

ядро и митоз лекция 6 2012.ppt
- Количество слайдов: 77
 Ядро/а
Ядро/а
 вешенки и спорах (HCl Гимза-Романовского) шампиньона 6 Nectria heematococca DIC") Ядра в мицелии (DAPI) вешенки и спорах (HCl Гимза-Романовского) шампиньона 6 Nectria heematococca DIC Inoue et al. , 1998
Ядра в мицелии (DAPI) вешенки и спорах (HCl Гимза-Романовского) шампиньона 6 Nectria heematococca DIC Inoue et al. , 1998
 Функции ядра Ядро - имеет две главные функции: хранение наследственной информации и координирование всех клеточных активностей (метаболизм, рост, со всеми синтетическими процессами от которых он зависит и клеточное деление)
Функции ядра Ядро - имеет две главные функции: хранение наследственной информации и координирование всех клеточных активностей (метаболизм, рост, со всеми синтетическими процессами от которых он зависит и клеточное деление)
 Число ядер • У грибов встречаются одноядерные и многоядерные вегетативные клетки. Споры также могут быть одно-, двух-, четырехядерные и более в зависимости от систематической принадлежности гриба (у A. bisporus до 8 ядер). Грибы, имеющие неклеточный мицелий (Zygomycetes), содержат большое количество ядер. Хитридиевые большей частью одноядерны. У сумчатых и базидиальных грибов число ядер варьирует от 1 -2 до 20 -30 и более в зависимости от систематического положения гриба, типа клеток (вегетативной или репродуктивной структуры) и ее возраста.
Число ядер • У грибов встречаются одноядерные и многоядерные вегетативные клетки. Споры также могут быть одно-, двух-, четырехядерные и более в зависимости от систематической принадлежности гриба (у A. bisporus до 8 ядер). Грибы, имеющие неклеточный мицелий (Zygomycetes), содержат большое количество ядер. Хитридиевые большей частью одноядерны. У сумчатых и базидиальных грибов число ядер варьирует от 1 -2 до 20 -30 и более в зависимости от систематического положения гриба, типа клеток (вегетативной или репродуктивной структуры) и ее возраста.
 Таксон размеры") Количество ядер в клетках ножки плодового тела у некоторых базидиомицетов (Chiu, 1996) Таксон размеры Agaricus bisporus Armillaria mellea 4 -32 1 Coprinus cinereus Flammulina velutipes Lentinula edodes Pholiota nameko Volvariella bombycina 2 -156 2 -32 2 -8 2 -4 5 -30
Количество ядер в клетках ножки плодового тела у некоторых базидиомицетов (Chiu, 1996) Таксон размеры Agaricus bisporus Armillaria mellea 4 -32 1 Coprinus cinereus Flammulina velutipes Lentinula edodes Pholiota nameko Volvariella bombycina 2 -156 2 -32 2 -8 2 -4 5 -30
 Количество ядер у Agaricus bisporus • Клетки вегетативного мицелия содержат от 3 до 20 ядер • Клетки шляпки плодового тела имеют в среднем 2 -6 ядер • Клетки ножки – до 32 ядер • Базидиоспоры – 2 -8 ядер
Количество ядер у Agaricus bisporus • Клетки вегетативного мицелия содержат от 3 до 20 ядер • Клетки шляпки плодового тела имеют в среднем 2 -6 ядер • Клетки ножки – до 32 ядер • Базидиоспоры – 2 -8 ядер
. Диплоидные") Размер ядер Гаплоидные ядра у большинства грибов - очень мелкие (1 -3 мкм). Диплоидные в 2 -2. 5 раза больше. Диплоидное ядро Verpa bogemica достигает 15 мкм. Наиболее крупные ядра встречаются в органах, связанных с половым воспроизведением, например, а базидиях, сумках (до 30 мкм в диаметре) - Basidiоbоlus ranarum.
Размер ядер Гаплоидные ядра у большинства грибов - очень мелкие (1 -3 мкм). Диплоидные в 2 -2. 5 раза больше. Диплоидное ядро Verpa bogemica достигает 15 мкм. Наиболее крупные ядра встречаются в органах, связанных с половым воспроизведением, например, а базидиях, сумках (до 30 мкм в диаметре) - Basidiоbоlus ranarum.
