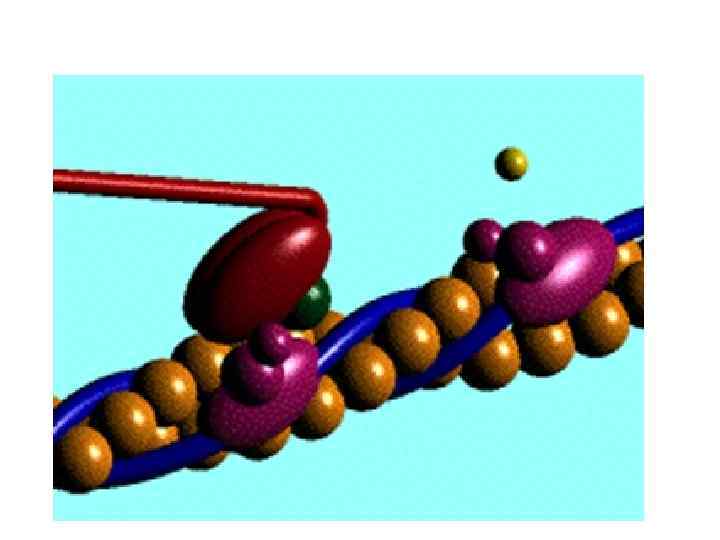

мембрана арқылы заттардың белсенді тасымалдануына

генетикалық ақпаратты беру үшін ДНҚ, РНҚ, ақуыздың биосинтезі .

АТФ-ң гидролизі пирофосфаттық АТФ + Н 2 О АМФ + Н 4 Р 2 О 7 + + бос энергия Май қышқыл. белсендіреді
 ЦТФ – (")
БАСҚА М. Қ-Ң пайдалануы УТФ – ( УДФ –глюкоза ) ЦТФ – ( ЦДФ – холин ) ГТФ - ( белок синтезі )
 д. АТФ, д. ГТФ, д.")
АТФ, ГТФ, ЦТФ, ТТФ – ( РНҚ синтезі ) д. АТФ, д. ГТФ, д. ЦТФ, д. ТТФ – ( ДНҚ синтезі )
. Креатинфосфат + АДФ креатин киназа АТФ")
Креатинфосфат (энергияның қоры, АТФ тұрақты мөлшерін қамтамасыз етеді). Креатинфосфат + АДФ креатин киназа АТФ + креатин

1, 3 – дифосфоглицерат, фосфоенолпируват – макроэргиялық қосылыстар глюкоза катаболизмінің аралық заттары болып табылады .

Олар гидролизге ұшы рамайды. Сондықтан фосфорильді тобы АДФ – ке тасымалданады ( реакцияны өзгешелік киназалар катализдейді.

Нәтижесінде АТФ түзіледі. Мыс. : фосфоенолпируват + АДФ пируваткиназа пируват + АТФ

Жасушаның биоэнергетикасы Биоэнергетика - тірі жасушаларда энергияның алмасуы және пайдалану механизмдерін оқып біледі.

Жасушаның биоэнергетикасы Молекулалардың син тезі үшін көміртектін органикалық қайнар қөздерін пайдаланатың ағзалар гетеротрофты деп аталады.

Күн сәулесінің энергиясы АТФ Бос энергия Химиялық энергия-

Термодинамиканың зандары Химиялық процестердің бағыты пайдалы энергияның ретсіз түріне аусуына сәйкес келеді. Бул аусудың өлшемі - энтропия (ΔS) деп аталады.

Энтропия жүйдегі энергия ретсіздіктің өлшемі болып табылады. Бірақ биожүйелер қоршаған ортамен
 жоғарлау тірі жүйеде емес, қоршаған ортада орын алады.")
Сондықтан энтропияның (ΔS) жоғарлау тірі жүйеде емес, қоршаған ортада орын алады.
 • Жасушада бос энергия жоғарылау немесе")
Экзергониқалық және эндергониқалық процесстер ( катаболизм, анаболизм ) • Жасушада бос энергия жоғарылау немесе төмендеу мүмкін. Себебі метаболити калық процестер бос энергияны (ΔG) бөлу немесе пайдаланумен жүреді.
 Жасушада бос энергия жоғарылау немесе төмендеу")
Экзергониқалық және эндергониқалық процесстер ( катаболизм, анаболизм ) Жасушада бос энергия жоғарылау немесе төмендеу мүмкін. Себебі метаболити калық процестер бос энергияны (ΔG) бөлу немесе пайдаланумен жүреді.
 Осымен байланысты метаболитикалық – реакциялардың 2 түрі болады экзергоникалық эндергоникалық")
) Осымен байланысты метаболитикалық – реакциялардың 2 түрі болады экзергоникалық эндергоникалық
 бөлінеді , ал энтропия (ΔS) жоғарлайды.")
Экзергоникалық реакцияларда бос энергия (ΔG) бөлінеді , ал энтропия (ΔS) жоғарлайды.
 пайдаланады немесе жиналады")
Эндергоникалық реакцияларда бос энергия (ΔG) пайдаланады немесе жиналады

Бул жағдайда химиялық заттар синтезделеді анаболизм .


