

5. Ядро-2014.ppt
- Количество слайдов: 60
 Ядро. Организация генетического материала у эукариот
Ядро. Организация генетического материала у эукариот
 Научно-исследовательская работа студента • Индивидуальный портфолио • Индивидуальный проект • До 15 октября – выбрать тему • После 1 ноября - представление работ на практических занятиях
Научно-исследовательская работа студента • Индивидуальный портфолио • Индивидуальный проект • До 15 октября – выбрать тему • После 1 ноября - представление работ на практических занятиях
 Индивидуальный портфолио 1. Личные данные 2. Тема работы, научный руководитель 3. План работы, сроки выполнения 4. Разделы портфолио а) Список изученных научных статей б) Обзоры статей (не менее 3 -х) в) Выводы г) Рекомендации по практическому использованию в медицине 5. Отзывы коллег
Индивидуальный портфолио 1. Личные данные 2. Тема работы, научный руководитель 3. План работы, сроки выполнения 4. Разделы портфолио а) Список изученных научных статей б) Обзоры статей (не менее 3 -х) в) Выводы г) Рекомендации по практическому использованию в медицине 5. Отзывы коллег
 Индивидуальный проект 1. Тема работы, научный руководитель 2. План работы, сроки выполнения 3. Разделы проекта (представление в виде Power Point) – 10 мин а) Тема проекта б) Цель проекта в) Результаты исследования г) Выводы д) Рекомендации по практическому использованию в медицине е) Список литературы 4. Отзывы коллег
Индивидуальный проект 1. Тема работы, научный руководитель 2. План работы, сроки выполнения 3. Разделы проекта (представление в виде Power Point) – 10 мин а) Тема проекта б) Цель проекта в) Результаты исследования г) Выводы д) Рекомендации по практическому использованию в медицине е) Список литературы 4. Отзывы коллег
 Организация генетического материала у эукариот
Организация генетического материала у эукариот
 Эукариотная ДНК Расположена: • В ядре • В митохондриях
Эукариотная ДНК Расположена: • В ядре • В митохондриях
 Ядерная ДНК: • Длинные линейные молекулы; • В виде хроматина ↔ хромосом; • Около 10% ДНК – кодирующая, остальная – некодирующая; • Активность ядерных генов зависит от: • Онтогенетического периода; • Типа клетки !!! Необходим механизм сохранения ГИ для оптимальной пространственной и временной активности
Ядерная ДНК: • Длинные линейные молекулы; • В виде хроматина ↔ хромосом; • Около 10% ДНК – кодирующая, остальная – некодирующая; • Активность ядерных генов зависит от: • Онтогенетического периода; • Типа клетки !!! Необходим механизм сохранения ГИ для оптимальной пространственной и временной активности