 Форма ядер • сферические, эллипсоидальные или лопастные. Подробное изучение ядерного аппарата проводили у сумчатых грибов. Для каждого типа клеток у одного и того же дискомицета характерны свои размеры и форма ядер. Пример форма и размер ядер (мкм) у разных клеток Pustularia cupularis (Tarzetta cupularis) (Ascomycetes) в мкм: • Парафизы эллипсоидальная 4, 5 х1, 5 • Сумки сферическая до 6 • Мицелий сферические 1 В развевающихся спорах ядра могут быть эллипсоидальными, сферическими или лопастными. Причем форма ядра часто коррелирует с формой споры.
Форма ядер • сферические, эллипсоидальные или лопастные. Подробное изучение ядерного аппарата проводили у сумчатых грибов. Для каждого типа клеток у одного и того же дискомицета характерны свои размеры и форма ядер. Пример форма и размер ядер (мкм) у разных клеток Pustularia cupularis (Tarzetta cupularis) (Ascomycetes) в мкм: • Парафизы эллипсоидальная 4, 5 х1, 5 • Сумки сферическая до 6 • Мицелий сферические 1 В развевающихся спорах ядра могут быть эллипсоидальными, сферическими или лопастными. Причем форма ядра часто коррелирует с формой споры.
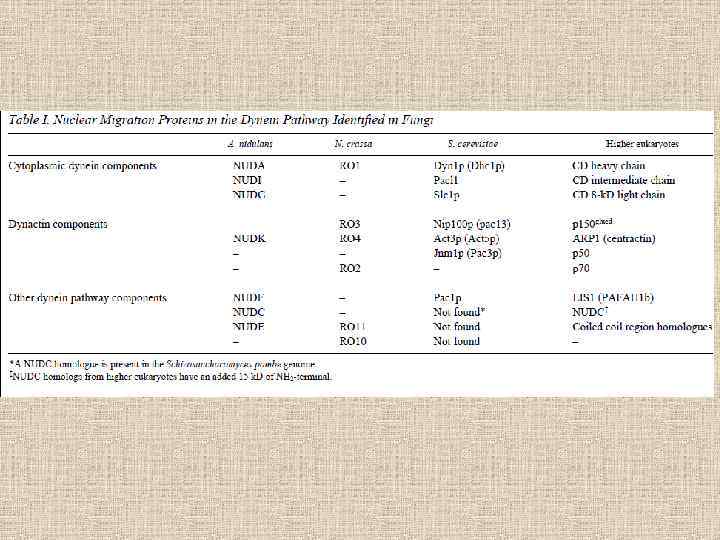
 Nuclear migration in S. cerevisiae . Successive stages of nuclear migration during bud formation in wild-type yeast cells are shown in A-C. A: The nucleus has migrated to the small bud neck. B: Mitosis occurs through the neck. C: Both daughter cells received one nucleus. D^H: Nuclear migration defects in yeast. In a kar 9 mutant cell (E) nuclei failed to move up to the bud neck and (F) anaphase occurs in the mother cell which results in (G) binucleate mothers. D and H show two kar 9, dynein double mutant cells in which a massive defect in nuclear migration results in multinucleate mothers and anucleate buds. Nuclei were stained with DAPI and a epifluorescence picture was overlaid with a phase contrast view of the same cells. The figure was kindly provided by M. D. Rose. Morris, 2000
Nuclear migration in S. cerevisiae . Successive stages of nuclear migration during bud formation in wild-type yeast cells are shown in A-C. A: The nucleus has migrated to the small bud neck. B: Mitosis occurs through the neck. C: Both daughter cells received one nucleus. D^H: Nuclear migration defects in yeast. In a kar 9 mutant cell (E) nuclei failed to move up to the bud neck and (F) anaphase occurs in the mother cell which results in (G) binucleate mothers. D and H show two kar 9, dynein double mutant cells in which a massive defect in nuclear migration results in multinucleate mothers and anucleate buds. Nuclei were stained with DAPI and a epifluorescence picture was overlaid with a phase contrast view of the same cells. The figure was kindly provided by M. D. Rose. Morris, 2000
 Строение ядер ЯО - двумембранная оболочка толщиной 7, 5 нм с порами Ядр -нуклеоплазма с хроматином -ядрышко Интерфазное ядро Agaricus bisporus (химическая фиксация)
Строение ядер ЯО - двумембранная оболочка толщиной 7, 5 нм с порами Ядр -нуклеоплазма с хроматином -ядрышко Интерфазное ядро Agaricus bisporus (химическая фиксация)
 Saccharomyces cerevisiae, Ashbya gossypii freese-substitution
Saccharomyces cerevisiae, Ashbya gossypii freese-substitution
 Субкомпартменты ядра дрожжей
Субкомпартменты ядра дрожжей
 Субкомпартменты ядра дрожжей
Субкомпартменты ядра дрожжей
