

Биология.pptx
- Количество слайдов: 55
 ЗАКОНОМЕРНОСТИ НАСЛЕДСТВЕННОСТИ
ЗАКОНОМЕРНОСТИ НАСЛЕДСТВЕННОСТИ
 План Что такое генетика? Методы исследований Ученые генетики Закономерности установленные Г. Менделем Явление сцепленного наследования Наследование сцепленное с полом ü Генетические карты хромосом Как устроен геном у разных организмов? Взаимодействие 24 и более генов Взаимодействие аллельных генов I. ü ü ü Взаимодействие неаллельных генов II. ü ü Полное доминирование Неполное доминирование Кодоминантность Сверхдоминирование Множественный аллелизм Комплиментарность Эпистаз Полимерия Плейотропия
План Что такое генетика? Методы исследований Ученые генетики Закономерности установленные Г. Менделем Явление сцепленного наследования Наследование сцепленное с полом ü Генетические карты хромосом Как устроен геном у разных организмов? Взаимодействие 24 и более генов Взаимодействие аллельных генов I. ü ü ü Взаимодействие неаллельных генов II. ü ü Полное доминирование Неполное доминирование Кодоминантность Сверхдоминирование Множественный аллелизм Комплиментарность Эпистаз Полимерия Плейотропия
 – биологическая наука,") Что такое генетика? Генетика (греч. genetikоs – что относится к происхождению) – биологическая наука, предметом изучения которой есть наследственность и изменчивость. Наследственность и изменчивость – основные свойства всех живых организмов. Срок “генетика” впервые в 1906 г. предложил английский ученый В. Бетсон. v Наследственность – свойство организмов передавать свои признаки и особенности развития потомству; свойство обеспечивать материальную и функциональную преемственность между поколениями. Наследственность реализуется при размножении. Каждый вид организмов сохраняет и воссоздает себе подобное в ряде поколений. В процессе размножения воссоздается не только подобное, но возникает и новое. Дети всегда похожи на своих родителей, но никогда не бывают их точными копиями. Они отличаются как от своих родителей, так и между собою. v
Что такое генетика? Генетика (греч. genetikоs – что относится к происхождению) – биологическая наука, предметом изучения которой есть наследственность и изменчивость. Наследственность и изменчивость – основные свойства всех живых организмов. Срок “генетика” впервые в 1906 г. предложил английский ученый В. Бетсон. v Наследственность – свойство организмов передавать свои признаки и особенности развития потомству; свойство обеспечивать материальную и функциональную преемственность между поколениями. Наследственность реализуется при размножении. Каждый вид организмов сохраняет и воссоздает себе подобное в ряде поколений. В процессе размножения воссоздается не только подобное, но возникает и новое. Дети всегда похожи на своих родителей, но никогда не бывают их точными копиями. Они отличаются как от своих родителей, так и между собою. v
 Методы исследований Исследование генетики человека проходит с большими трудностями, причины которых связаны с : v невозможностью экспериментального скрещивания v медленной сменой поколений v малым количеством потомков в каждой семье v тем, что у человека сложный кариотип, большое число групп сцепления Однако, несмотря на все эти затруднения, генетика человека успешно развивается. Невозможность экспериментального скрещивания компенсируется тем, что исследователь, наблюдая обширную человеческую популяцию, может брать из тысячи брачных пар те, которые необходимы для генетического анализа. При изучении генетики человека используются следующие методы: ü генеалогический ü близнецовый ü популяционно-статистический ü дерматоглифический ü биохимический ü цитогенетический ü гибридизации соматических клеток ü моделирования
Методы исследований Исследование генетики человека проходит с большими трудностями, причины которых связаны с : v невозможностью экспериментального скрещивания v медленной сменой поколений v малым количеством потомков в каждой семье v тем, что у человека сложный кариотип, большое число групп сцепления Однако, несмотря на все эти затруднения, генетика человека успешно развивается. Невозможность экспериментального скрещивания компенсируется тем, что исследователь, наблюдая обширную человеческую популяцию, может брать из тысячи брачных пар те, которые необходимы для генетического анализа. При изучении генетики человека используются следующие методы: ü генеалогический ü близнецовый ü популяционно-статистический ü дерматоглифический ü биохимический ü цитогенетический ü гибридизации соматических клеток ü моделирования
 Генеалогический метод Этот метод основан на прослеживании какого-либо нормального или патологического признака в ряде поколений с указанием родственных связей между членами родословной. Суть этого метода состоит в том, чтобы выяснить родственные связи и проследить наличие нормального или патологического признака среди близких и дальних родственников в данной семье. Сбор сведений начинается от пробанда. Пробандом называется лицо, родословную которого необходимо составить. Им может быть больной или здоровый человек – носитель какого-либо признака или лицо, обратившееся за советом к врачу-генетику. Братья и сестры пробанда называются сибсами. Обычно родословная составляется по одному или нескольким признакам. Метод включает два этапа: I. сбор сведений о семье II. генеалогический анализ Для составления родословной проводят краткие записи о каждом члене родословной с точным указанием его родства по отношению к пробанду. Затем делают графическое изображение родословной. Чем больше имеется достоверных сведений о здоровье родственников больного, тем информативнее генеалогический метод. При собирании генетических сведений и их анализе надо иметь в виду, что признак может быть выражен в разной степени, иногда незначительной – микропризнаки.
Генеалогический метод Этот метод основан на прослеживании какого-либо нормального или патологического признака в ряде поколений с указанием родственных связей между членами родословной. Суть этого метода состоит в том, чтобы выяснить родственные связи и проследить наличие нормального или патологического признака среди близких и дальних родственников в данной семье. Сбор сведений начинается от пробанда. Пробандом называется лицо, родословную которого необходимо составить. Им может быть больной или здоровый человек – носитель какого-либо признака или лицо, обратившееся за советом к врачу-генетику. Братья и сестры пробанда называются сибсами. Обычно родословная составляется по одному или нескольким признакам. Метод включает два этапа: I. сбор сведений о семье II. генеалогический анализ Для составления родословной проводят краткие записи о каждом члене родословной с точным указанием его родства по отношению к пробанду. Затем делают графическое изображение родословной. Чем больше имеется достоверных сведений о здоровье родственников больного, тем информативнее генеалогический метод. При собирании генетических сведений и их анализе надо иметь в виду, что признак может быть выражен в разной степени, иногда незначительной – микропризнаки.
 После составления родословной начинается второй этап – генеалогический анализ, целью которого является установление генетических закономерностей: v вначале требуется установить, имеет ли признак наследственный характер; если какой-либо признак встречался в родословной несколько раз, то можно думать о его наследственной природе; однако это может быть и не так, например, какие-то внешние факторы или профессиональные вредности могут вызывать сходные заболевания у членов одной семьи v в случае обнаружения наследственного характера признака необходимо установить тип наследования: доминантный, рецессивный, сцепленный с полом Основные признаки аутосомно-доминантного наследования: проявление признака в равной мере у представителей обоих полов Ø наличие больных во всех поколениях (по вертикали) при относительно большом количестве сибсов Ø наличие больных и по горизонтали (у сестер и братьев пробанда) Ø у гетерозиготного родителя вероятность рождения больного ребенка (если второй родитель здоров) составляет 50% Следует учесть, что при доминантном типе наследования может быть пропуск в поколениях за счет слабо выраженных, «стертых» форм заболевания (малая экспрессивность мутантного гена) или за счет его низкой пенетрантности (когда у носителя донного гена признак отсутствует). Ø Основные признаки аутосомно-рецессивного наследования: относительно небольшое число больных в родословной Ø наличие больных «по горизонтали» (болеют сибсы – родные, двоюродные) Ø родители больного ребенка чаще фенотипически здоровы, но являются гетерозиготными носителями рецессивного гена Ø вероятность рождения больного ребенка составляет 25% Рецессивный признак проявляется тогда, когда в генотипе имеются оба рецессивных аллеля. При проявлении рецессивных заболеваний нередко встречается кровное родство родителей больных. Следует иметь в виду, что наличие отдаленного родства бывает неизвестно членам семьи. Приходиться учитывать косвенные соображения, например, происхождение из одного и того же малонаселенного пункта, или принадлежность к какой-либо изолированной этнической или социальной группе. Ø Основные признаки наследования, сцепленного с полом: Øзаболевания, обусловленные геном, локализованным в Х-хромосоме, могут быть как доминантными, так и рецессивными Øпри доминантном Х-сцепленном наследовании заболевание одинаково проявляется как у мужчин, так и у женщин и в дальнейшем может передаваться по потомству (в этом случае женщина может передавать этот ген половине дочерей и половине сыновей) Øпри рецессивном наследовании заболеваний, сцепленными с Х-хромосомой, как правило, страдают мужчины (гетерозиготная носительница – мать – передает мутантный ген половине сыновей, которые будут больны и половине дочерей, которые оставаясь фенотипически здоровыми, как и мать, тоже являются носительницами и передают рецессивный ген вместе с Х-хромосомой следующему поколению)