 Ядро • Содержит ~ 98% клеточной ДНК; • Обеспечивает функциональную компактизацию ДНК: • Эухроматин – активная ГИ; • Гетерохроматин – неактивная ДНК. • Контроль жизнедеятельности клетки путем дифференциальной экспрессии генов: • Транскрипция и процессинг РНК; • Контроль клеточного деления и передачи ГИ от клетки – к клетке: • Репликация ДНК и митоз • Биогенез рибосом
Ядро • Содержит ~ 98% клеточной ДНК; • Обеспечивает функциональную компактизацию ДНК: • Эухроматин – активная ГИ; • Гетерохроматин – неактивная ДНК. • Контроль жизнедеятельности клетки путем дифференциальной экспрессии генов: • Транскрипция и процессинг РНК; • Контроль клеточного деления и передачи ГИ от клетки – к клетке: • Репликация ДНК и митоз • Биогенез рибосом
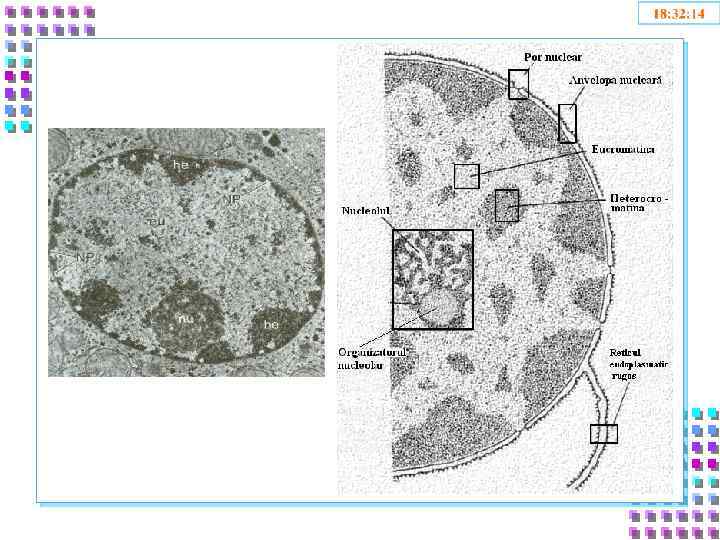
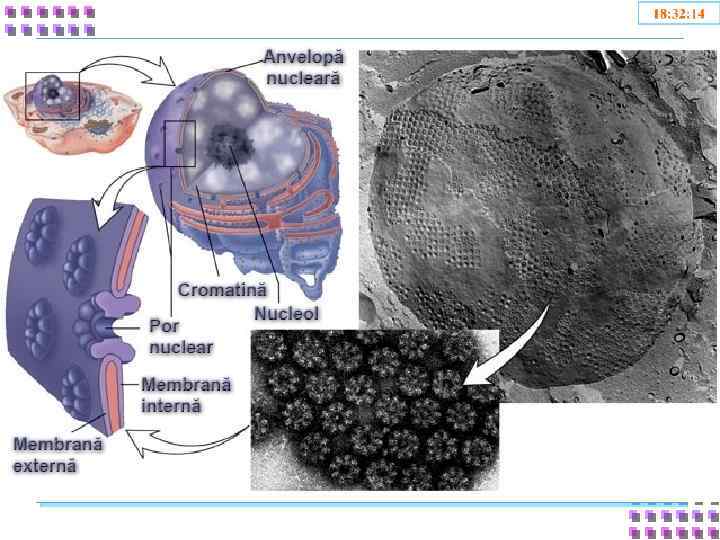
 Ядро • Ядерная оболочка: • • • Внешняя ядерная мембрана Внутренняя ядерная мембрана Перинуклеарное пространство Ядерная ламина Поровый комплекс • Кариоплазма • Лабильная фаза • Ядерный скелет • Хроматин • Эухроматин • Гетерохроматин • Ядрышки
Ядро • Ядерная оболочка: • • • Внешняя ядерная мембрана Внутренняя ядерная мембрана Перинуклеарное пространство Ядерная ламина Поровый комплекс • Кариоплазма • Лабильная фаза • Ядерный скелет • Хроматин • Эухроматин • Гетерохроматин • Ядрышки
 Ядро в разных этапах клеточного цикла
Ядро в разных этапах клеточного цикла
 -Белки ядерного матрикса: матрины HMG (High Mobility Group) SAP (Scaffold") -Нуклеопорины -Транспортины (импортины, экспортины) -Белки ядерного матрикса: матрины HMG (High Mobility Group) SAP (Scaffold Associated Proteins) ламины A, B, C нуклеоплазмины -Нуклеолины -Гистоны (H 1, H 2 A, H 2 B, H 3, H 4) -Негистоны : ДНК-полимераза РНК-полимераза топоизомеразы лигазы геликаза праймаза …
-Нуклеопорины -Транспортины (импортины, экспортины) -Белки ядерного матрикса: матрины HMG (High Mobility Group) SAP (Scaffold Associated Proteins) ламины A, B, C нуклеоплазмины -Нуклеолины -Гистоны (H 1, H 2 A, H 2 B, H 3, H 4) -Негистоны : ДНК-полимераза РНК-полимераза топоизомеразы лигазы геликаза праймаза …
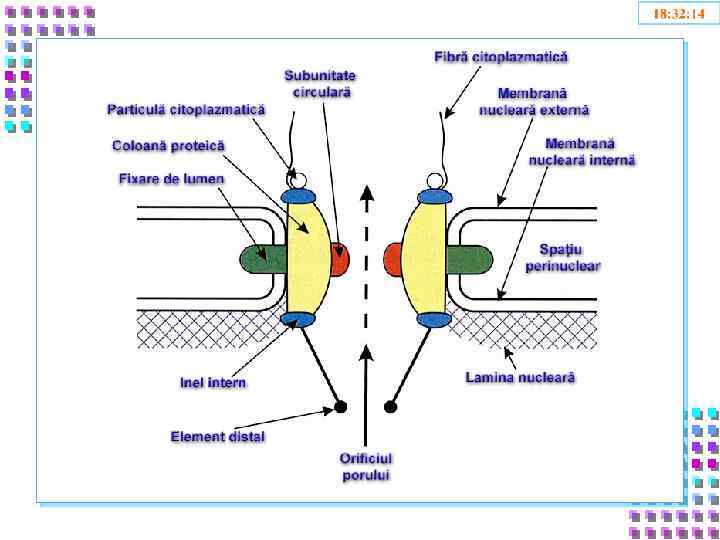
 - экспортины") Белки ядерной мембраны • Нуклеопорины • Транспортины - импортины (цитоплазма → ядро) - экспортины ( ядро → цитоплазма)