• 44444 ЛЕКЦИЯ


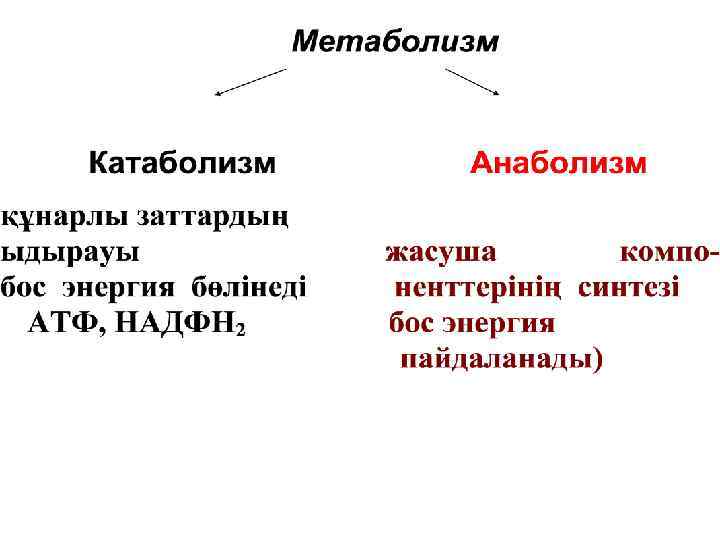
Тірі жуйелерде заттар мен энергияның алма суы катаболитика лық және анаболити калық процестердің қосындысы болып та былады.

Биожүйелерде органи калық қосылыстардың катаболизмі бос энер гияны бөлумен үлеседі. Бул процесстін негізі тотығу- тотықсыз дану реакциялар.

Тотығудың негізгі түрлері Микросомальді эндоплазматиқалық ретикулумның мембранасында Пероксидты пероксиомаларда

Оксидазды митохондрия ларда
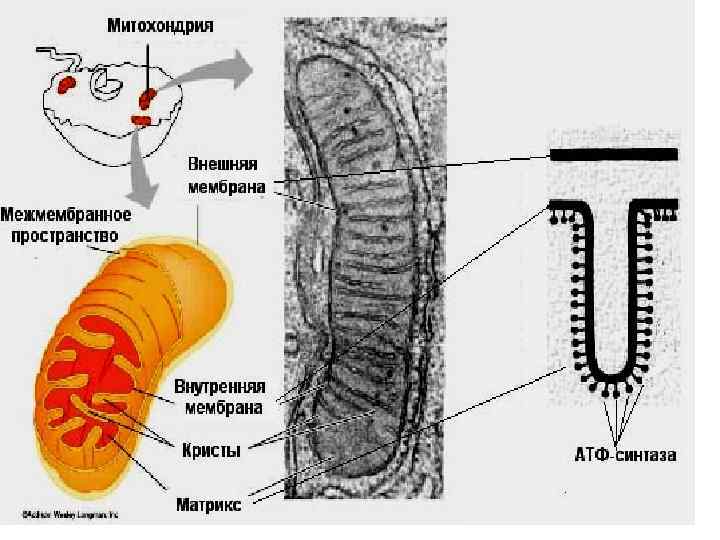

Эуокариодтердің жасу шаларында субстрат биология лық тотығуы МХң ішкі мембрана тардың сында жүреді.



Олардың коферменттері НАД, НАДФ Дәр. РР туындысы
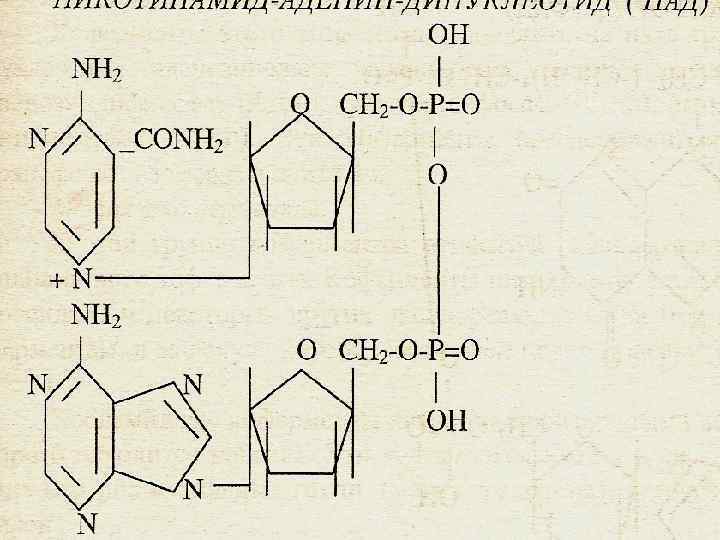