 Параллельные механизмы приводят к прикреплению теломер дрожжей к ядерной оболочке. На разных стадиях клеточного цикла ассоциированные с теломерой белки осуществляют разные контакты с компонентами внутренней ядерной мембраны. Домен Sir 4 -PAD связывает Esc 1 C концы а также y. Ku 80 и Mps 3. y. Ku 80 связывает теломеразу, которая также ассоциирует с Mps 3 в S фазу через Est 1. Имеется неидентифицированный якорьдля y. Ku в G 1 фазу, который не завсист ни от Esc 1, ни от Mps 3. Taddei et al. , 2010
Параллельные механизмы приводят к прикреплению теломер дрожжей к ядерной оболочке. На разных стадиях клеточного цикла ассоциированные с теломерой белки осуществляют разные контакты с компонентами внутренней ядерной мембраны. Домен Sir 4 -PAD связывает Esc 1 C концы а также y. Ku 80 и Mps 3. y. Ku 80 связывает теломеразу, которая также ассоциирует с Mps 3 в S фазу через Est 1. Имеется неидентифицированный якорьдля y. Ku в G 1 фазу, который не завсист ни от Esc 1, ни от Mps 3. Taddei et al. , 2010
 Хроматин • Хроматин можно определить как комплекс ДНК и связанных с ней белков. У всех эукариот структурной единицей хроматина является нуклеосома – образование из 8 молекул гистонов, формирующих ядро, вокруг которого обвит участок ДНК длиной примерно 145 п. н. ДНК делает около 1, 7 оборота вокруг ядра. Как и у высших эукариот грибной хроматин сжат в нуклеосомы, но нуклеосомные повторы короче (140 п. н. ), чем у животных (200 п. н. ), при этом нуклеосомная сердцевина консервативна (145 п. н. ) по своей природе, а нуклеосомный повтор нет. •
Хроматин • Хроматин можно определить как комплекс ДНК и связанных с ней белков. У всех эукариот структурной единицей хроматина является нуклеосома – образование из 8 молекул гистонов, формирующих ядро, вокруг которого обвит участок ДНК длиной примерно 145 п. н. ДНК делает около 1, 7 оборота вокруг ядра. Как и у высших эукариот грибной хроматин сжат в нуклеосомы, но нуклеосомные повторы короче (140 п. н. ), чем у животных (200 п. н. ), при этом нуклеосомная сердцевина консервативна (145 п. н. ) по своей природе, а нуклеосомный повтор нет. •
 ДНК") • Компактная укладка происходит на нескольких уровнях. Длина 146 bp (пар оснований) ДНК комплекса с октамером из гистнных белков (две копии каждого гистона H 3, H 4, H 2 A, и H 2 B) в центре нуклеосомной частицы и нуклеосомы далее скручены в иерархии 10 nm фибрилл, 30 nm фибрилл, и хромосомные петли
• Компактная укладка происходит на нескольких уровнях. Длина 146 bp (пар оснований) ДНК комплекса с октамером из гистнных белков (две копии каждого гистона H 3, H 4, H 2 A, и H 2 B) в центре нуклеосомной частицы и нуклеосомы далее скручены в иерархии 10 nm фибрилл, 30 nm фибрилл, и хромосомные петли
 • хроматин не является статичной структурой. Хроматин участвует в ежеминутной рабочей активности ядра и имеется много белков, которые требуются для модификации и адаптации хроматина для того, чтобы поготовить ДНК к экспрессии. Гистоны ацетилируются и деацетилируются для модификации структуры хроматина.
• хроматин не является статичной структурой. Хроматин участвует в ежеминутной рабочей активности ядра и имеется много белков, которые требуются для модификации и адаптации хроматина для того, чтобы поготовить ДНК к экспрессии. Гистоны ацетилируются и деацетилируются для модификации структуры хроматина.