Белки ядерной мембраны • Нуклеопорины • Транспортины - импортины (цитоплазма → ядро) - экспортины ( ядро → цитоплазма)
 Ядерная ламина • Поддерживает целостность и форму ядра, • Отвечает за дезинтеграцию и реорганизацию ядерной оболочки в процессе митоза, • Обеспечивает фиксацию хроматиновых нитей в интерфазе, • Обеспечивает работу порового комплекса
Ядерная ламина • Поддерживает целостность и форму ядра, • Отвечает за дезинтеграцию и реорганизацию ядерной оболочки в процессе митоза, • Обеспечивает фиксацию хроматиновых нитей в интерфазе, • Обеспечивает работу порового комплекса
 ØНа молекулярном уровне – синтез белка Lamin A Øна клеточном уровне – организация ядерной ламины ØНа уровне организма –участие в контроле процессов старения
ØНа молекулярном уровне – синтез белка Lamin A Øна клеточном уровне – организация ядерной ламины ØНа уровне организма –участие в контроле процессов старения
 Механизм синтеза ламинов
Механизм синтеза ламинов
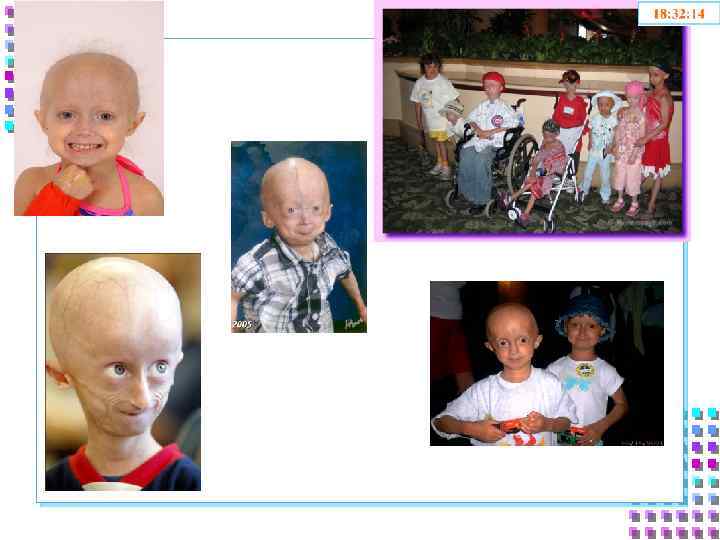
 Сравнительная форма ядер
Сравнительная форма ядер
 • Собственно матрикс • Белки") Ядерный матрикс • Лабильная фракция (вода, ионы, растворимые белки) • Собственно матрикс • Белки ядерного матрикса: - матрины - негистоны (ферменты, HMG) - нуклеоплазмины - SAP
Ядерный матрикс • Лабильная фракция (вода, ионы, растворимые белки) • Собственно матрикс • Белки ядерного матрикса: - матрины - негистоны (ферменты, HMG) - нуклеоплазмины - SAP
 Функции ядерного матрикса • Обеспечивает форму и внутреннюю организацию ядра • Участвует в процессах упаковки хроматина • Обеспечивает процессы репликации и транскрипции
Функции ядерного матрикса • Обеспечивает форму и внутреннюю организацию ядра • Участвует в процессах упаковки хроматина • Обеспечивает процессы репликации и транскрипции
 профаза ХРОМАТИН телофаза ХРОМОСОМЫ • Хроматин – это частично упакованный генетический материал в интерфазе; • Хромосома – максимально упакованный генетический материал в митозе. • Химический состав: • 30% ДНК + 40% гистоны + 25% негистоновые белки + 5% РНК
профаза ХРОМАТИН телофаза ХРОМОСОМЫ • Хроматин – это частично упакованный генетический материал в интерфазе; • Хромосома – максимально упакованный генетический материал в митозе. • Химический состав: • 30% ДНК + 40% гистоны + 25% негистоновые белки + 5% РНК
 Хроматин = хромосома Хромосомная ДНК – содержит, передает и реализует ГИ; • Длинные, линейные молекулы; • Гетерогенные молекулы: • Кодирующие последовательности (гены): • Активные – транскрибируемые; • Неактивные – нетранскрибируемые. • Некодирующие последовательности: • Структурные – теломеры, центромеры, сателлиты; • Регуляторные