После составления родословной начинается второй этап – генеалогический анализ, целью которого является установление генетических закономерностей: v вначале требуется установить, имеет ли признак наследственный характер; если какой-либо признак встречался в родословной несколько раз, то можно думать о его наследственной природе; однако это может быть и не так, например, какие-то внешние факторы или профессиональные вредности могут вызывать сходные заболевания у членов одной семьи v в случае обнаружения наследственного характера признака необходимо установить тип наследования: доминантный, рецессивный, сцепленный с полом Основные признаки аутосомно-доминантного наследования: проявление признака в равной мере у представителей обоих полов Ø наличие больных во всех поколениях (по вертикали) при относительно большом количестве сибсов Ø наличие больных и по горизонтали (у сестер и братьев пробанда) Ø у гетерозиготного родителя вероятность рождения больного ребенка (если второй родитель здоров) составляет 50% Следует учесть, что при доминантном типе наследования может быть пропуск в поколениях за счет слабо выраженных, «стертых» форм заболевания (малая экспрессивность мутантного гена) или за счет его низкой пенетрантности (когда у носителя донного гена признак отсутствует). Ø Основные признаки аутосомно-рецессивного наследования: относительно небольшое число больных в родословной Ø наличие больных «по горизонтали» (болеют сибсы – родные, двоюродные) Ø родители больного ребенка чаще фенотипически здоровы, но являются гетерозиготными носителями рецессивного гена Ø вероятность рождения больного ребенка составляет 25% Рецессивный признак проявляется тогда, когда в генотипе имеются оба рецессивных аллеля. При проявлении рецессивных заболеваний нередко встречается кровное родство родителей больных. Следует иметь в виду, что наличие отдаленного родства бывает неизвестно членам семьи. Приходиться учитывать косвенные соображения, например, происхождение из одного и того же малонаселенного пункта, или принадлежность к какой-либо изолированной этнической или социальной группе. Ø Основные признаки наследования, сцепленного с полом: Øзаболевания, обусловленные геном, локализованным в Х-хромосоме, могут быть как доминантными, так и рецессивными Øпри доминантном Х-сцепленном наследовании заболевание одинаково проявляется как у мужчин, так и у женщин и в дальнейшем может передаваться по потомству (в этом случае женщина может передавать этот ген половине дочерей и половине сыновей) Øпри рецессивном наследовании заболеваний, сцепленными с Х-хромосомой, как правило, страдают мужчины (гетерозиготная носительница – мать – передает мутантный ген половине сыновей, которые будут больны и половине дочерей, которые оставаясь фенотипически здоровыми, как и мать, тоже являются носительницами и передают рецессивный ген вместе с Х-хромосомой следующему поколению)
 Пример генеалогического метода q - здоровый мужчина o- здоровая женщина
Пример генеалогического метода q - здоровый мужчина o- здоровая женщина
 Близнецовый метод Это один из наиболее ранних методов изучения генетики человека, однако, он не утратил своего значения и в настоящее время. Близнецовый метод был введен Ф. Гамильтоном, который выделил среди близнецов две группы: § однояйцевые (монозиготные) § двуяйцевые (дизиготные) Монозиготные близнецы при нормальном эмбриональном развитии всегда одного пола. Дизиготные близнецы рождаются чаще (2/3 общего количества двоен), они развиваются из двух одновременно созревших и оплодотворенных яйцеклеток. Такие близнецы могут быть и однополые и разнополые. С генетической точки зрения они сходны как обычные сибсы, но у них большая общность факторов среды во внутриутробном (пренатальном) и частично в постнатальном периодах. Если изучаемый признак проявляется у обоих близнецов пары, их называют конкордантными. Конкордантность – это процент сходства по изучаемому признаку. Отсутствие признака у одного из близнецов – дискордантность. Близнецовый метод используется в генетике человека для того, чтобы оценить степень влияния наследственности и среды на развитие какого-либо нормального или патологического признака. Для оценки роли наследственности в развитии того или иного признака производят расчет по формуле: Н = (% сходства ОБ - % сходства ДБ) / (100 - % сходства ДБ) где: Н- коэффициент наследственности ОБ – однояйцевые близнецы ДБ – двуяйцевые близнецы • При Н = 1 признак полностью определяется наследственным компонентом • При Н = 0 признак определяется влиянием среды • При Н = близкий к 0, 5 признак определяется примерно одинаковым влиянием наследственности и среды на формирование признака
Близнецовый метод Это один из наиболее ранних методов изучения генетики человека, однако, он не утратил своего значения и в настоящее время. Близнецовый метод был введен Ф. Гамильтоном, который выделил среди близнецов две группы: § однояйцевые (монозиготные) § двуяйцевые (дизиготные) Монозиготные близнецы при нормальном эмбриональном развитии всегда одного пола. Дизиготные близнецы рождаются чаще (2/3 общего количества двоен), они развиваются из двух одновременно созревших и оплодотворенных яйцеклеток. Такие близнецы могут быть и однополые и разнополые. С генетической точки зрения они сходны как обычные сибсы, но у них большая общность факторов среды во внутриутробном (пренатальном) и частично в постнатальном периодах. Если изучаемый признак проявляется у обоих близнецов пары, их называют конкордантными. Конкордантность – это процент сходства по изучаемому признаку. Отсутствие признака у одного из близнецов – дискордантность. Близнецовый метод используется в генетике человека для того, чтобы оценить степень влияния наследственности и среды на развитие какого-либо нормального или патологического признака. Для оценки роли наследственности в развитии того или иного признака производят расчет по формуле: Н = (% сходства ОБ - % сходства ДБ) / (100 - % сходства ДБ) где: Н- коэффициент наследственности ОБ – однояйцевые близнецы ДБ – двуяйцевые близнецы • При Н = 1 признак полностью определяется наследственным компонентом • При Н = 0 признак определяется влиянием среды • При Н = близкий к 0, 5 признак определяется примерно одинаковым влиянием наследственности и среды на формирование признака

 Метод дерматоглифики Дерматоглифика – это изучение рельефа кожи на пальцах, ладонях и подошвенных поверхностях стоп, который образован эпидермальными выступами – гребнями, которые образуют сложные узоры. Ф. Гальтон предложил классификацию этих узоров, позволившую использовать этот метод для идентификации личности в криминалистике. Разделы дерматоглифики: дактилоскопия – изучение узоров на подушечках пальцев пальмоскопия – изучение рисунка на ладонях плантоскопия – изучение дерматоглифики подошвенной поверхности стопы Дактилоскопия. Гребни на коже пальцев рук соответствуют сосочкам дермы, поэтому их называют также папиллярными линиями, рельеф этих выступов повторяет пласт эпидермиса. Межсосочковые углубления образуют бороздки. Закладка узоров происходит между 10 и 19 неделями внутриутробного развития; у 20 недельных плодов уже хорошо различимы формы узоров. Формирование папиллярного рельефа зависит от характера ветвления нервных волокон. Полное формирование деталей строения тактильных узоров отмечается к шести месяцам, после чего они остаются неизменными до конца жизни. Дерматоглифические исследования имеют важное значение в определении зиготности близнецов, в диагностике некоторых наследственных заболеваний, в судебной медицине, в криминалистике для идентификации личности. Пальмоскопия. Ладонный рельеф очень сложный, в нем выделяют ряд полей, подушечек и ладонных линий. У правшей более сложные узоры встречаются на правой руке, у левшей – на левой. Индивидуальные особенности кожных узоров наследственно обусловлены. Это доказано многими генетическими исследованиями, в частности, на монозиготных близнецах.
Метод дерматоглифики Дерматоглифика – это изучение рельефа кожи на пальцах, ладонях и подошвенных поверхностях стоп, который образован эпидермальными выступами – гребнями, которые образуют сложные узоры. Ф. Гальтон предложил классификацию этих узоров, позволившую использовать этот метод для идентификации личности в криминалистике. Разделы дерматоглифики: дактилоскопия – изучение узоров на подушечках пальцев пальмоскопия – изучение рисунка на ладонях плантоскопия – изучение дерматоглифики подошвенной поверхности стопы Дактилоскопия. Гребни на коже пальцев рук соответствуют сосочкам дермы, поэтому их называют также папиллярными линиями, рельеф этих выступов повторяет пласт эпидермиса. Межсосочковые углубления образуют бороздки. Закладка узоров происходит между 10 и 19 неделями внутриутробного развития; у 20 недельных плодов уже хорошо различимы формы узоров. Формирование папиллярного рельефа зависит от характера ветвления нервных волокон. Полное формирование деталей строения тактильных узоров отмечается к шести месяцам, после чего они остаются неизменными до конца жизни. Дерматоглифические исследования имеют важное значение в определении зиготности близнецов, в диагностике некоторых наследственных заболеваний, в судебной медицине, в криминалистике для идентификации личности. Пальмоскопия. Ладонный рельеф очень сложный, в нем выделяют ряд полей, подушечек и ладонных линий. У правшей более сложные узоры встречаются на правой руке, у левшей – на левой. Индивидуальные особенности кожных узоров наследственно обусловлены. Это доказано многими генетическими исследованиями, в частности, на монозиготных близнецах.
 Рельеф кожи на подушечках пальцев уникален для каждого человека
Рельеф кожи на подушечках пальцев уникален для каждого человека
 Биохимические методы Эти методы используются для диагностики болезней обмена веществ, причиной которых является изменение активности определенных ферментов. С помощью биохимических методов открыто около 500 молекулярных болезней, являющихся следствием проявления мутантных генов. Эти методы отличаются большой трудоемкостью, требуют специального оборудования и поэтому не могут быть широко использованы для массовых популяционных исследований с целью раннего выявления больных с наследственной патологией обмена. В последние десятилетия в разных странах разрабатываются и применяются для массовых исследований специальные программы: I. первый этап такой программы состоит в том, чтобы среди большого количества обследуемых выделить предположительно больных, имеющих какое-то наследственное отклонение от нормы, такая программа называется просеивающей, или скрининг-программой. Для этого этапа обычно используется небольшое количество простых, доступных методик (эксперессметодов). II. второй этап проводится с целью уточнения (подтверждение диагноза или отклонение при ложно-положительной реакции на первом этапе). Для этого используются точные хроматографические методы определения ферментов, аминокислот и т. п. Применяют также микробиологические тесты, они основаны на том, что некоторые штаммы бактерий могут расти только на средах, содержащих определенные аминокислоты, углеводы.