CONH 2 N CH 2 O O P P O o OH H H O H NH 2 OH O N O P O OH H H OH OH OH N N O N CH 2 OH N НАД+ H OH N O O H NH 2 N N CH 2 O H OH OH O O N H НАДФ+ OH O O H P OH OH

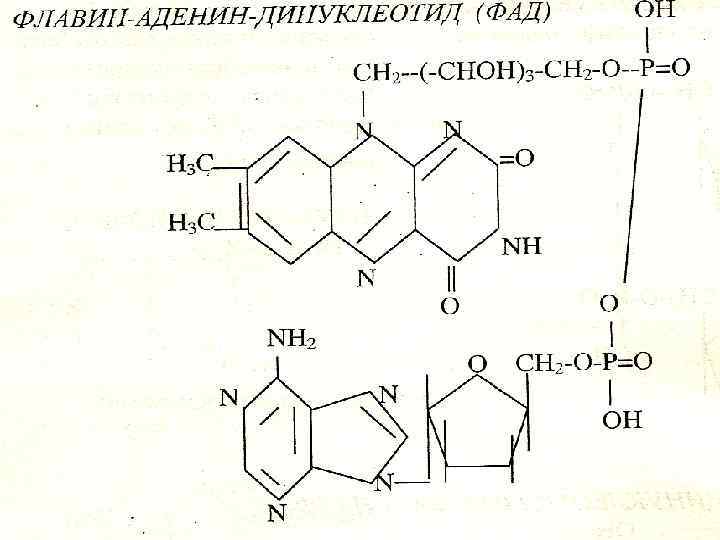

H С N С Н 3 С N С С NH Н 3 С H С С С С H 3 С С NH H 3 С С С H N N С H СН 2 O N Н ОН Н CH 2 ОН Н H C OH О N CH O OH OH ФМН О NH 2 H C OH CH 2 ОН СН 2 H C OH O P N C C N ОН О CH О N Н Р ОН О О Н Р Н СН 2 Н ФАД O
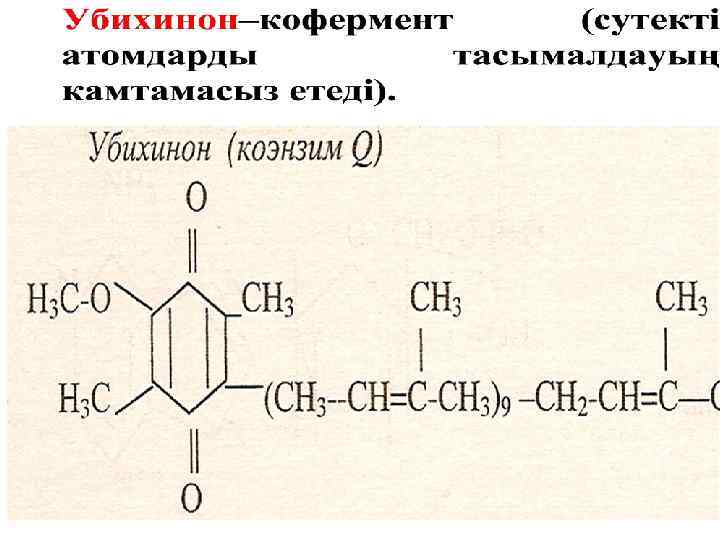

Кофермент Q О С ОH H 3 С СН 3 H 3 С С С R С O + 2 H+ + 2 е- С H 3 С СН 3 H 3 С С С R С OH

ЦИТОХРОМДАР b, c 1, c, Коферменті ++ Fe гем +++ Fe a, a 3
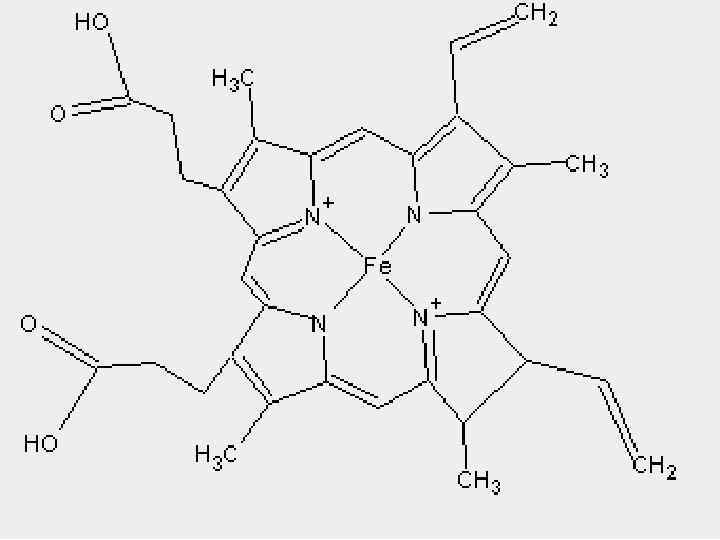

Цитохромдардың әсерінің механизмі Fe 3+ + e. Fe 2+ - e - Fe 2+ Fe 3+ Цитохромдар b , с1 фермент Ко. QH 2 дегидрогеназа

аа 3 цитохромоксидаза аа 3 электрондарды тек оттеке тасымалдайды терминальді тыныс алудың ферментті








Тыныс алу тізбекте әртүрлі субстраттардың изоцитрат, альфакетоглатарат, сукцинат, малат, лактат және т. б. тотығуы жүреді - 2 Н (2 е + 2 Н + ) .