Хроматин = хромосома Хромосомная ДНК – содержит, передает и реализует ГИ; • Длинные, линейные молекулы; • Гетерогенные молекулы: • Кодирующие последовательности (гены): • Активные – транскрибируемые; • Неактивные – нетранскрибируемые. • Некодирующие последовательности: • Структурные – теломеры, центромеры, сателлиты; • Регуляторные
 Хроматин = хромосома Гистоновые белки: • Глобулярные • Богатые Liz и Arg; • 5 классов: H 1, H 2 A, H 2 B, H 3, H 4 • Функция: • Упаковка ДНК на нуклеосомном уровне • Неспецифичный контроль экспрессии генов Тип гистонов Характерные аминокислоты К-во аминокислот Конформация белка H 1 Liz 215 NH 2 COOH H 2 A Leu, Liz 129 NH 2 COOH H 2 B Ser, Pro, Liz 125 NH 2 COOH H 3 Arg, Cis 135 NH 2 H 4 Arg, Liz 102 NH 2 COOH
Хроматин = хромосома Гистоновые белки: • Глобулярные • Богатые Liz и Arg; • 5 классов: H 1, H 2 A, H 2 B, H 3, H 4 • Функция: • Упаковка ДНК на нуклеосомном уровне • Неспецифичный контроль экспрессии генов Тип гистонов Характерные аминокислоты К-во аминокислот Конформация белка H 1 Liz 215 NH 2 COOH H 2 A Leu, Liz 129 NH 2 COOH H 2 B Ser, Pro, Liz 125 NH 2 COOH H 3 Arg, Cis 135 NH 2 H 4 Arg, Liz 102 NH 2 COOH
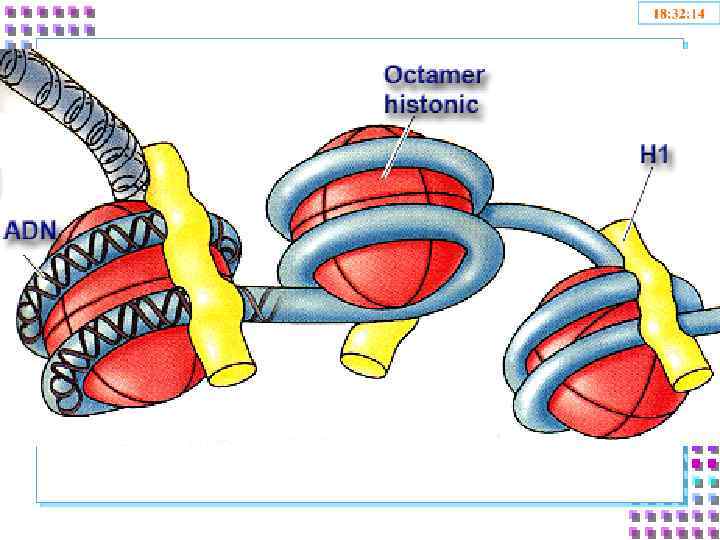
 Гистоновые белки ДНК + гистоны = нуклеосома a. Нуклеосомное ядро: 2 H 2 A Гистоновый октамер 2 H 2 B 2 H 3 2 H 4 b. 200 п. о. нуклеосомная ДНК c. H 1 – стабилизирует DNP
Гистоновые белки ДНК + гистоны = нуклеосома a. Нуклеосомное ядро: 2 H 2 A Гистоновый октамер 2 H 2 B 2 H 3 2 H 4 b. 200 п. о. нуклеосомная ДНК c. H 1 – стабилизирует DNP

 Хромосомная РНК • Первичные транскрипты • микро. РНК в составе ядерных ферментов
Хромосомная РНК • Первичные транскрипты • микро. РНК в составе ядерных ферментов
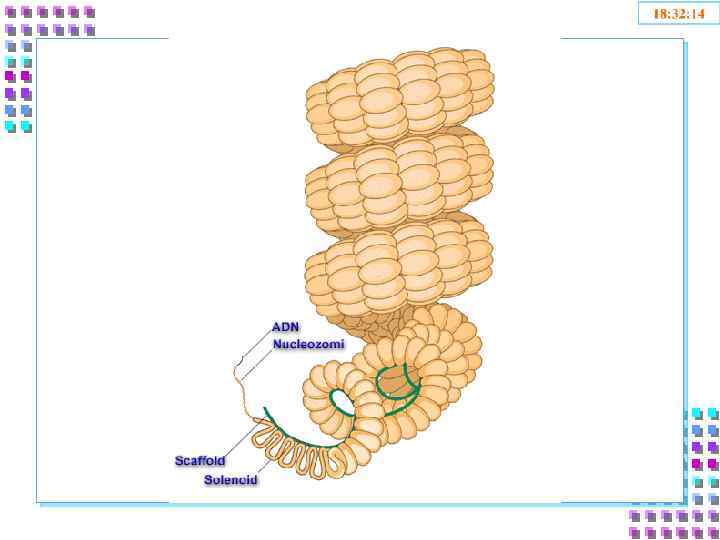
 Уровни упаковки ДНК • I уровень – нуклеосомный = нити хроматина; • II уровень – соленоид = нити хроматина; • III уровень – петли • IV уровень – метафазная хромосома
Уровни упаковки ДНК • I уровень – нуклеосомный = нити хроматина; • II уровень – соленоид = нити хроматина; • III уровень – петли • IV уровень – метафазная хромосома
 Уровни упаковки ГИ отличаются: - Степенью конденспации ДНК; - Толщиной хроматиновой нити; - Механизмом образования; - Уровнем транскрипционной активности; - Периодом клеточного цикла.