Биохимические методы Эти методы используются для диагностики болезней обмена веществ, причиной которых является изменение активности определенных ферментов. С помощью биохимических методов открыто около 500 молекулярных болезней, являющихся следствием проявления мутантных генов. Эти методы отличаются большой трудоемкостью, требуют специального оборудования и поэтому не могут быть широко использованы для массовых популяционных исследований с целью раннего выявления больных с наследственной патологией обмена. В последние десятилетия в разных странах разрабатываются и применяются для массовых исследований специальные программы: I. первый этап такой программы состоит в том, чтобы среди большого количества обследуемых выделить предположительно больных, имеющих какое-то наследственное отклонение от нормы, такая программа называется просеивающей, или скрининг-программой. Для этого этапа обычно используется небольшое количество простых, доступных методик (эксперессметодов). II. второй этап проводится с целью уточнения (подтверждение диагноза или отклонение при ложно-положительной реакции на первом этапе). Для этого используются точные хроматографические методы определения ферментов, аминокислот и т. п. Применяют также микробиологические тесты, они основаны на том, что некоторые штаммы бактерий могут расти только на средах, содержащих определенные аминокислоты, углеводы.
 Популяционно-статистический метод Этот метод позволяет изучить распространение отдельных генов в человеческих популяциях. Обычно производится непосредственное выборочное исследование части популяции либо изучают архивы больниц, родильных домов, а также проводят опрос путем анкетирования. Выбор способа зависит от цели исследования. Последний этап состоит в статистическом анализе. Одним из наиболее простых и универсальных математических методов является метод, предложенный Г. Харди и В. Вайнбергом. Имеется и ряд других специальных математических методов. В результате становится возможным определить частоту генов в различных группах населения, частоту гетерозиготных носителей ряда наследственных аномалий и болезней. Изучение распространенности генов на определенных территориях показывает, что в этом отношении их можно разделить на две категории: имеющие универсальное распространение (к их числу относится большинство известных генов) встречающиеся локально, преимущественно в определенных районах; к их числу относятся, например, ген серповидноклеточной анемии и ген, определяющий врожденный вывих бедра. Популяционно-статистический метод позволяет определить генетическую структуру популяций (соотношение между частотой гомозигот и гетерозигот). Знание генетического состава популяций имеет большое значение для социальной гигиены и профилактической медицины.
Популяционно-статистический метод Этот метод позволяет изучить распространение отдельных генов в человеческих популяциях. Обычно производится непосредственное выборочное исследование части популяции либо изучают архивы больниц, родильных домов, а также проводят опрос путем анкетирования. Выбор способа зависит от цели исследования. Последний этап состоит в статистическом анализе. Одним из наиболее простых и универсальных математических методов является метод, предложенный Г. Харди и В. Вайнбергом. Имеется и ряд других специальных математических методов. В результате становится возможным определить частоту генов в различных группах населения, частоту гетерозиготных носителей ряда наследственных аномалий и болезней. Изучение распространенности генов на определенных территориях показывает, что в этом отношении их можно разделить на две категории: имеющие универсальное распространение (к их числу относится большинство известных генов) встречающиеся локально, преимущественно в определенных районах; к их числу относятся, например, ген серповидноклеточной анемии и ген, определяющий врожденный вывих бедра. Популяционно-статистический метод позволяет определить генетическую структуру популяций (соотношение между частотой гомозигот и гетерозигот). Знание генетического состава популяций имеет большое значение для социальной гигиены и профилактической медицины.
 Благодаря популяционно-статистическому методу обнаружили , что рецессивная аллель, вызывающая альбинизм ( врожденное отсутствие пигментации кожи и радужной оболочки глаз), встречается с частотой 1 альбинос на 15000 людей с нормальной пигментацией.
Благодаря популяционно-статистическому методу обнаружили , что рецессивная аллель, вызывающая альбинизм ( врожденное отсутствие пигментации кожи и радужной оболочки глаз), встречается с частотой 1 альбинос на 15000 людей с нормальной пигментацией.
 Цитогенетический метод Принципы цитогенетических исследований сформировались в течение 20 -30 -х годов на классическом объекте генетики – дрозофиле и на некоторых растениях. Метод основан на микроскопическом исследовании хромосом. Для идентификации хромосом применяют количественный морфометрический анализ. С этой целью проводят измерение длины хромосомы в микрометрах (микроскопия хромосом производится в остановленной фазе митоза посредством колхицина и отброшенными посредством гипотонического раствора, в результате чего хромосомы лежат свободно), определяют также соотношение длины короткого плеча к длине всей хромосомы (центромерный индекс). В 1960 году была разработана первая классификация хромосом человека (Денверская). В основу ее были положены особенности величины хромосом и расположение первичной перетяжки. По форме и общим размерам все аутосомы человека подразделяются на 7 групп, обозначаемых латинскими буквами: A, B, C, D, E, F, G. Все хромосомы имеют порядковые номера. Наиболее крупная пара гомологичных хромосом имеет № 1, следующая - № 2 и т. д. Половые хромосомы - крупная X и мелкая Y – выделяются отдельно. В 1968 -1970 гг. были опубликованы работы шведского генетика Касперссона, который применил для изучения хромосом флуоресцентные красители, в частности акрихин-иприт и его производные. Последующее изучение в люминесцентном микроскопе показало, что хромосомы не дают равномерного свечения по длине. В ней выявляется несколько светящихся полос, совпадающих с локализацией структурного гетерохромтина. После удаления их хромосом ДНК они теряют почти полностью способность к флюоресценции. Если после денатурации ДНК, вызванной нагреванием и некоторыми другими факторами, провести затем ее ренатурацию – восстановление исходной двунитчатой структуры, а затем окрасить хромосомы красителем Гимзы, то в них выявляется четкая дифференцировка на темноокрашенные и светлые полосы – диски. Последовательность расположения этих дисков, их рисунок – строго специфичен для каждой хромосомы. В результате различных вариантов метода удается выявить центромерный и околоцентромерный гетерохроматин (С -диски), диски расположенные по длине хромосом (соответственно Гимзы-диски, G-диски).
Цитогенетический метод Принципы цитогенетических исследований сформировались в течение 20 -30 -х годов на классическом объекте генетики – дрозофиле и на некоторых растениях. Метод основан на микроскопическом исследовании хромосом. Для идентификации хромосом применяют количественный морфометрический анализ. С этой целью проводят измерение длины хромосомы в микрометрах (микроскопия хромосом производится в остановленной фазе митоза посредством колхицина и отброшенными посредством гипотонического раствора, в результате чего хромосомы лежат свободно), определяют также соотношение длины короткого плеча к длине всей хромосомы (центромерный индекс). В 1960 году была разработана первая классификация хромосом человека (Денверская). В основу ее были положены особенности величины хромосом и расположение первичной перетяжки. По форме и общим размерам все аутосомы человека подразделяются на 7 групп, обозначаемых латинскими буквами: A, B, C, D, E, F, G. Все хромосомы имеют порядковые номера. Наиболее крупная пара гомологичных хромосом имеет № 1, следующая - № 2 и т. д. Половые хромосомы - крупная X и мелкая Y – выделяются отдельно. В 1968 -1970 гг. были опубликованы работы шведского генетика Касперссона, который применил для изучения хромосом флуоресцентные красители, в частности акрихин-иприт и его производные. Последующее изучение в люминесцентном микроскопе показало, что хромосомы не дают равномерного свечения по длине. В ней выявляется несколько светящихся полос, совпадающих с локализацией структурного гетерохромтина. После удаления их хромосом ДНК они теряют почти полностью способность к флюоресценции. Если после денатурации ДНК, вызванной нагреванием и некоторыми другими факторами, провести затем ее ренатурацию – восстановление исходной двунитчатой структуры, а затем окрасить хромосомы красителем Гимзы, то в них выявляется четкая дифференцировка на темноокрашенные и светлые полосы – диски. Последовательность расположения этих дисков, их рисунок – строго специфичен для каждой хромосомы. В результате различных вариантов метода удается выявить центромерный и околоцентромерный гетерохроматин (С -диски), диски расположенные по длине хромосом (соответственно Гимзы-диски, G-диски).
 Болезнь Дауна вызвана дополнительной хромосомой 21 -й пары. С помощью цитогенетического метода еще на ранних этапах развития человека можно определить наличие синдрома Дауна.
Болезнь Дауна вызвана дополнительной хромосомой 21 -й пары. С помощью цитогенетического метода еще на ранних этапах развития человека можно определить наличие синдрома Дауна.
 Метод гибридизации соматических клеток Соматические клетки содержат весь объем генетической информации. Это дает возможность изучать многие вопросы генетики человека, которые невозможно исследовать на целом организме. Благодаря методам генетики соматических клеток человек как бы стал одним из экспериментальных объектов. Чаще всего используют клетки соединительной ткани (фибробласты) и лимфоциты крови. Культивирование клеток вне организма позволяет получить достаточное количество материала для исследования, что не всегда возможно взять у человека без ущерба для здоровья. В 1960 г. французский биолог Ж. Барский, выращивая вне организма в культуре ткани клетки двух линий мышей, обнаружил, что некоторые клетки по своим морфологическим и биохимическим признакам были промежуточными между исходными родительскими клетками. Эти клетки оказались гибридными. Такое спонтанное слияние клеток в культуре ткани происходит довольно редко. В дальнейшем оказалось, что частота гибридизации соматических клеток повышается при введении в культуру клеток РНК-содержащего вируса парагриппа Сендай, который как вообще все вирусы, изменяет свойства клеточных мембран и делает возможным слияние клеток. Под влиянием такого вируса в смешанной культуре двух типов клеток образуются клетки, содержащие в общей цитоплазме ядра обеих родительских клеток – гетерокарионы. После митоза и последующего разделения цитоплазмы из двуядерного гетерокариона образуются две одноядерные клетки, каждая из которых представляет собой синкарион – настоящую гибридную клетку, имеющую хромосомы обеих родительских клеток.