Глицерол-3 -фосфат Ко-А-производные жирных кислот Изоцитрат ФП 4 ФП 3 сукцинат Малат ФП 2 Аскорбат Пируват АТФ Глутамат НАД АДФ 1 - ФП АТФ Ко. Q b АДФ 2 - АТФ C 1 a 3 c АДФ a 3 - Аминобарбитал, ротенон Антимицин Цианид Локализация трех пунктов фосфорилирования в цепи (1, 2, 3) О 2

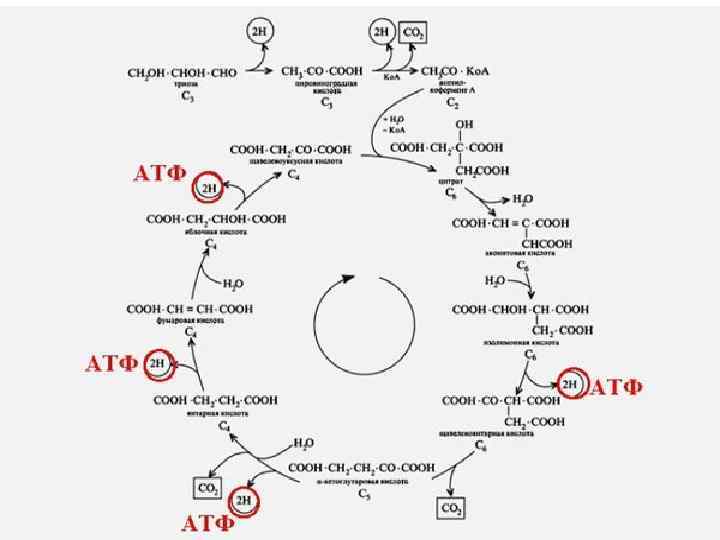
О 2 суға дейін тотық сызданады. Субстрат тың биологиялық тоты ғуында бос энергия бөлінеді (ең жоғары 3 бөлікте). Осының есе бінен 3 АТФ түзіледі

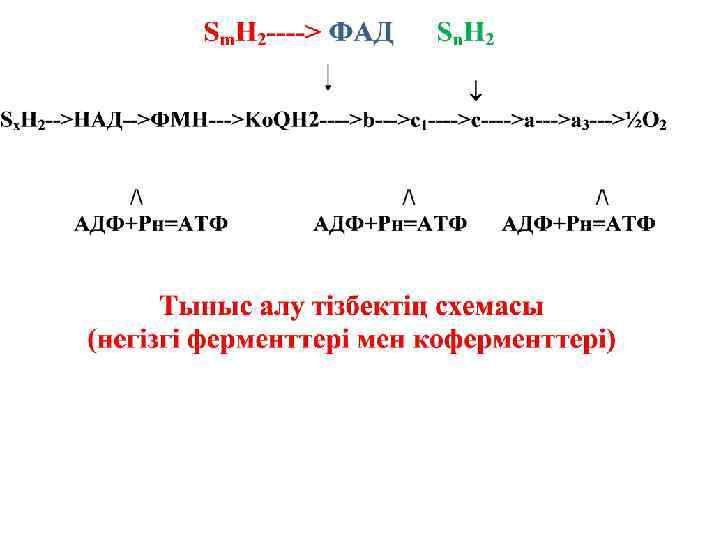
Бос энергия 3 АДФ + 3 H 3 PO 4 АТФ - синтетаза 3 АТФ АДФ-ң тотығып фосфорлануы