Уровни упаковки ГИ отличаются: - Степенью конденспации ДНК; - Толщиной хроматиновой нити; - Механизмом образования; - Уровнем транскрипционной активности; - Периодом клеточного цикла.
 I уровень- нити хроматина: • Полинуклеосомная нить 11 nm; • Степень конденсации – x 6 раз; • Механизм образования: • Взаимодействие ДНК- с гистоновыми основными белками+ • Транскрипция - возможна.
I уровень- нити хроматина: • Полинуклеосомная нить 11 nm; • Степень конденсации – x 6 раз; • Механизм образования: • Взаимодействие ДНК- с гистоновыми основными белками+ • Транскрипция - возможна.
 I уровень- нити хроматина 10 nm
I уровень- нити хроматина 10 nm
 – активная экспрессия гена • Метилирование") Изменение гистонов: • Метилирование H 3 (Lys 4) – активная экспрессия гена • Метилирование H 3 (Lys 9) – атенуация транскрипции • Ацетилирование гистонов – расплетание хроматина, транскрипция • Дезацетилирование гистонов – упаковка хроматина, инактивация транскрипции • Фосфорилирование H 1 – сверхконденсация ДНК • Дефосфорилирование H 1 – деконденсация хроматина
Изменение гистонов: • Метилирование H 3 (Lys 4) – активная экспрессия гена • Метилирование H 3 (Lys 9) – атенуация транскрипции • Ацетилирование гистонов – расплетание хроматина, транскрипция • Дезацетилирование гистонов – упаковка хроматина, инактивация транскрипции • Фосфорилирование H 1 – сверхконденсация ДНК • Дефосфорилирование H 1 – деконденсация хроматина
 Пора переходить от слов к делу… • Как повлиять на ацетилирование гистонов для защиты от стресса и старения, для борьбы с онкологическими заболеваниями? • Каков точный механизм участия фосфорилирования гистонов в возникновении онкологических заболевания и старении? • Как можно повлиять на механизмы модификации гистонов с целью достижения терапевтического эффекта при лечении заболеваний, связанных с возрастом?
Пора переходить от слов к делу… • Как повлиять на ацетилирование гистонов для защиты от стресса и старения, для борьбы с онкологическими заболеваниями? • Каков точный механизм участия фосфорилирования гистонов в возникновении онкологических заболевания и старении? • Как можно повлиять на механизмы модификации гистонов с целью достижения терапевтического эффекта при лечении заболеваний, связанных с возрастом?
 II уровень – соленоид • • • Нить хроматина 30 nm; Степень конденсации – x 40 раз; Механизм образования: • Фосфорилирование H 1 сверхспирализация полинуклеосомной нити – по 6 нуклеосом в витке; • Блокирование транскрипции ДНК.
II уровень – соленоид • • • Нить хроматина 30 nm; Степень конденсации – x 40 раз; Механизм образования: • Фосфорилирование H 1 сверхспирализация полинуклеосомной нити – по 6 нуклеосом в витке; • Блокирование транскрипции ДНК.