 1 2 3 4 5") Разные уровни упаковки хроматина (Perez-Martin, 1999) 1 2 3 4 5
Разные уровни упаковки хроматина (Perez-Martin, 1999) 1 2 3 4 5
") Структурная модель взаимодействия гистонов с ДНК (Perez-Martin, 1999)
Структурная модель взаимодействия гистонов с ДНК (Perez-Martin, 1999)
 • Молекулы ДНК очень длинные. Например, III дрожжевая хромосома, полностью секвенированная первой содержит 3. 15 × 105 оснований и, так как соседние основания разделены пространством 0. 34 nm, хромосомальная молекула может достигать около 107 μm длиной. Дрожжевые клетки варьируют по размеру, но в среднем они имеют размер 5 - 7 μm; то есть одна молекула ДНК примерно в 15 -20 раз длиннее самих клеток, которые ее содержат. Молекула ДНК 2 nm в диаметре за счет высокой конденсации хроматина
• Молекулы ДНК очень длинные. Например, III дрожжевая хромосома, полностью секвенированная первой содержит 3. 15 × 105 оснований и, так как соседние основания разделены пространством 0. 34 nm, хромосомальная молекула может достигать около 107 μm длиной. Дрожжевые клетки варьируют по размеру, но в среднем они имеют размер 5 - 7 μm; то есть одна молекула ДНК примерно в 15 -20 раз длиннее самих клеток, которые ее содержат. Молекула ДНК 2 nm в диаметре за счет высокой конденсации хроматина
 Хромосомы и кариотип • Термин «хромосома» от греческого chroma окрашенный. После того как стало известно что ДНК – носитель информации понятие «хромомсома» стали применять ко всем структурам несущим гены и даже к пластидам и митохондриям. Совокупность генов в клетке геном (греч genos – потомство, род). Наряду с ядерным геномом (нуклеомом) различают хондром (=хондриом) митохондрий, который меньше нуклеома. Размер генома дается как общее число пар оснований ДНК.
Хромосомы и кариотип • Термин «хромосома» от греческого chroma окрашенный. После того как стало известно что ДНК – носитель информации понятие «хромомсома» стали применять ко всем структурам несущим гены и даже к пластидам и митохондриям. Совокупность генов в клетке геном (греч genos – потомство, род). Наряду с ядерным геномом (нуклеомом) различают хондром (=хондриом) митохондрий, который меньше нуклеома. Размер генома дается как общее число пар оснований ДНК.
 • Набор хромосом, содержащийся в ядрах представителей одного вида называют кариотипом. Этот термин охватывает всю совокупность свойств хромомсом, рассматриваемых цитологией (размер, число, облик). Это особенно важный признак для генетики, систематики и филогении. Число однородных хромосомных наборов в ядре определяет степень его плоидности (n). Ядра с одинарным набором хромосом являются гаплоидными.
• Набор хромосом, содержащийся в ядрах представителей одного вида называют кариотипом. Этот термин охватывает всю совокупность свойств хромомсом, рассматриваемых цитологией (размер, число, облик). Это особенно важный признак для генетики, систематики и филогении. Число однородных хромосомных наборов в ядре определяет степень его плоидности (n). Ядра с одинарным набором хромосом являются гаплоидными.