Метод гибридизации соматических клеток Соматические клетки содержат весь объем генетической информации. Это дает возможность изучать многие вопросы генетики человека, которые невозможно исследовать на целом организме. Благодаря методам генетики соматических клеток человек как бы стал одним из экспериментальных объектов. Чаще всего используют клетки соединительной ткани (фибробласты) и лимфоциты крови. Культивирование клеток вне организма позволяет получить достаточное количество материала для исследования, что не всегда возможно взять у человека без ущерба для здоровья. В 1960 г. французский биолог Ж. Барский, выращивая вне организма в культуре ткани клетки двух линий мышей, обнаружил, что некоторые клетки по своим морфологическим и биохимическим признакам были промежуточными между исходными родительскими клетками. Эти клетки оказались гибридными. Такое спонтанное слияние клеток в культуре ткани происходит довольно редко. В дальнейшем оказалось, что частота гибридизации соматических клеток повышается при введении в культуру клеток РНК-содержащего вируса парагриппа Сендай, который как вообще все вирусы, изменяет свойства клеточных мембран и делает возможным слияние клеток. Под влиянием такого вируса в смешанной культуре двух типов клеток образуются клетки, содержащие в общей цитоплазме ядра обеих родительских клеток – гетерокарионы. После митоза и последующего разделения цитоплазмы из двуядерного гетерокариона образуются две одноядерные клетки, каждая из которых представляет собой синкарион – настоящую гибридную клетку, имеющую хромосомы обеих родительских клеток.
 Ученые-генетики Большой вклад в исследования закономерностей наследственности внесли такие ученые как : • Грегор Мендель Томас Хант Морган • Гуго де Фриз • Карл Корренс • Иоган Чермак •
Ученые-генетики Большой вклад в исследования закономерностей наследственности внесли такие ученые как : • Грегор Мендель Томас Хант Морган • Гуго де Фриз • Карл Корренс • Иоган Чермак •
 Грегор Мендель Родился 20 июля 1822 года в Австралии. Он достиг успеха в своих экспериментах благодаря использованию гибридологического метода —скрещивания организмов, различающихся по каким-либо признакам, и анализа всех последующих поколений с целью установления закономерностей наследования этих признаков Г. Мендель усовершенствовал данный метод, и в отличие от своих предшественников, анализировал наследование ограниченного количества признаков (одного, двух, трех). При этом он выбирал признак с альтернативным(контрастирующим) проявлением его у скрещиваемых организмов. Так, он скрещивал сорта гороха с окрашенными и белыми цветками, гладкими и морщинистыми семенами и т. п. Кроме того, Мендель проверял перед скрещиванием, насколько устойчиво наследуются выбранные им признаки в ряду поколений при самоопылении.
Грегор Мендель Родился 20 июля 1822 года в Австралии. Он достиг успеха в своих экспериментах благодаря использованию гибридологического метода —скрещивания организмов, различающихся по каким-либо признакам, и анализа всех последующих поколений с целью установления закономерностей наследования этих признаков Г. Мендель усовершенствовал данный метод, и в отличие от своих предшественников, анализировал наследование ограниченного количества признаков (одного, двух, трех). При этом он выбирал признак с альтернативным(контрастирующим) проявлением его у скрещиваемых организмов. Так, он скрещивал сорта гороха с окрашенными и белыми цветками, гладкими и морщинистыми семенами и т. п. Кроме того, Мендель проверял перед скрещиванием, насколько устойчиво наследуются выбранные им признаки в ряду поколений при самоопылении.
 Томас Хант Морган Родился 25 сентября 1866 года в США. С 1910 изучал наследование мутаций, обнаруженных у нового генетического объекта — плодовой мухи дрозофилы, в результате чего экспериментально обосновал (совместно с А. Стёртевантом, Г. Мёллером и К. Бриджесом) представления о материальных основах наследственности (корпускулярную природу генетического материала — генов, линейную локализацию генов в хромосомах, закономерности их мутационной изменчивости. цитогенетические механизмы их наследственной передачи и др. ), приведшие к окончательному доказательству и завершению в основных чертах хромосомной теории наследственности. Установленные М. и его сотрудниками закономерности сцепления генов и кроссинговера (иногда называют законом М. , или морганизмом) полностью разъяснили цитологический механизм Менделя законов и послужили стимулом к разработке генетических основ теории естественного отбора
Томас Хант Морган Родился 25 сентября 1866 года в США. С 1910 изучал наследование мутаций, обнаруженных у нового генетического объекта — плодовой мухи дрозофилы, в результате чего экспериментально обосновал (совместно с А. Стёртевантом, Г. Мёллером и К. Бриджесом) представления о материальных основах наследственности (корпускулярную природу генетического материала — генов, линейную локализацию генов в хромосомах, закономерности их мутационной изменчивости. цитогенетические механизмы их наследственной передачи и др. ), приведшие к окончательному доказательству и завершению в основных чертах хромосомной теории наследственности. Установленные М. и его сотрудниками закономерности сцепления генов и кроссинговера (иногда называют законом М. , или морганизмом) полностью разъяснили цитологический механизм Менделя законов и послужили стимулом к разработке генетических основ теории естественного отбора
 Гуго де Фриз Родился 16 февраля 1848 года в Нидерландах. Научную деятельность Де Фриз начал с изучения роли осмотического давления в жизнедеятельности растительной клетки. Исследуя явление плазмолиза (сокращения клеток в растворе, концентрация которого выше концентрации их содержимого), разработал метод определения осмотического давления в клетке Де Фриз придерживался теории внутриклеточного пангенеза. Он считал, что в ядре клетки содержатся «пангены» *, определяющие все признаки целого организма, а в протоплазму входят лишь те «пангены» , которые определяют тип клеток. Тем самым отвергался принцип Ламарка о наследовании приобретенных признаков. Кроме того, Де Фриз сделал важный вывод, что панген вообще не является клеткой или органом, а представляет собой некий фактор, определяющий признак, т. е. уже в 1889 вплотную приблизился к точке зрения Менделя. Другая важная идея Де Фриза заключалась в том, что факторы наследственности не могут трансформироваться постепенно, а претерпевают резкие скачкообразные изменения. Изучая изменчивость энотеры (растения из семейства кипрейных), ученый пришел к выводу, что вид может внезапно распасться на несколько новых видов. Этот феномен Де Фриз назвал мутацией. *Пангены- по теории Дарвина это невидимые мельчайшие частицы, которые перемещаются от места своего появления к половым клеткам, частью которых они становятся.
Гуго де Фриз Родился 16 февраля 1848 года в Нидерландах. Научную деятельность Де Фриз начал с изучения роли осмотического давления в жизнедеятельности растительной клетки. Исследуя явление плазмолиза (сокращения клеток в растворе, концентрация которого выше концентрации их содержимого), разработал метод определения осмотического давления в клетке Де Фриз придерживался теории внутриклеточного пангенеза. Он считал, что в ядре клетки содержатся «пангены» *, определяющие все признаки целого организма, а в протоплазму входят лишь те «пангены» , которые определяют тип клеток. Тем самым отвергался принцип Ламарка о наследовании приобретенных признаков. Кроме того, Де Фриз сделал важный вывод, что панген вообще не является клеткой или органом, а представляет собой некий фактор, определяющий признак, т. е. уже в 1889 вплотную приблизился к точке зрения Менделя. Другая важная идея Де Фриза заключалась в том, что факторы наследственности не могут трансформироваться постепенно, а претерпевают резкие скачкообразные изменения. Изучая изменчивость энотеры (растения из семейства кипрейных), ученый пришел к выводу, что вид может внезапно распасться на несколько новых видов. Этот феномен Де Фриз назвал мутацией. *Пангены- по теории Дарвина это невидимые мельчайшие частицы, которые перемещаются от места своего появления к половым клеткам, частью которых они становятся.