 Соленоид
Соленоид
 Нить хроматина Соленоид– 30 nm Нуклеосомная нить – 10 nm
Нить хроматина Соленоид– 30 nm Нуклеосомная нить – 10 nm
 III уровень- петли • Петли большие и малые, диффузные или компактные, толщина – 300 nm; • Степень конденсации – x 600 -1000; • Механизм образования: • Прикрепление нити хроматина 30 nm к белковой оси – scaffold; • Взаимодействие SAR – scaffold • Транскрипция генов – заблокированна
III уровень- петли • Петли большие и малые, диффузные или компактные, толщина – 300 nm; • Степень конденсации – x 600 -1000; • Механизм образования: • Прикрепление нити хроматина 30 nm к белковой оси – scaffold; • Взаимодействие SAR – scaffold • Транскрипция генов – заблокированна
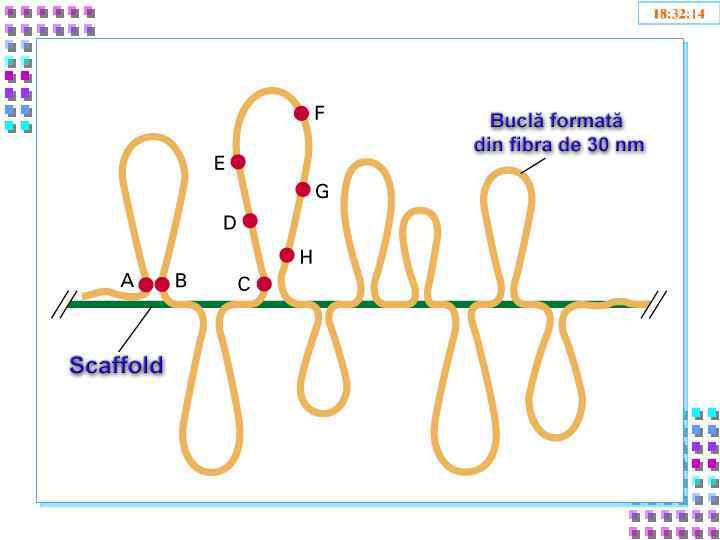
 - (Scaffold /Matrix Associated Region) – специфические сайты ДНК ответственные за") !!! SAR (MAR) - (Scaffold /Matrix Associated Region) – специфические сайты ДНК ответственные за присоединение к scaffold – хромосомной оси; !!! SAP- (Scaffold /Matrix Associated Proteins) - сайт специфические белки взаимодействуют с: - нитями хроматина; - обеспечивают III уровень упаковки - ламиной lamina fibrosa с поверхности внутренней мембраны ядерной оболочки.
!!! SAR (MAR) - (Scaffold /Matrix Associated Region) – специфические сайты ДНК ответственные за присоединение к scaffold – хромосомной оси; !!! SAP- (Scaffold /Matrix Associated Proteins) - сайт специфические белки взаимодействуют с: - нитями хроматина; - обеспечивают III уровень упаковки - ламиной lamina fibrosa с поверхности внутренней мембраны ядерной оболочки.
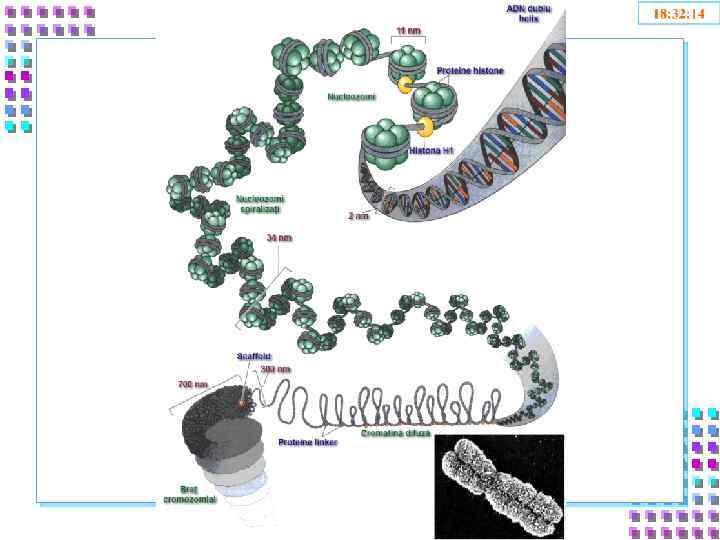
 IV уровень – метафазная хромосома • • • Наивысший уровень упаковки ДНК – x 10000 раз; Толщина хроматиды ~700 nm; Механизм образования: • Фосфорилирование ламины --- диссоциация ядерной оболочки; • Полное фосфорилирование H 1 – конденсация хроматина. • Транскрипция заблокирована
IV уровень – метафазная хромосома • • • Наивысший уровень упаковки ДНК – x 10000 раз; Толщина хроматиды ~700 nm; Механизм образования: • Фосфорилирование ламины --- диссоциация ядерной оболочки; • Полное фосфорилирование H 1 – конденсация хроматина. • Транскрипция заблокирована
 2 nm 11 nm 300 nm 700 nm 1400 nm
2 nm 11 nm 300 nm 700 nm 1400 nm
 Период клеточного цикла и степень конденсации хроматина • Интерфаза: • • • Период G 1; Период S; Период G 2; I уровень III уровень • Митоз: • • Профаза Метафаза Анафаза Телофаза IV уровень
Период клеточного цикла и степень конденсации хроматина • Интерфаза: • • • Период G 1; Период S; Период G 2; I уровень III уровень • Митоз: • • Профаза Метафаза Анафаза Телофаза IV уровень
 Функциональная классификация хроматина: • Эухроматин: - Активные фрагменты ДНК; - Содержит кодогенную ДНК. • Гетерохроматин: • • • Неактивные фрагменты ДНК; Содержит некодогенную ДНК; Содержит кодогенную ДНК, но неактивную в данный момент; • Может быть конститутивным и факультативным.