 Число хромосом в ядрах разных видов грибов Вид гриба № п 1 Achlya 3 2 Agaricus bisporus 14 3 Allomyces 16 4 Aspergillus nidulans 8 5 Candida utilis 2 6 Neurospora crassa 7 8 Pleurotus ostreatus 11 9 P. pulmonarius 8 10 Saccharomyces cerevisiae 16
Число хромосом в ядрах разных видов грибов Вид гриба № п 1 Achlya 3 2 Agaricus bisporus 14 3 Allomyces 16 4 Aspergillus nidulans 8 5 Candida utilis 2 6 Neurospora crassa 7 8 Pleurotus ostreatus 11 9 P. pulmonarius 8 10 Saccharomyces cerevisiae 16
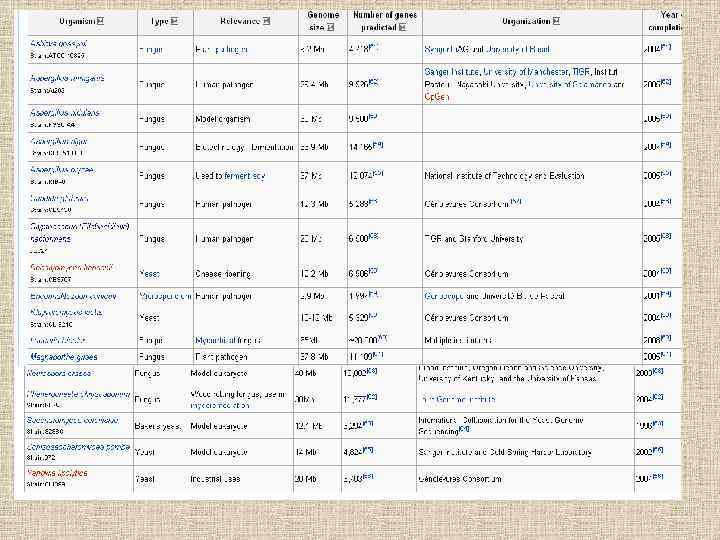
 Rhizopus oryzae 35 Saccharomyces") Примерный размер генома у представленных эукариот Organism Genome size (Mb*) Rhizopus oryzae 35 Saccharomyces cerevisiae 12 Aspergillus nidulans 31 Neurospora crassa 39 Coprinopsis cinerea 36 Ustilago maydis 20 Drosophila melanogaster 122 Sea Urchin 814 Human 3 300
Примерный размер генома у представленных эукариот Organism Genome size (Mb*) Rhizopus oryzae 35 Saccharomyces cerevisiae 12 Aspergillus nidulans 31 Neurospora crassa 39 Coprinopsis cinerea 36 Ustilago maydis 20 Drosophila melanogaster 122 Sea Urchin 814 Human 3 300
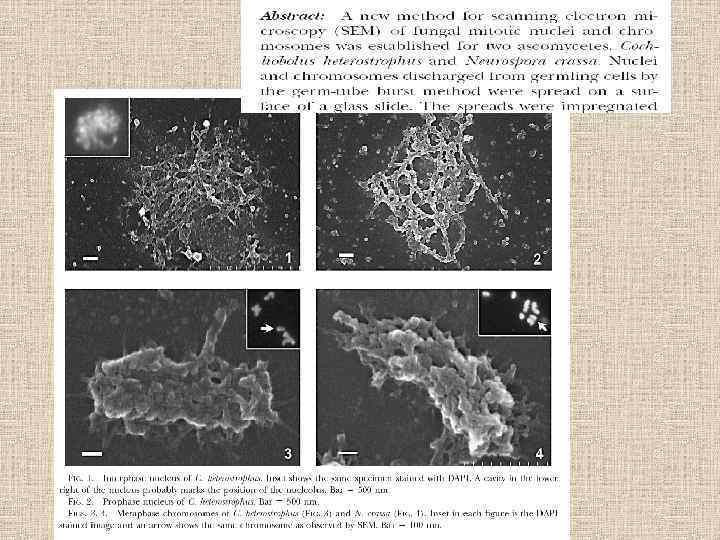
 Наблюдение хромосом Fig. la, b. Fluorescence staining of chromosomes before and after their discharge from a germ tube cell of Botrytis cinema, a Clumps of condensed chromosomes, presumably at metaphase, in an intact germ tube cell. Conidia were incubated to germinate for 7. 5 h and subjected to 4", 6 -diamidino-2 -phenylindole (DAPI) staining, b Chromosomes discharged by the germ tube burst method. Each discrete group of chromosomes presumably comes from a single nucleus. Long, slender chromatid threads (arrows) were shown to be the site of the nucleolus organizer region (NOR) by fluorescence in situ hybridization (FISH). Bar represents 5 lam
Наблюдение хромосом Fig. la, b. Fluorescence staining of chromosomes before and after their discharge from a germ tube cell of Botrytis cinema, a Clumps of condensed chromosomes, presumably at metaphase, in an intact germ tube cell. Conidia were incubated to germinate for 7. 5 h and subjected to 4", 6 -diamidino-2 -phenylindole (DAPI) staining, b Chromosomes discharged by the germ tube burst method. Each discrete group of chromosomes presumably comes from a single nucleus. Long, slender chromatid threads (arrows) were shown to be the site of the nucleolus organizer region (NOR) by fluorescence in situ hybridization (FISH). Bar represents 5 lam
 Fig. 2 a-e. Chromosome composition of the genome of B. cinerea.") Fluorescent dye (DAPI) Fig. 2 a-e. Chromosome composition of the genome of B. cinerea. a Sixteen chromosomes, the usual number observed in this fungus, can be counted, b Thirteen chromosomes, an unusual number, can be counted, e Prominent chromatin thread stemming from a relatively small chromosome. Arrows indicate centromere-like constrictions. Bar represents 2 gm
Fluorescent dye (DAPI) Fig. 2 a-e. Chromosome composition of the genome of B. cinerea. a Sixteen chromosomes, the usual number observed in this fungus, can be counted, b Thirteen chromosomes, an unusual number, can be counted, e Prominent chromatin thread stemming from a relatively small chromosome. Arrows indicate centromere-like constrictions. Bar represents 2 gm