 Карл Корренс и Эрих Чермак Основная заслуга Корренса и Эриха - вторичное открытие и подтверждение (одновременно с Г. Де Фризом и Э. Чермаком) законов наследственности, установленных Г. Менделем. Карл Корренс родился в Мюнхене 19 сентября 1864 года. Труды Корренса и Эриха посвящены дальнейшему изучению явлении наследственности у растений: ксений, определению по ла, пестролистности и плазматической наследственности. Корренс предвосхитил понимание закономерностей сцепления и обмена наследственных факторов в хромосомах (1902) и менделевского наследования пола у растений. Эрих Чермак родился 15 ноября 1871 года в Праге
Карл Корренс и Эрих Чермак Основная заслуга Корренса и Эриха - вторичное открытие и подтверждение (одновременно с Г. Де Фризом и Э. Чермаком) законов наследственности, установленных Г. Менделем. Карл Корренс родился в Мюнхене 19 сентября 1864 года. Труды Корренса и Эриха посвящены дальнейшему изучению явлении наследственности у растений: ксений, определению по ла, пестролистности и плазматической наследственности. Корренс предвосхитил понимание закономерностей сцепления и обмена наследственных факторов в хромосомах (1902) и менделевского наследования пола у растений. Эрих Чермак родился 15 ноября 1871 года в Праге
 Закономерности установленные Г. Менделем 1. Закон единообразия гибридов первого поколения При скрещивании двух гомозиготных организмов, относящихся к разным чистым линиям и отличающихся друг от друга по одной паре альтернативных признаков, всё первое поколение гибридов (F 1) окажется единообразным и будет нести признак одного из родителей 2. Закон расщепления признаков Закон расщепления, или второй закон Менделя: при скрещивании двух гетерозиготных потомков первого поколения между собой во втором поколении наблюдается расщепление в определенном числовом отношении: по фенотипу 3: 1, по генотипу 1: 2: 1. 3. Закон независимого наследования признаков Закон независимого наследования (третий закон Менделя) — при скрещивании двух гомозиготных особей, отличающихся друг от друга по двум (и более) парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании). Когда скрещивались растения, отличающиеся по нескольким признакам, таким как белые и пурпурные цветы и желтые или зелёные
Закономерности установленные Г. Менделем 1. Закон единообразия гибридов первого поколения При скрещивании двух гомозиготных организмов, относящихся к разным чистым линиям и отличающихся друг от друга по одной паре альтернативных признаков, всё первое поколение гибридов (F 1) окажется единообразным и будет нести признак одного из родителей 2. Закон расщепления признаков Закон расщепления, или второй закон Менделя: при скрещивании двух гетерозиготных потомков первого поколения между собой во втором поколении наблюдается расщепление в определенном числовом отношении: по фенотипу 3: 1, по генотипу 1: 2: 1. 3. Закон независимого наследования признаков Закон независимого наследования (третий закон Менделя) — при скрещивании двух гомозиготных особей, отличающихся друг от друга по двум (и более) парам альтернативных признаков, гены и соответствующие им признаки наследуются независимо друг от друга и комбинируются во всех возможных сочетаниях (как и при моногибридном скрещивании). Когда скрещивались растения, отличающиеся по нескольким признакам, таким как белые и пурпурные цветы и желтые или зелёные
 Явление сцепленного наследования Т. Морган установил некоторые исключения из третьего закона Менделя. Он выяснил, что иногда два или несколько признаков не дают в потомстве независимого расщепления. Признаки оказывается сцеплены между собой, наследуются совместно. Причина этого явления в том, что в любом организме значительно меньше хромосом, чем генов. Например, у человека в каждой клетке 23 пары хромосом, а генов - много тысяч. Хромосома наследуется как единое целое, поскольку целиком оказывается в половых клетках. Следовательно, расположенные в одной хромосоме гены, как правило, наследуется совместно. Открытое Морганом явление носит название сцепленного наследования. Из правила, установленного Морганом, в свою очередь, встречаются исключения. Эти исключения возникают благодаря процессу кроссинговера. В ходе этого процесса параллельные хромосомы обмениваются генами, и сцепленные гены процесса параллельны хромосомы обмениваются генами, и специальные гены могут быть "расцеплены". Кроссинговер чрезвычайно полезен для выживания вида в ходе эволюции, так как создаёт новые сочетания наследственных
Явление сцепленного наследования Т. Морган установил некоторые исключения из третьего закона Менделя. Он выяснил, что иногда два или несколько признаков не дают в потомстве независимого расщепления. Признаки оказывается сцеплены между собой, наследуются совместно. Причина этого явления в том, что в любом организме значительно меньше хромосом, чем генов. Например, у человека в каждой клетке 23 пары хромосом, а генов - много тысяч. Хромосома наследуется как единое целое, поскольку целиком оказывается в половых клетках. Следовательно, расположенные в одной хромосоме гены, как правило, наследуется совместно. Открытое Морганом явление носит название сцепленного наследования. Из правила, установленного Морганом, в свою очередь, встречаются исключения. Эти исключения возникают благодаря процессу кроссинговера. В ходе этого процесса параллельные хромосомы обмениваются генами, и сцепленные гены процесса параллельны хромосомы обмениваются генами, и специальные гены могут быть "расцеплены". Кроссинговер чрезвычайно полезен для выживания вида в ходе эволюции, так как создаёт новые сочетания наследственных
 Явление сцепленного наследования В 1911 — 1912 годах Т. Морган и сотрудники проверили проявление третьего закона Менделя на мухах-дрозофилах. Они учитывали две пары альтернативных признаков: серый (В) и черный (b) цвет тела и нормальные (V) и короткие (v) крылья. При скрещивании гомозиготных особей с серым цветом тела и нормальными крыльями с мухами с черным цветом тела и короткими крыльями получили единообразие гибридов первого поколения — мух с серым телом и нормальными крыльями. Подтвердился I закон Менделя.
Явление сцепленного наследования В 1911 — 1912 годах Т. Морган и сотрудники проверили проявление третьего закона Менделя на мухах-дрозофилах. Они учитывали две пары альтернативных признаков: серый (В) и черный (b) цвет тела и нормальные (V) и короткие (v) крылья. При скрещивании гомозиготных особей с серым цветом тела и нормальными крыльями с мухами с черным цветом тела и короткими крыльями получили единообразие гибридов первого поколения — мух с серым телом и нормальными крыльями. Подтвердился I закон Менделя.
 Далее Морган решил провести анализирующее скрещивание гибридов первого поколения. Рецессивную гомозиготную самку он скрестил с дигетерозиготным самцом. Морган ожидал получить, согласно третьему закону Менделя, мух четырех разных фенотипов в равном количестве (по 25%), а получил двух фенотипов (по 50% каждого). Морган пришел к выводу, что поскольку у организмов генов много, а хромосом относительно мало, то, следовательно, в каждой хромосоме содержится большое количество генов, и гены, локализованные в одной хромосоме, передаются вместе (сцепленно). Цитологические основы этого явления можно пояснить следующей схемой. Одна из пары гомологичных хромосом содержит два доминантных гена (BV), а другая — два рецессивных (bv). При мейозе хромосома с генами BV попадет в одну гамету, а хромосома с генами bv в другую.
Далее Морган решил провести анализирующее скрещивание гибридов первого поколения. Рецессивную гомозиготную самку он скрестил с дигетерозиготным самцом. Морган ожидал получить, согласно третьему закону Менделя, мух четырех разных фенотипов в равном количестве (по 25%), а получил двух фенотипов (по 50% каждого). Морган пришел к выводу, что поскольку у организмов генов много, а хромосом относительно мало, то, следовательно, в каждой хромосоме содержится большое количество генов, и гены, локализованные в одной хромосоме, передаются вместе (сцепленно). Цитологические основы этого явления можно пояснить следующей схемой. Одна из пары гомологичных хромосом содержит два доминантных гена (BV), а другая — два рецессивных (bv). При мейозе хромосома с генами BV попадет в одну гамету, а хромосома с генами bv в другую.
 Таким образом, у дигетерозиготного организма образуются не четыре типа гамет (когда гены расположены в разных хромосомах), а только два, и, следовательно, потомки будут иметь два сочетания признаков (как у родителей). Гены, локализованные в одной хромосоме, обычно передаются вместе и составляют одну группу сцепления. Так как в гомологичных хромосомах локализованы аллельные гены, то группу сцепления составляют две гомологичные хромосомы, и, следовательно, количество групп сцепления соответствует количеству пар хромосом (или гаплоидному числу хромосом). Так, у мухи-дрозофилы всего 8 хромосом — 4 труппы сцепления, у человека 46 хромосом — 23 группы сцепления. Если гены, локализованные в одной хромосоме, передаются всегда вместе, то такое сцепление называется полным. Однако при дальнейшем анализе сцепления генов было обнаружено, что в некоторых случаях оно может нарушаться. Если дигетерозиготную самку мухи-дрозофилы скрестить с рецессивным самцом, результат будет следующий:
Таким образом, у дигетерозиготного организма образуются не четыре типа гамет (когда гены расположены в разных хромосомах), а только два, и, следовательно, потомки будут иметь два сочетания признаков (как у родителей). Гены, локализованные в одной хромосоме, обычно передаются вместе и составляют одну группу сцепления. Так как в гомологичных хромосомах локализованы аллельные гены, то группу сцепления составляют две гомологичные хромосомы, и, следовательно, количество групп сцепления соответствует количеству пар хромосом (или гаплоидному числу хромосом). Так, у мухи-дрозофилы всего 8 хромосом — 4 труппы сцепления, у человека 46 хромосом — 23 группы сцепления. Если гены, локализованные в одной хромосоме, передаются всегда вместе, то такое сцепление называется полным. Однако при дальнейшем анализе сцепления генов было обнаружено, что в некоторых случаях оно может нарушаться. Если дигетерозиготную самку мухи-дрозофилы скрестить с рецессивным самцом, результат будет следующий:
 Морган предполагал получить опять мух четырех фенотипов по 25%, а получил потомков четырех фенотипов, но в другом соотношении: по 41, 5% особей с серым телом и нормальными крыльями и с черным телом и короткими крыльями и по 8, 5% мух с серым телом и короткими крыльями и с черным телом и нормальными крыльями. В этом случае сцепление генов неполное, т. е. гены, локализованные в одной хромосоме, не всегда передаются вместе. Это связано с явлением кроссинговера, которое заключается в обмене участками гомологичных хроматид в процессе их конъюгации в профазе мейоза I. Кроссинговер у гетерозиготных организмов приводит к перекомбинации генетического материала. Схема Кроссинговера
Морган предполагал получить опять мух четырех фенотипов по 25%, а получил потомков четырех фенотипов, но в другом соотношении: по 41, 5% особей с серым телом и нормальными крыльями и с черным телом и короткими крыльями и по 8, 5% мух с серым телом и короткими крыльями и с черным телом и нормальными крыльями. В этом случае сцепление генов неполное, т. е. гены, локализованные в одной хромосоме, не всегда передаются вместе. Это связано с явлением кроссинговера, которое заключается в обмене участками гомологичных хроматид в процессе их конъюгации в профазе мейоза I. Кроссинговер у гетерозиготных организмов приводит к перекомбинации генетического материала. Схема Кроссинговера
 Наследование, сцепленное с полом Пол организма — это совокупность признаков и анатомических структур, обеспечивающих половой путь размножения и передачу наследственной информации. В определении пола будущей особи ведущую роль играет хромосомный аппарат зиготы — кариотип. Различают хромосомы, одинаковые для обоих полов — аутосомы, и половые хромосомы. В кариотипе человека содержится 44 аутосомы и 2 половых хромосомы — Х и Y. За развитие женского пола у человека отвечают две Х-хромосомы, т. е. женский пол гомогаметен. Развитие мужского пола определяется наличием Х- и Y-хромосом, т. е. мужской пол гетерогаметен. Сочетание половых хромосом в зиготе определяет пол будущего организма ( У всех млекопитающих, человека и мухи-дрозофилы, гомогаметным является женский пол, а гетерогаметным — мужской. У птиц и бабочек, наоборот, гомогаметен мужской пол, а женский — гетерогаметен. Признаки, сцепленные с полом Это признаки, которые кодируются генами, находящимися на половых хромосомах. У человека признаки, кодируемые генами Х-хромосомы, могут проявляться у представителей обоих полов, а кодируемые генами Y-хромосомы — только у мужчин. Следует иметь в виду, что в мужском генотипе только одна Х-хромосома, которая почти не содержит участков, гомологичных с Y-хромосомой, поэтому все локализованные в Х-хромосоме гены, в том числе и рецессивные, проявляются в фенотипе в первом же поколении. В половых хромосомах содержатся гены, регулирующие проявление не только половых признаков. Ххромосома имеет гены, отвечающие за свертываемость крови, цветовое восприятие, синтез ряда ферментов. В Y-хромосоме содержится ряд генов, контролирующих признаки, наследуемые по мужской линии (голандрические признаки): волосистость ушной раковины, наличие кожной перепонки между пальцами и др. Известно очень мало генов, общих для Х- и Y-хромосом. Различают Х-сцепленное и Y-сцепленное (голандрическое) наследование.