Функциональная классификация хроматина: • Эухроматин: - Активные фрагменты ДНК; - Содержит кодогенную ДНК. • Гетерохроматин: • • • Неактивные фрагменты ДНК; Содержит некодогенную ДНК; Содержит кодогенную ДНК, но неактивную в данный момент; • Может быть конститутивным и факультативным.
 Гетерохроматин • Конститутивный: • Некодирующая, репетитивная ДНК; • Ценромеры, теломеры, сателлиты, спейсеры. • Факультативный: • • Кодирующая, неактивная ДНК; Может переходить в эухроматин; Может быть аутосомным или половым; Определяет: • Клеточную дифференциацию; • Половую дифференциацию; • Контроль онтогенеза
Гетерохроматин • Конститутивный: • Некодирующая, репетитивная ДНК; • Ценромеры, теломеры, сателлиты, спейсеры. • Факультативный: • • Кодирующая, неактивная ДНК; Может переходить в эухроматин; Может быть аутосомным или половым; Определяет: • Клеточную дифференциацию; • Половую дифференциацию; • Контроль онтогенеза
 • Центромера = первичная перетяжка • Теломера
• Центромера = первичная перетяжка • Теломера
 Центромера • Повторяющиеся последовательности, богатые A/T • Конститутивный гетерохроматин • H 3 замещен CENP-A • Связывает хроматиды до анафазы митоза (коэзин) • Обеспечивает присоединение кинетохоров • Обеспечивает взаимодействие с веретеном деления
Центромера • Повторяющиеся последовательности, богатые A/T • Конститутивный гетерохроматин • H 3 замещен CENP-A • Связывает хроматиды до анафазы митоза (коэзин) • Обеспечивает присоединение кинетохоров • Обеспечивает взаимодействие с веретеном деления
 Теломера
Теломера
 длиной 3 -20 kb") Теломера • Последовательности богатая G/C (у H. sapiens - TTAGGG) длиной 3 -20 kb • Образует петли на концах хромосом (TFR 1, TFR 2) • Предотвращает укорачивание хромосом как следствие репликации • Предотвращает соединение хромосом (индивидуализация хромосом) • Обеспечивает защиту хромосом
Теломера • Последовательности богатая G/C (у H. sapiens - TTAGGG) длиной 3 -20 kb • Образует петли на концах хромосом (TFR 1, TFR 2) • Предотвращает укорачивание хромосом как следствие репликации • Предотвращает соединение хромосом (индивидуализация хромосом) • Обеспечивает защиту хромосом
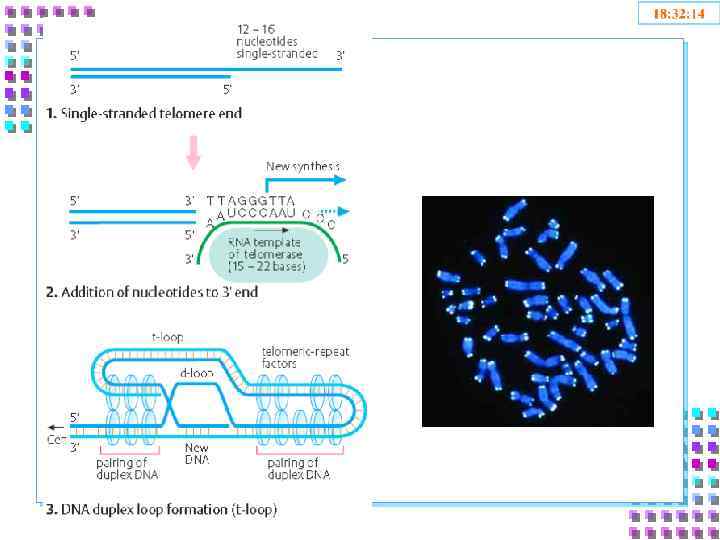
 Функции теломеров • Предотвращают слипание хромосом • Обеспечивают индивидуальность и целостность хромосомы, ее длину • Защищают концы хромосом от экзонуклеаз • Обеспечивают коньюгацию и правильную рекомбинацию хромосом