 Fig. 4 a-e. FISH of r. DNAs") FISH (метод флуоресцентной гибридизации in situ ) Fig. 4 a-e. FISH of r. DNAs in B. cinerea and A. alternata, a-c FISH in B. cinerea, d, e FISH in A. alternata. Fluorescein isothiocyanate (FITC) signals take various shapes, from heavy dot to rod (a, d), slender thread (b), or long string of beads (e, e). Bar represents 2 gm Связывании ДНК-зонда с хромосомной ДНК исследуемого образца. Зонд представляет собой небольшой фрагмент ДНК, помеченный флуоресцентным красителем, который связывается с определенным участком хромосомы.
FISH (метод флуоресцентной гибридизации in situ ) Fig. 4 a-e. FISH of r. DNAs in B. cinerea and A. alternata, a-c FISH in B. cinerea, d, e FISH in A. alternata. Fluorescein isothiocyanate (FITC) signals take various shapes, from heavy dot to rod (a, d), slender thread (b), or long string of beads (e, e). Bar represents 2 gm Связывании ДНК-зонда с хромосомной ДНК исследуемого образца. Зонд представляет собой небольшой фрагмент ДНК, помеченный флуоресцентным красителем, который связывается с определенным участком хромосомы.
") Хромосомы Uromyces phaseoli (Heath, 1976)
Хромосомы Uromyces phaseoli (Heath, 1976)
 Cytological and electrophoretical examination of Nectria haematococca chromosomes
Cytological and electrophoretical examination of Nectria haematococca chromosomes
 белковый комплекс примерно 50 MDa содержит 456 нуклеопоринов 30") Ядерные поры Saccharomyces cerevisiae (200) белковый комплекс примерно 50 MDa содержит 456 нуклеопоринов 30 разных типов.
Ядерные поры Saccharomyces cerevisiae (200) белковый комплекс примерно 50 MDa содержит 456 нуклеопоринов 30 разных типов.
 Ядерные поры Schizosaccharomyces pombe
Ядерные поры Schizosaccharomyces pombe
 CF, cytoplasmic filaments; CRS, cytoplasmic ring subunits;") Ядерные поры (Kiseleva et al. , 2004) CF, cytoplasmic filaments; CRS, cytoplasmic ring subunits; TR, thin ring; ONM, outer nuclear membrane; IF, internal filaments; MR, membrane ring; T, transporter; ISD, inner spoke domain; INM, inner nuclear membrane; NR, nucleoplasmic ring; B, basket. Colored components are those observed by fe. SEM, although the basket had previously been indicated and the nucleoplasmic internal filaments remain a speculation. Bar 50 nm.
Ядерные поры (Kiseleva et al. , 2004) CF, cytoplasmic filaments; CRS, cytoplasmic ring subunits; TR, thin ring; ONM, outer nuclear membrane; IF, internal filaments; MR, membrane ring; T, transporter; ISD, inner spoke domain; INM, inner nuclear membrane; NR, nucleoplasmic ring; B, basket. Colored components are those observed by fe. SEM, although the basket had previously been indicated and the nucleoplasmic internal filaments remain a speculation. Bar 50 nm.
 • В общем случае трехмерная конструкция NPC консервативна от дрожжей до высших эукариот, хотя дрожжевой NPC мельче, чем у позвоночных. Белковые компоненты NPCs называются нуклеопоринами; дрожжевой NPC состоит из 30 -50 разных нуклеопоринов (до 100 у позвоночных).