Наследование, сцепленное с полом Пол организма — это совокупность признаков и анатомических структур, обеспечивающих половой путь размножения и передачу наследственной информации. В определении пола будущей особи ведущую роль играет хромосомный аппарат зиготы — кариотип. Различают хромосомы, одинаковые для обоих полов — аутосомы, и половые хромосомы. В кариотипе человека содержится 44 аутосомы и 2 половых хромосомы — Х и Y. За развитие женского пола у человека отвечают две Х-хромосомы, т. е. женский пол гомогаметен. Развитие мужского пола определяется наличием Х- и Y-хромосом, т. е. мужской пол гетерогаметен. Сочетание половых хромосом в зиготе определяет пол будущего организма ( У всех млекопитающих, человека и мухи-дрозофилы, гомогаметным является женский пол, а гетерогаметным — мужской. У птиц и бабочек, наоборот, гомогаметен мужской пол, а женский — гетерогаметен. Признаки, сцепленные с полом Это признаки, которые кодируются генами, находящимися на половых хромосомах. У человека признаки, кодируемые генами Х-хромосомы, могут проявляться у представителей обоих полов, а кодируемые генами Y-хромосомы — только у мужчин. Следует иметь в виду, что в мужском генотипе только одна Х-хромосома, которая почти не содержит участков, гомологичных с Y-хромосомой, поэтому все локализованные в Х-хромосоме гены, в том числе и рецессивные, проявляются в фенотипе в первом же поколении. В половых хромосомах содержатся гены, регулирующие проявление не только половых признаков. Ххромосома имеет гены, отвечающие за свертываемость крови, цветовое восприятие, синтез ряда ферментов. В Y-хромосоме содержится ряд генов, контролирующих признаки, наследуемые по мужской линии (голандрические признаки): волосистость ушной раковины, наличие кожной перепонки между пальцами и др. Известно очень мало генов, общих для Х- и Y-хромосом. Различают Х-сцепленное и Y-сцепленное (голандрическое) наследование.
 Х-сцепленное наследование Так как Х-хромосома присутствует в кариотипе каждого человека, то и признаки, наследуемые сцеплено с Х-хромосомой, проявляются у представителей обоих полов. Женщины получают эти гены от обоих родителей и через свои гаметы передают их потомкам. Мужчины получают Х-хромосому от матери и передают ее своему потомству женского пола. Различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. У человека Х-сцепленный доминантный признак передается матерью всему потомству. Мужчина передает свой Х-сцепленный доминантный признак лишь своим дочерям. Х-сцепленный рецессивный признак у женщин проявляется лишь при получении ими соответствующего аллеля от обоих родителей. У мужчин он развивается при получении рецессивного аллеля от матери. Женщины передают рецессивный аллель потомкам обоих полов, а мужчины — только дочерям. При Х-сцепленном наследовании возможен промежуточный характер проявления признака у гетерозигот. Y-сцепленные гены присутствуют в генотипе только мужчин и передаются из поколения в поколение от отца к сыну.
Х-сцепленное наследование Так как Х-хромосома присутствует в кариотипе каждого человека, то и признаки, наследуемые сцеплено с Х-хромосомой, проявляются у представителей обоих полов. Женщины получают эти гены от обоих родителей и через свои гаметы передают их потомкам. Мужчины получают Х-хромосому от матери и передают ее своему потомству женского пола. Различают Х-сцепленное доминантное и Х-сцепленное рецессивное наследование. У человека Х-сцепленный доминантный признак передается матерью всему потомству. Мужчина передает свой Х-сцепленный доминантный признак лишь своим дочерям. Х-сцепленный рецессивный признак у женщин проявляется лишь при получении ими соответствующего аллеля от обоих родителей. У мужчин он развивается при получении рецессивного аллеля от матери. Женщины передают рецессивный аллель потомкам обоих полов, а мужчины — только дочерям. При Х-сцепленном наследовании возможен промежуточный характер проявления признака у гетерозигот. Y-сцепленные гены присутствуют в генотипе только мужчин и передаются из поколения в поколение от отца к сыну.
 Половина сперматозоидов несет X-хромосому, а другая половина — Y-хромосому. Пол ребенка зависит от того, какой сперматозоид оплодотворит яйцеклетку Схема определения пола у человека.
Половина сперматозоидов несет X-хромосому, а другая половина — Y-хромосому. Пол ребенка зависит от того, какой сперматозоид оплодотворит яйцеклетку Схема определения пола у человека.
 Генетические карты хромосом — это схема взаимного расположения и относительных расстояний между генами определенных хромосом, находящихся в одной группе сцепления. Впервые в 1913 — 1915 годах на возможность построения генетических карт хромосом указывают Т. Морган и его сотрудники. Они экспериментально показали, что основываясь на явлениях сцепления генов и кроссинговера можно построить генетические карты хромосом. Возможность картирования основана на постоянстве процента кроссинговера между определенными генами. Генетические карты хромосом составлены для многих видов организмов: насекомых (дрозофила, комар, таракан и др. ), грибов (дрожжи, аспергилл), для бактерий и вирусов.
Генетические карты хромосом — это схема взаимного расположения и относительных расстояний между генами определенных хромосом, находящихся в одной группе сцепления. Впервые в 1913 — 1915 годах на возможность построения генетических карт хромосом указывают Т. Морган и его сотрудники. Они экспериментально показали, что основываясь на явлениях сцепления генов и кроссинговера можно построить генетические карты хромосом. Возможность картирования основана на постоянстве процента кроссинговера между определенными генами. Генетические карты хромосом составлены для многих видов организмов: насекомых (дрозофила, комар, таракан и др. ), грибов (дрожжи, аспергилл), для бактерий и вирусов.
 Параллельные линии схематически изображают участок хромосомы; числа указывают относительное расстояние между генами и верхним концом хромосомы, рядом с ними приведены обозначения мутантных аллелей; слева от хромосомы нарисован нормальный орган или нормальная особь, а справа - мутантная. Генетическая карта второй хромосомы дрозофилы
Параллельные линии схематически изображают участок хромосомы; числа указывают относительное расстояние между генами и верхним концом хромосомы, рядом с ними приведены обозначения мутантных аллелей; слева от хромосомы нарисован нормальный орган или нормальная особь, а справа - мутантная. Генетическая карта второй хромосомы дрозофилы
 Как устроен геном у разных организмов? Ген является целостной функциональной единицей наследственности, поскольку любые нарушения его структуры изменяют закодированную информацию или приводят к ее потере. У различных организмов количество генов в геноме может значительно варьировать. Проще всего организован геном вирусов. Он может включать от нескольких до сотен генов. Геном прокариот сложнее, например ДНК кишечной палочки состоит из 4, 6 X 106 пар нуклеотидов, а количество структурных генов – около 4100. Геном эукариот еще более сложен: геном дрозофилы состоит из почти 1, 3 x 108 пар нуклеотидов и насчитывает около 14 000 структурных генов. В геноме человека количество структурных генов составляет 20 000 -25 000. •
Как устроен геном у разных организмов? Ген является целостной функциональной единицей наследственности, поскольку любые нарушения его структуры изменяют закодированную информацию или приводят к ее потере. У различных организмов количество генов в геноме может значительно варьировать. Проще всего организован геном вирусов. Он может включать от нескольких до сотен генов. Геном прокариот сложнее, например ДНК кишечной палочки состоит из 4, 6 X 106 пар нуклеотидов, а количество структурных генов – около 4100. Геном эукариот еще более сложен: геном дрозофилы состоит из почти 1, 3 x 108 пар нуклеотидов и насчитывает около 14 000 структурных генов. В геноме человека количество структурных генов составляет 20 000 -25 000. •
 Взаимодействие генов Различают взаимодействие аллельных и неаллельных генов. Типы взаимодействия аллельных генов: 1. 2. 3. 4. Полное доминирование; Промежуточное наследование (или неполное доминирование); Сверхдоминирование. Кодоминантность. Типы взаимодействия неаллельных генов: 1. 2. 3. 4. Комплиментарность Эпистаз Полимерия Плейотропия
Взаимодействие генов Различают взаимодействие аллельных и неаллельных генов. Типы взаимодействия аллельных генов: 1. 2. 3. 4. Полное доминирование; Промежуточное наследование (или неполное доминирование); Сверхдоминирование. Кодоминантность. Типы взаимодействия неаллельных генов: 1. 2. 3. 4. Комплиментарность Эпистаз Полимерия Плейотропия
 Взаимодействие аллельных генов Полное доминирование — взаимодействие двух аллелей одного гена, когда доминантный аллель полностью исключает проявление действия второго аллеля. В фенотипе присутствует только признак, задаваемый доминантной аллелью. Неполное доминирование — доминантный аллель в гетерозиготном состоянии не полностью подавляет действие рецессивного аллеля. Гетерозиготы имеют промежуточный характер признака. Сверхдоминирование — более сильное проявление признака у гетерозиготной особи, чем у любой гомозиготной. Кодоминирование — проявление у гибридов нового признака, обусловленного взаимодействием двух разных аллелей одного гена. Фенотип гетерозигот не является чемто промежуточным между фенотипами разных гомозигот.