НАДН Sx. H 2 -2 H 2 НАД -2 H ФМНH 2 ФМН 2 H + Ko. QH 2 Ko. Q 2 e -2 H

-2 е -2 е +2/+3 Fe Fe +2/+3 -2 е Fe +2/+3

-2 е -2 е ----> +2/+3 Fe 2 - O + 2 Н + +2/+3 Fe H 2 O + бос энергия 3 АТФ Жылу
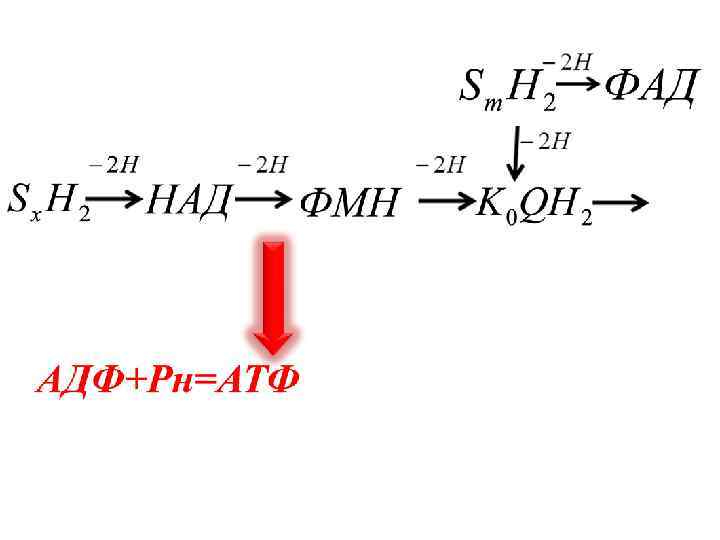

-2ē +Бос энергия 3 АТФ

Тыныс алу тізбек тін әрбір фермен тіне редокс- потенциалдың белгілі мөлшері тәң.
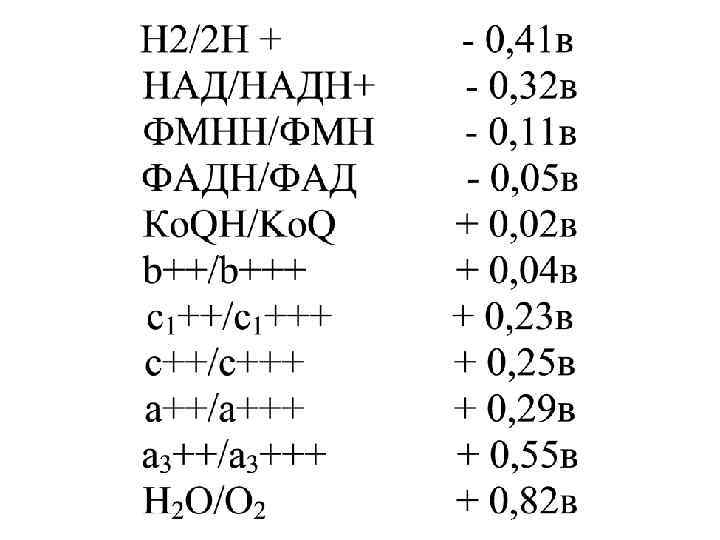
 О 2 -ң дейін (+0, 82 в) жоғарлайды.")
Ол НАД-ң (-0, 32 в) О 2 -ң дейін (+0, 82 в) жоғарлайды.

Олардың арасын дағы редокс потенциалдың ЖАЛПЫ АЙЫРМАШЫЛЫҒЫ - 1, 14 Вольт.

МХ-ғы тыныс алу тізбегінде бос энергияның мөлшерінің өзгеруі тен: ∆G = - 2 х 23062 х 1, 14 = 52, 581

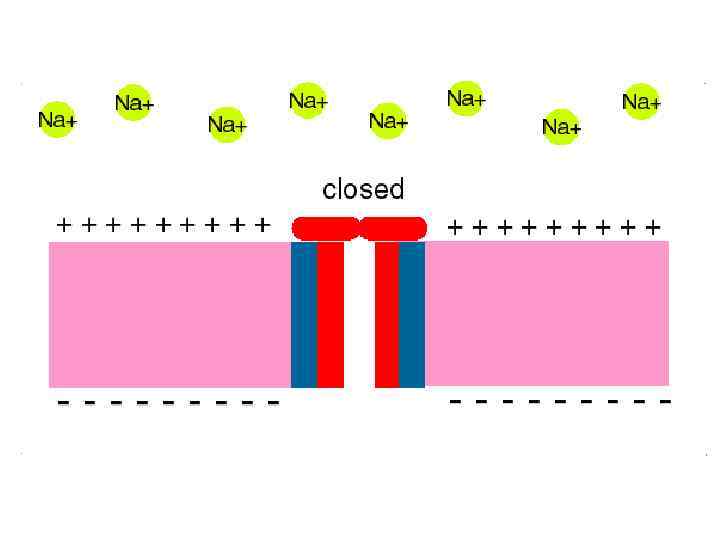
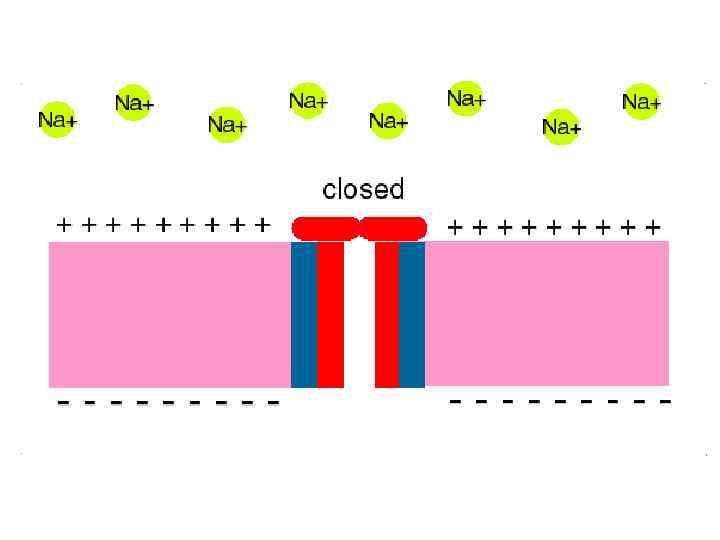
Мембранада Н+ градиентінің есебінең протонды /трансмембранды/ электрохимиялық потенциал дамиді.