• В общем случае трехмерная конструкция NPC консервативна от дрожжей до высших эукариот, хотя дрожжевой NPC мельче, чем у позвоночных. Белковые компоненты NPCs называются нуклеопоринами; дрожжевой NPC состоит из 30 -50 разных нуклеопоринов (до 100 у позвоночных).
 • Ядерные поры представляют собой комплексы, встроенные в двойную мембрану ядерной оболочки. Они снабжены каналами (около 9 nm в диаметре), которые пропускают ионы и маленькие молекулы (меньше, чем 50 k. Da) путем диффузии, но белки, RNAs, и рибонуклеопротеиновые частицы (RNP) крупнее 9 nm селективно транспортируются через NPCs энергозависимым механизмом. Транспортные потоки селективно регулируются сигналами развития и окружающими сигналами.
• Ядерные поры представляют собой комплексы, встроенные в двойную мембрану ядерной оболочки. Они снабжены каналами (около 9 nm в диаметре), которые пропускают ионы и маленькие молекулы (меньше, чем 50 k. Da) путем диффузии, но белки, RNAs, и рибонуклеопротеиновые частицы (RNP) крупнее 9 nm селективно транспортируются через NPCs энергозависимым механизмом. Транспортные потоки селективно регулируются сигналами развития и окружающими сигналами.
 • Транскрипция рибосомальной RNA , процессинг и сборка рибосом важные аспекты функции ядра. Все эти процессы происходят в ядрышке. • ОНО не является особой органеллой, оно является продуктом активности отдельных локусов хромосом и является центром синтеза рибосомальной РНК в клетке. Выявляется в виде особой электронноплотной зоны и состоит из гранул и фибрилл. У активных клеток (крючки, сумки, молодые споры) оно составляет до 60 % от объема ядра (аскомицеты). Оно чаще занимает эксцентрическое, реже - центральное положение. Форма ядрышка обычно сферическая. Построено оно из двух основных компонентов - гранулярного и фибриллярного. Гранулярный компонент локализован по периферии, фибриллярный - в центре. Гранулярный компонент состоит из гранул диаметром 15 -20 нм - предшественники рибосом (эти гранулы мельче рибосом цитоплазмы). Фибриллярный компонент - фибриллы РНП 68 нм. Предполагается, что гранулярный компонент происходит из фибриллярного.
• Транскрипция рибосомальной RNA , процессинг и сборка рибосом важные аспекты функции ядра. Все эти процессы происходят в ядрышке. • ОНО не является особой органеллой, оно является продуктом активности отдельных локусов хромосом и является центром синтеза рибосомальной РНК в клетке. Выявляется в виде особой электронноплотной зоны и состоит из гранул и фибрилл. У активных клеток (крючки, сумки, молодые споры) оно составляет до 60 % от объема ядра (аскомицеты). Оно чаще занимает эксцентрическое, реже - центральное положение. Форма ядрышка обычно сферическая. Построено оно из двух основных компонентов - гранулярного и фибриллярного. Гранулярный компонент локализован по периферии, фибриллярный - в центре. Гранулярный компонент состоит из гранул диаметром 15 -20 нм - предшественники рибосом (эти гранулы мельче рибосом цитоплазмы). Фибриллярный компонент - фибриллы РНП 68 нм. Предполагается, что гранулярный компонент происходит из фибриллярного.
 • Ядрышки – это места биосинтеза рибосом. Каждое ядрышко пронизано отрезком хромосомной ДНК, который обозначается как область ядрышкового организатора (nucleolus organizer region, NOR) и несет повторяющиеся гены для р. РНК. • Размер ядрышек соответствует интенсивности биосинтеза белка в клетке. В клетках, которые не синтезируют белков (генеративные) ядрышки очень мелкие.
• Ядрышки – это места биосинтеза рибосом. Каждое ядрышко пронизано отрезком хромосомной ДНК, который обозначается как область ядрышкового организатора (nucleolus organizer region, NOR) и несет повторяющиеся гены для р. РНК. • Размер ядрышек соответствует интенсивности биосинтеза белка в клетке. В клетках, которые не синтезируют белков (генеративные) ядрышки очень мелкие.
 Schizosccharomyces pombe Ashbya gossypii") Полярное тело веретена (ПТВ, spindle pole body, SPB) Schizosccharomyces pombe Ashbya gossypii
Полярное тело веретена (ПТВ, spindle pole body, SPB) Schizosccharomyces pombe Ashbya gossypii
 В 1897 году Harper, что SPBs присутствуют около ядерной оболочке в интерфазе. Позднее он сообщил, что интерфазные SPBs характерны для всех грибов и обнаружены на всех стадиях жизненного цикла аскомицетов (Harper, 1905). Kniep (1915 год) проиллюстрировал интерфазные SPBs у базидиомицетов.