Функции теломеров • Предотвращают слипание хромосом • Обеспечивают индивидуальность и целостность хромосомы, ее длину • Защищают концы хромосом от экзонуклеаз • Обеспечивают коньюгацию и правильную рекомбинацию хромосом
 The Nobel Prize in Medicine 2009 • "for the discovery of how chromosomes are protected by telomeres and the enzyme telomerase" • Elizabeth H. Blackburn, University of California San Francisco, CA, USA • Carol W. Greider, Johns Hopkins University School of Medicine , Baltimore, MD, USA • Jack W. Szostak, Harvard Medical School; Massachusetts General Hospital , Boston, MA, USA; Howard Hughes Medical Institute
The Nobel Prize in Medicine 2009 • "for the discovery of how chromosomes are protected by telomeres and the enzyme telomerase" • Elizabeth H. Blackburn, University of California San Francisco, CA, USA • Carol W. Greider, Johns Hopkins University School of Medicine , Baltimore, MD, USA • Jack W. Szostak, Harvard Medical School; Massachusetts General Hospital , Boston, MA, USA; Howard Hughes Medical Institute
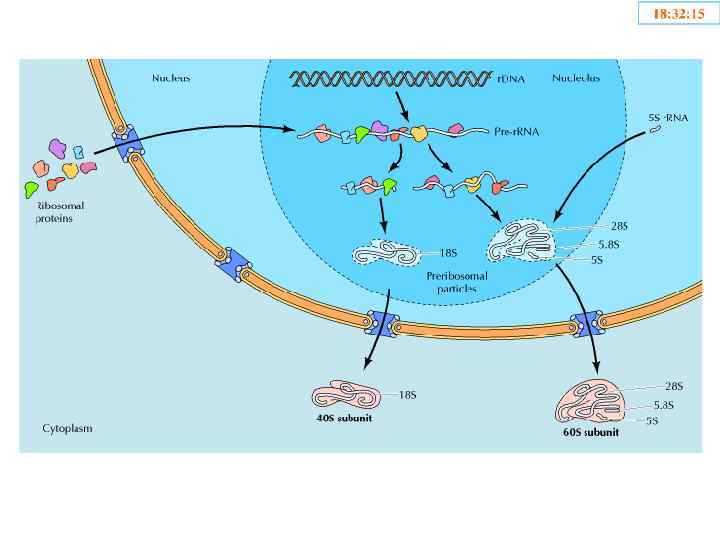
 Ядрышко • Ядрышковый организатор – фрагменты ДНК содержащие информацию о р. РНК; • р. РНК: первичные транскрипты 45 S; р. РНК 5 S, р. РНК 5, 8 S, р. РНК 18 S, р. РНК 28 S; • Рибосомные белки; • Белки аппарата синтеза р. РНК; • RNP 40 S и 60 S.
Ядрышко • Ядрышковый организатор – фрагменты ДНК содержащие информацию о р. РНК; • р. РНК: первичные транскрипты 45 S; р. РНК 5 S, р. РНК 5, 8 S, р. РНК 18 S, р. РНК 28 S; • Рибосомные белки; • Белки аппарата синтеза р. РНК; • RNP 40 S и 60 S.
 The Nobel Prize in Chemistry 2009 • "for studies of the structure and function of the ribosome“ • Venkatraman Ramakrishnan, MRC Laboratory of Molecular Biology , Cambridge, United Kingdom • Thomas A. Steitz, Yale University New Haven, CT, USA; Howard Hughes Medical Institute • Ada E. Yonath, Weizmann Institute of Science Rehovot, Israel 59
The Nobel Prize in Chemistry 2009 • "for studies of the structure and function of the ribosome“ • Venkatraman Ramakrishnan, MRC Laboratory of Molecular Biology , Cambridge, United Kingdom • Thomas A. Steitz, Yale University New Haven, CT, USA; Howard Hughes Medical Institute • Ada E. Yonath, Weizmann Institute of Science Rehovot, Israel 59
 Следующая тема: Строение и функции генов
Следующая тема: Строение и функции генов