Протонды/трансмембранды/ электрохимиялық потенциал Ол жасушада энергияның ерекше түрі болып табылыды.

Сыртқы орта Ішкі мембрана + + Н + Матрикс SH 2 ē + Н + + + H + + О 2 АДФ+Н 3 РО 4 + Н + - АТФ-синтетаза Н АТФ
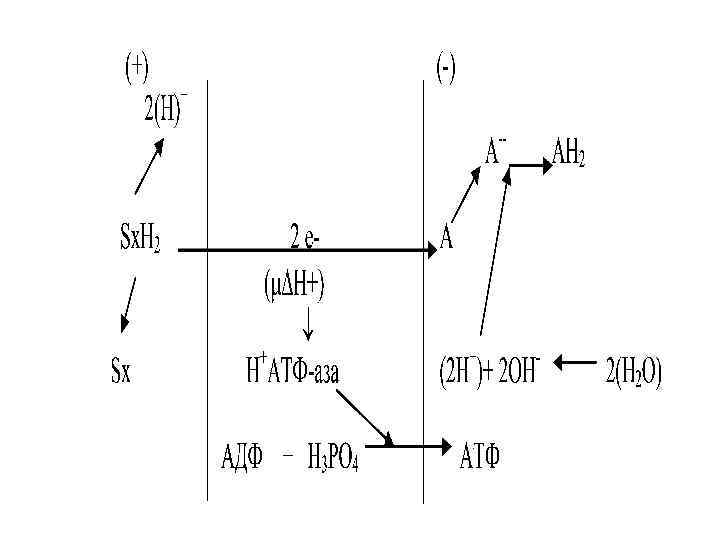
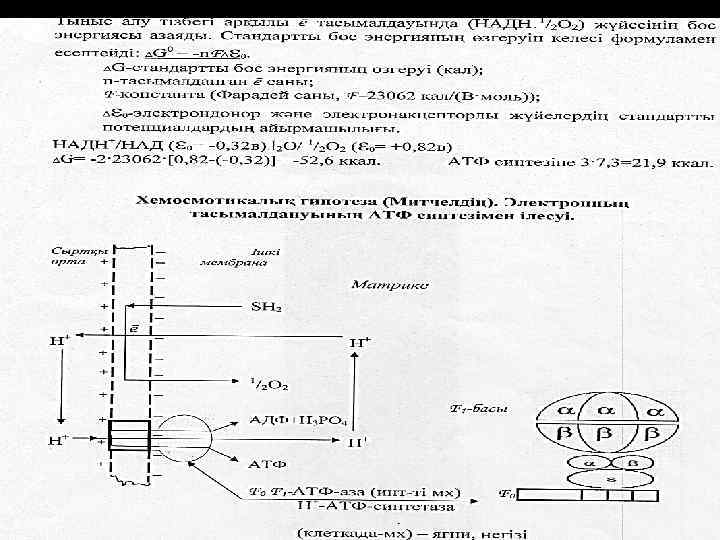

Протонды/трансмембранды/ электрохимиялық потенциал Ол жасушада энергияның ерекше түрі болып табылыды.

Осымен байланысты пайдаланады: - АТФ синтезіне / қорға жиналады- 3 2, 1 моле кула АТФ тыныс алу тізбектің ұзынды ғына байланысты

- Са 2+ және басқа иондардың, фосфат тардың тасымалда нуына - жылу энергияның пайда болуына - жиырлу үшін және т. б.


Глицерол-3 -фосфат Ко-А-производные жирных кислот Изоцитрат ФП 4 ФП 3 сукцинат Малат ФП 2 Аскорбат Пируват АТФ Глутамат НАД АДФ 1 - ФП АТФ Ко. Q b АДФ 2 - АТФ C 1 a 3 c АДФ a 3 - Аминобарбитал, ротенон Антимицин Цианид Локализация трех пунктов фосфорилирования в цепи (1, 2, 3) О 2