Взаимодействие аллельных генов Полное доминирование — взаимодействие двух аллелей одного гена, когда доминантный аллель полностью исключает проявление действия второго аллеля. В фенотипе присутствует только признак, задаваемый доминантной аллелью. Неполное доминирование — доминантный аллель в гетерозиготном состоянии не полностью подавляет действие рецессивного аллеля. Гетерозиготы имеют промежуточный характер признака. Сверхдоминирование — более сильное проявление признака у гетерозиготной особи, чем у любой гомозиготной. Кодоминирование — проявление у гибридов нового признака, обусловленного взаимодействием двух разных аллелей одного гена. Фенотип гетерозигот не является чемто промежуточным между фенотипами разных гомозигот.
 в гетерозиготном организме(Аа) полностью скрывает присутствие") Полное доминирование это когда доминантный аллельный ген (А) в гетерозиготном организме(Аа) полностью скрывает присутствие рецессивного аллеля гена (а). Как раз явление полного доминирования отражено в 1 законе Менделя – «Законе единообразия гибридов первого поколения» Опыты Менделя по изучению наследования у гороха формы семян(1), цвета семян(2) цвета цветков(3) расположения цветков – пазушные или верхушечные(4), высоты растений(5), цвета бобов(6) и их формы(7) оказались именно примерами взаимодействия аллельных генов с полным доминированием одного аллеля над другим
Полное доминирование это когда доминантный аллельный ген (А) в гетерозиготном организме(Аа) полностью скрывает присутствие рецессивного аллеля гена (а). Как раз явление полного доминирования отражено в 1 законе Менделя – «Законе единообразия гибридов первого поколения» Опыты Менделя по изучению наследования у гороха формы семян(1), цвета семян(2) цвета цветков(3) расположения цветков – пазушные или верхушечные(4), высоты растений(5), цвета бобов(6) и их формы(7) оказались именно примерами взаимодействия аллельных генов с полным доминированием одного аллеля над другим
 Всегда при полном доминировании у генетических гибридов первого поколения, то есть гетерозигот – Аа, полученных от скрещивания генетически чистых гомозиготных родительских линий АА(пурпурные цветки) с аа( белые цветки), полностью исчезали фенотипически растения с белыми цветками. Все гибридные гетерозиготные растения Аа по фенотипу получались как и растения с генотипом АА – то есть с пурпурными цветками.
Всегда при полном доминировании у генетических гибридов первого поколения, то есть гетерозигот – Аа, полученных от скрещивания генетически чистых гомозиготных родительских линий АА(пурпурные цветки) с аа( белые цветки), полностью исчезали фенотипически растения с белыми цветками. Все гибридные гетерозиготные растения Аа по фенотипу получались как и растения с генотипом АА – то есть с пурпурными цветками.
 Неполное доминирование Этим термином обозначается такой тип взаимодействия аллельных генов, когда в первом поколении от скрещивания двух чистых линий АА и аа тоже проявляется как генотипическое, так и фенотипическое единообразие гибридов первого поколения: все Аа. Но все эти гибриды фенотипически не похожи ни на одну родительскую форму, а имеют новое третье промежуточное проявление признака. Например, от скрещивания растений ночной красавицы с красными цветками (ВВ) с растениями с белыми цветками (bb), в первом поколении образуются растения с розовыми цветками(Вb).
Неполное доминирование Этим термином обозначается такой тип взаимодействия аллельных генов, когда в первом поколении от скрещивания двух чистых линий АА и аа тоже проявляется как генотипическое, так и фенотипическое единообразие гибридов первого поколения: все Аа. Но все эти гибриды фенотипически не похожи ни на одну родительскую форму, а имеют новое третье промежуточное проявление признака. Например, от скрещивания растений ночной красавицы с красными цветками (ВВ) с растениями с белыми цветками (bb), в первом поколении образуются растения с розовыми цветками(Вb).
 Неполное доминирование на примере ночной красавицы. В результате образуется новый фенотип.
Неполное доминирование на примере ночной красавицы. В результате образуется новый фенотип.
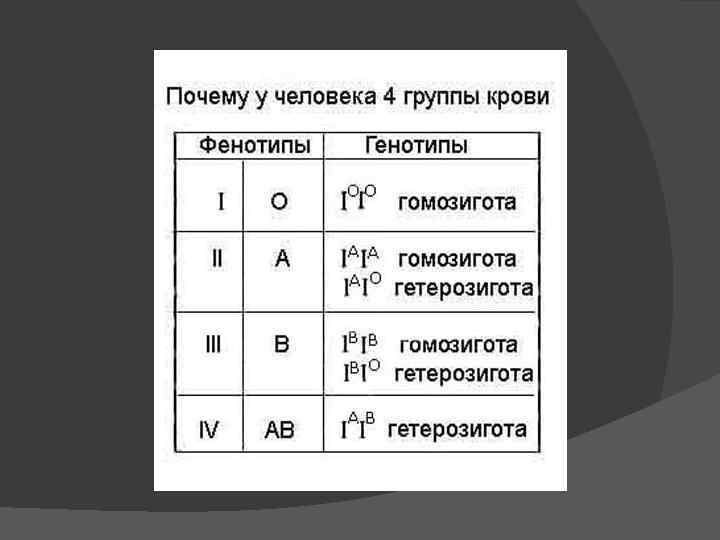
 Кодоминирование Это явление возможно в том случае, когда какойто определенный признак в популяции организмов контролируется не двумя аллельными генами, а тремя. Именно в популяции организмов, так как и в этом случае у любого конкретного организма аллельных генов может быть всегда только два и этих трёх. Чтобы это было понятнее, разберем как наследуются группы крови у человека, которых насчитывается 4 фенотипических и 6 генотипических
Кодоминирование Это явление возможно в том случае, когда какойто определенный признак в популяции организмов контролируется не двумя аллельными генами, а тремя. Именно в популяции организмов, так как и в этом случае у любого конкретного организма аллельных генов может быть всегда только два и этих трёх. Чтобы это было понятнее, разберем как наследуются группы крови у человека, которых насчитывается 4 фенотипических и 6 генотипических
 Группы крови в популяции человека контролируются не двумя аллельными генами, а тремя, названными IA, IO, IB ( аллельные гены, ответственные за формирование группы крови человека обозначаются буквой I с индексами О, А или B) Стало известно, что аллельные гены IA и IB, ответственные за синтез определенных белков агглютиногенов доминируют над третьим рецессивным аллелем IO, контролирующим отсутствие кодирования агглютиногенов. Если в организме человека присутствуют оба рецессивных аллельных гена, то есть если его генотип IOIO, то это будет человек фенотипически с I или, так называемой, нулевой группой крови. А II и III группы крови формируются по правилу полного доминирования: генетически гомозиготный организм IAIA и гетерозиготный IAIO , формируют II фенотипическую группу крови, а гомозиготный IBIB и гетерозиготный IBIO формируют III группу крови. Но при совместном присутствии в генотипе какого-либо человека аллелей IA и IB ни один из них не доминирует над другим. Говорят – они кодоминантны. Что значит фенотипически? Поскольку эти два доминантных аллеля IA IB , отвечая за образование в организме человека двух разных белков агглютиногенов, обеспечат у него проявление нового свойства – IV группу крови.
Группы крови в популяции человека контролируются не двумя аллельными генами, а тремя, названными IA, IO, IB ( аллельные гены, ответственные за формирование группы крови человека обозначаются буквой I с индексами О, А или B) Стало известно, что аллельные гены IA и IB, ответственные за синтез определенных белков агглютиногенов доминируют над третьим рецессивным аллелем IO, контролирующим отсутствие кодирования агглютиногенов. Если в организме человека присутствуют оба рецессивных аллельных гена, то есть если его генотип IOIO, то это будет человек фенотипически с I или, так называемой, нулевой группой крови. А II и III группы крови формируются по правилу полного доминирования: генетически гомозиготный организм IAIA и гетерозиготный IAIO , формируют II фенотипическую группу крови, а гомозиготный IBIB и гетерозиготный IBIO формируют III группу крови. Но при совместном присутствии в генотипе какого-либо человека аллелей IA и IB ни один из них не доминирует над другим. Говорят – они кодоминантны. Что значит фенотипически? Поскольку эти два доминантных аллеля IA IB , отвечая за образование в организме человека двух разных белков агглютиногенов, обеспечат у него проявление нового свойства – IV группу крови.