Затардың алмасуының кезендері


Катаболизмның өзгешелік және жалпы жолдары

Ақуыздар, көмірсулар, липидтердің катаболзмінің жалпы схемасы Көмірсулар Липидтер 1 кезен өзгешелік жолдар 2 кезен жалпы жол 3 кезен жалпы жол Пируват Ацетил-~SКо. А Кребс циклы NН 3 ( H 2 O, CO 2, n АТФ )
 СН 3 ФАД ТПФ НАД ЛК О 2 HS-Ko.")
Пируваттың тотығып декарбоксильденуі (жалпы теңдеу) СН 3 ФАД ТПФ НАД ЛК О 2 HS-Ko. A 1/2 С=О C=O + СО 2 + 3 АТФ E 1 COOH пируват СН 3 E 2 E 3 S-Ko. A ацетил- Ko. A Пируватдегидрогеназ ды комплекс
 2. Дигидролиполилацетил трансфераза ( липой")
Пируватдегидрогеназды комплекс: 1. Пируватдегидрогеназа ( ТПФ, вит. В 1) 2. Дигидролиполилацетил трансфераза ( липой қышқылы) 3. Дигидролипоилдегидро геназа ( ФАД, вит. В 2)
4 – СООH СООН S -------- S Липой")
СН 2 – CH – (СН 2)4 – СООH СООН S -------- S Липой қышқылы, тотықан түрі СН 2—CH 2 —CH —( СН 2)4 — SH SH Дигидролипой қышқылы, тотықсызданған түрі

CH 3 | | С=O + E 1 -ТДФ-H---H-С-ТДФ-E 1+CO 2 | (пируват| COOH дегидрогеназа) OH

CH 3 | S | H - C - ТПФ-Е 1+ | ЛК-Е 2 С=О+ТПФ-Е 1 | S/ S-ЛКE ОН дегидролипоилацетилтрансфераза

CH 3 | C=O | S-ЛК-E 2 + HS-Ko. A HS CH 3 ЛК-E 2 + | HS/ C=O `SKo. A

Е 3 -ФАДН 2 + НАДН 2 Е 3 -ФАД + НАДН 2 + 1/2 О 2 Н 2 О + тыныс алу тізбек 3 АТФ дигидролипоилдегидрогеназа
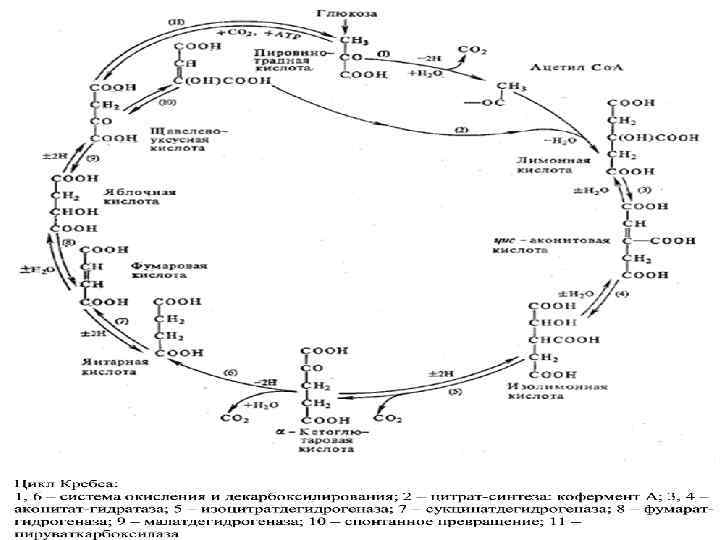

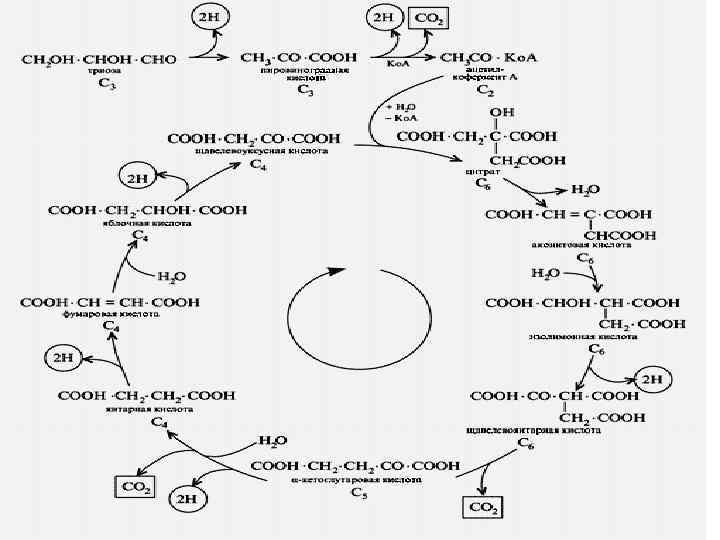
Кребс циклі СН 3 | С=О + S-Ko. A ацетил- Ko. A СООН | С=О | СН 2 цитратсинтетаза S-Ko. A / С=О | СН 2 | НО - С-СООН СН 2 СООН оксалоацетат (ЩУК) | СООН цитрил- Ko. A +НОН
 Н-С-СООН")
СООН СН 2 -НОН НО-С-СООН СН 2 СООН цитрат +НОН СООН аконитаза (Fe++) Н-С-СООН аконитаза Н- С-ОН (Fe++) СООН цис-аконитат изоцитрат СН

COOH CH 2 HC-COOH + НАД CHOH COOH ИЗОЦИТРАТ Вит РР ИЦД тын. алу тізбек 1/2 О 2 Н 2 О+ 3 АТФ CH 2 + НАДН 2 + СО 2 С О СООН α-КЕТОГЛУТАРАТ альфа-кетоглутаратдегидрогеназа

НАДН ↑ ФАД НАД ТПФ (Е 1 ЛК Е 2 АТФ СООН ГТФ / /суб фос/ СООН HS-Ko. A CH 2 Pн + ГДФ CH 2 Е 3) СН 2 сукцинаттиокиназа СН 2 С=О S-Ko. A сукцинил-Ko. A СООН сукцинат

СООН СН 2 | СН 2 ФАДН 2 ↑ ФАД Вит В 2 СООН | СН +НОН СООН НАДН 2 | ↑ НО-С-Н НАД Вит РР СООН С О СН 2 СДГ фумараза МДГ СООН СН СН 2 сукцинат СООН фумарат СООН малат СООН оксалоацетат (ЩУК)

COOH CH 2 HC - COOH -2 H С Н ОH COOH + ½O 2 НАДН 2 -2 H . . . НАД H 2 O + бос энергия 3 АТФ Жылу

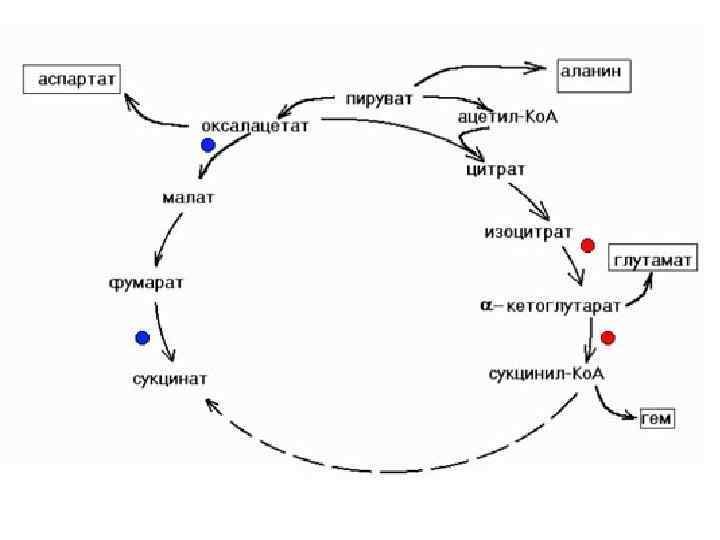
КРЕБС ЦИКЛДЫҢ БИОЛ, МӘНІ : ЭНЕРГИЯНЫҢ НЕГІЗГІ ҚАЙНАР КӨЗІ. Ацетил- Ko. A Н 2 О, СО 2 дейін тотығуыда - 12 молекула АТФ / тотығу фосфорлану -11, 1 субстратты фосфорлану / - Амфиболитикалық міндет

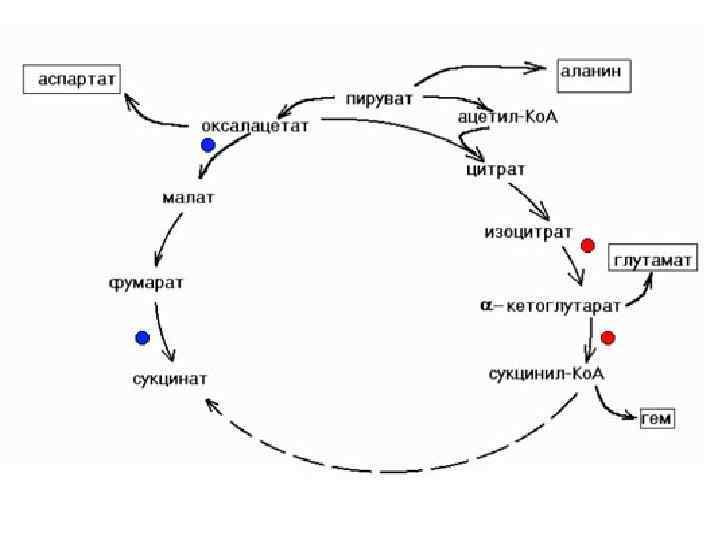

Қері байланыс • - Бос • • энергияның қайнар көзі - Энергия мен заттардың алмасуы ; - Катаболиэмның жалпы жолдары - Заттардың биологиялық тотығу - Тыныс алу тізбек

РЕФЛЕКСИВТЫ ТЕСТ • Рефлексия - субъектін өз өзіне, өз тұлғасына • /құндылықтар, қізіушіліқтар, се бептер, эмоциялар, қылықтары , • өз біліміне немесе өзінін меншікті жағдайына назар аудару

23_09_16_LAURA.ppt