 Множественный аллелизм Когда за определенный признак отвечает не одна пара аллельных генов, а несколько, то кроме основных( доминантного и рецессивного) генов, могут присутствовать аллельные гены, находящиеся по силе влияния между этими двумя основными генами. Эти аллельные гены обеспечивают промежуточный характер наследования: по отношению к доминантному они ведут себя как рецессивные, а по отношению к рецессивному- как доминантные
Множественный аллелизм Когда за определенный признак отвечает не одна пара аллельных генов, а несколько, то кроме основных( доминантного и рецессивного) генов, могут присутствовать аллельные гены, находящиеся по силе влияния между этими двумя основными генами. Эти аллельные гены обеспечивают промежуточный характер наследования: по отношению к доминантному они ведут себя как рецессивные, а по отношению к рецессивному- как доминантные
 Например, у кроликов сплошная черная окраска меха обусловлена доминантным геном А, гомозиготные рецессивные животные (аа) – белые. Но существуют еще два состояния этого гена, имеющих собственный фенотип в гомозиготном состоянии: шиншилловой аш и гималайской аг окрасок. Шиншилла фенотипически – сплошная серая масть, а гималайский фенотипически – белый, но кончики ушей, хвоста, лап и носа окрашены в черный цвет
Например, у кроликов сплошная черная окраска меха обусловлена доминантным геном А, гомозиготные рецессивные животные (аа) – белые. Но существуют еще два состояния этого гена, имеющих собственный фенотип в гомозиготном состоянии: шиншилловой аш и гималайской аг окрасок. Шиншилла фенотипически – сплошная серая масть, а гималайский фенотипически – белый, но кончики ушей, хвоста, лап и носа окрашены в черный цвет
 Все возможные генотипы данных четырёх фенотипов окрасок представлены в таблице:
Все возможные генотипы данных четырёх фенотипов окрасок представлены в таблице:
 Взаимодействие неаллельных генов Эпистаз - взаимодействие генов, при котором один из них подавляет проявление другого, неаллельного ему, называют эпистазом. Эпистаз противоположен комплементарному взаимодействию. Гены, которые подавляют действие других генов, называются генами-ингибиторами. Такие гены бывают и доминантными, и рецессивными, поэтому различают доминантный и рецессивный эпистаз. Зависимость нескольких признаков от одного гена получила название плейотропности. Это явление было обнаружено еще Менделем, заметившим, что у растений гороха с красными цветками стебли всегда темнее, чем у особей с белыми цветками. Полимерия — обусловленность определенного (обычно количественного) признака несколькими эквивалентными (полимерными) генами. При таком взаимодействии индивидуальное проявление каждого усиливается в результате взаимодействия. Коплиментарность – происходит когда один доминантный ген дополняет действие другого неаллельного ему доминантного гена.
Взаимодействие неаллельных генов Эпистаз - взаимодействие генов, при котором один из них подавляет проявление другого, неаллельного ему, называют эпистазом. Эпистаз противоположен комплементарному взаимодействию. Гены, которые подавляют действие других генов, называются генами-ингибиторами. Такие гены бывают и доминантными, и рецессивными, поэтому различают доминантный и рецессивный эпистаз. Зависимость нескольких признаков от одного гена получила название плейотропности. Это явление было обнаружено еще Менделем, заметившим, что у растений гороха с красными цветками стебли всегда темнее, чем у особей с белыми цветками. Полимерия — обусловленность определенного (обычно количественного) признака несколькими эквивалентными (полимерными) генами. При таком взаимодействии индивидуальное проявление каждого усиливается в результате взаимодействия. Коплиментарность – происходит когда один доминантный ген дополняет действие другого неаллельного ему доминантного гена.
 Комплиментарность Происходит когда один доминантный ген дополняет действие другого неаллельного ему доминантного гена. В результате происходит образование нового проявления признака по типу взаимопомощи, взаимодополнения друга. Образование не нового признака, а лишь образование нового проявления того же самого признака.
Комплиментарность Происходит когда один доминантный ген дополняет действие другого неаллельного ему доминантного гена. В результате происходит образование нового проявления признака по типу взаимопомощи, взаимодополнения друга. Образование не нового признака, а лишь образование нового проявления того же самого признака.
 Комплиментарность прослеживается у цветков душистого горошка. При скрещивании двух различных линий этого растения с белыми цветками у гибридов F 1 цветки оказались пурпурными. При скрещивании растений F 1 друг с другом ( самоопылении) в F 2 наблюдается расщепление признаков окраски цветков растений в отношении близком к 9: 7. Пурпурные цветки были обнаружены у 9/16 растений, белые у 7/16. Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментным белком антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника антоциана – пропигмента. Ген В определяет синтез ферментного белка, под действием которого из пропигмента образуется антоциан
Комплиментарность прослеживается у цветков душистого горошка. При скрещивании двух различных линий этого растения с белыми цветками у гибридов F 1 цветки оказались пурпурными. При скрещивании растений F 1 друг с другом ( самоопылении) в F 2 наблюдается расщепление признаков окраски цветков растений в отношении близком к 9: 7. Пурпурные цветки были обнаружены у 9/16 растений, белые у 7/16. Объяснение такого результата состоит в том, что каждый из доминантных генов не может вызвать появление окраски, определяемой пигментным белком антоцианом. У душистого горошка есть ген А, обусловливающий синтез бесцветного предшественника антоциана – пропигмента. Ген В определяет синтез ферментного белка, под действием которого из пропигмента образуется антоциан
 Эпистаз Сущность эпистаза сводится к подавлению проявления генов одной аллельной пары генами другой пары. Если, например, ген Ш подавляет действие гена С, то такой тип подавления носит название – доминантный эпистаз. Ген, который подавляет действие другого гена, называется ингибидор ( I-ингибидор). Если гены ii подавляют действие гена С, то это пример рецессивного эпистаза.
Эпистаз Сущность эпистаза сводится к подавлению проявления генов одной аллельной пары генами другой пары. Если, например, ген Ш подавляет действие гена С, то такой тип подавления носит название – доминантный эпистаз. Ген, который подавляет действие другого гена, называется ингибидор ( I-ингибидор). Если гены ii подавляют действие гена С, то это пример рецессивного эпистаза.
 Из таблицы видно, что формулы 3 и 4 одинаковые по фенотипу, но разные по генотипу. В строке 4 записано наследование генотипов при рецессивном эпистазе, а в строках 1, 2 и 3 – при доминативном.
Из таблицы видно, что формулы 3 и 4 одинаковые по фенотипу, но разные по генотипу. В строке 4 записано наследование генотипов при рецессивном эпистазе, а в строках 1, 2 и 3 – при доминативном.
 Полимерия Явление, когда несколько разных доминантных генов отвечают за проявление одного и того же признака называют полимерией Интенсивность проявления признака зависит от количества доминантных генов в генотипе. Обычно этими генами определяются количественные признаки ( интенсивность роста, яйценоскость, жирность и количество молока и т. п. ). Но могут определяться и качественные признаки. Полимерные гены обозначают одной буквой, но с различной нумерацией, например, А 1, А 2, А 3 и т. д.
Полимерия Явление, когда несколько разных доминантных генов отвечают за проявление одного и того же признака называют полимерией Интенсивность проявления признака зависит от количества доминантных генов в генотипе. Обычно этими генами определяются количественные признаки ( интенсивность роста, яйценоскость, жирность и количество молока и т. п. ). Но могут определяться и качественные признаки. Полимерные гены обозначают одной буквой, но с различной нумерацией, например, А 1, А 2, А 3 и т. д.
 В F 1 по первому закону Менделя наблюдается единообразие гибридов первого поколения – все потомки получились красными , как доминантная родительская форма. За интенсивность красной окраски отвечают оба разных гена А 1 и А 2 , а присутствие в генотипе потомства F 1 только по одному их аллелю обеднило интенсивность красной окраски. Вторые пары алелей в F 1 рецессивные а 1 и а 2 , поэтому они не несут признака красной окраски В F 2 от скрещивания красной пшеницы А 1 а 1 А 2 а 2 с такой же красной А 1 а 1 А 2 а 2 обнаруживаются и темно-красные и темноватокрасные( с разными генотипами) и светлокрасные и чисто белые.
В F 1 по первому закону Менделя наблюдается единообразие гибридов первого поколения – все потомки получились красными , как доминантная родительская форма. За интенсивность красной окраски отвечают оба разных гена А 1 и А 2 , а присутствие в генотипе потомства F 1 только по одному их аллелю обеднило интенсивность красной окраски. Вторые пары алелей в F 1 рецессивные а 1 и а 2 , поэтому они не несут признака красной окраски В F 2 от скрещивания красной пшеницы А 1 а 1 А 2 а 2 с такой же красной А 1 а 1 А 2 а 2 обнаруживаются и темно-красные и темноватокрасные( с разными генотипами) и светлокрасные и чисто белые.
 Плеотропия Это зависимость нескольких признаков от одного гена. Сам механизм множественного действия гена заключается в том, что белок, кодируемый этим геном, может в той или иной степени участвовать в различных процессах. Большинство генов имеют плейотропное действие. В плейотропии заключается соотносительный характер наследования признаков организмом.
Плеотропия Это зависимость нескольких признаков от одного гена. Сам механизм множественного действия гена заключается в том, что белок, кодируемый этим геном, может в той или иной степени участвовать в различных процессах. Большинство генов имеют плейотропное действие. В плейотропии заключается соотносительный характер наследования признаков организмом.
 Над проектом работал Ученик 11 -А класса Одесской ООШ № 100 Сивков Никита Тимурович
Над проектом работал Ученик 11 -А класса Одесской ООШ № 100 Сивков Никита Тимурович