В 1897 году Harper, что SPBs присутствуют около ядерной оболочке в интерфазе. Позднее он сообщил, что интерфазные SPBs характерны для всех грибов и обнаружены на всех стадиях жизненного цикла аскомицетов (Harper, 1905). Kniep (1915 год) проиллюстрировал интерфазные SPBs у базидиомицетов.
 Поведение ПТВ в процессе клеточного цикла у Schizosaccharomyces pombe
Поведение ПТВ в процессе клеточного цикла у Schizosaccharomyces pombe
") Полярные тела веретена (spindle pole body)
Полярные тела веретена (spindle pole body)
 Митоз-механизм, при помощи которого материал ядра равномерно распределяется между дочерними клетками
Митоз-механизм, при помощи которого материал ядра равномерно распределяется между дочерними клетками
 Клеточный цикл почкующихся дрожжей может быть суммирован следующим образом и те же основные принципы применимы к клеточным циклам всех эукариот (Field et al. , 1999): • На ‘start’, активируется транскрипция и трансляция циклинов G 1 стадии CLN 1 и CLN 2, и эти белки комбинируются с Cdc 28 для формирования протеин киназы которая инициирует формирование почки и подготовку клетки для S-phase (G 1 к S transition). Это позднее включит активацию генов, ответственных за циклины CLB 5 и CLB 6, которые возвращают CLN 1 и CLN 2 как партнеры Cdc 28 для содействия синтеза DNA в процессе S фазы. • К концу S фазы, запускается синтез ‘поздних’ циклинов CLB 1 и CLB 2, которые выключают транскрипцию генов ‘ранних’ циклинов, и так как эти ‘поздние’ циклины аккумулируются, они трансформируют киназу Cdc 28 в митотическую форму, облегчающую переход от G 2 к M. • Инициация митоза у клеток эукариот регулируется каскадом фосфорилирования, кульминация которого в активации mitosis promoting factor (MPF). • MPF состоит как минимум из циклин зависимой киназы Cdc 28/Cdc 2 у дрожжей, Cdk 1 у высших эукариот и B-тип циклин регуляторной субъединицы (Ohi & Gould, 1999). • Они вводят/takes клетку в телофазу, после которой активируется протеаза, выжившие циклины деградируют и цикл завершается. • в конце митоза, митотические циклины деградируют протеазами, которые остаются активными до ‘start’. Это защищает клетку от немедленного митотического пред-пуска.
Клеточный цикл почкующихся дрожжей может быть суммирован следующим образом и те же основные принципы применимы к клеточным циклам всех эукариот (Field et al. , 1999): • На ‘start’, активируется транскрипция и трансляция циклинов G 1 стадии CLN 1 и CLN 2, и эти белки комбинируются с Cdc 28 для формирования протеин киназы которая инициирует формирование почки и подготовку клетки для S-phase (G 1 к S transition). Это позднее включит активацию генов, ответственных за циклины CLB 5 и CLB 6, которые возвращают CLN 1 и CLN 2 как партнеры Cdc 28 для содействия синтеза DNA в процессе S фазы. • К концу S фазы, запускается синтез ‘поздних’ циклинов CLB 1 и CLB 2, которые выключают транскрипцию генов ‘ранних’ циклинов, и так как эти ‘поздние’ циклины аккумулируются, они трансформируют киназу Cdc 28 в митотическую форму, облегчающую переход от G 2 к M. • Инициация митоза у клеток эукариот регулируется каскадом фосфорилирования, кульминация которого в активации mitosis promoting factor (MPF). • MPF состоит как минимум из циклин зависимой киназы Cdc 28/Cdc 2 у дрожжей, Cdk 1 у высших эукариот и B-тип циклин регуляторной субъединицы (Ohi & Gould, 1999). • Они вводят/takes клетку в телофазу, после которой активируется протеаза, выжившие циклины деградируют и цикл завершается. • в конце митоза, митотические циклины деградируют протеазами, которые остаются активными до ‘start’. Это защищает клетку от немедленного митотического пред-пуска.

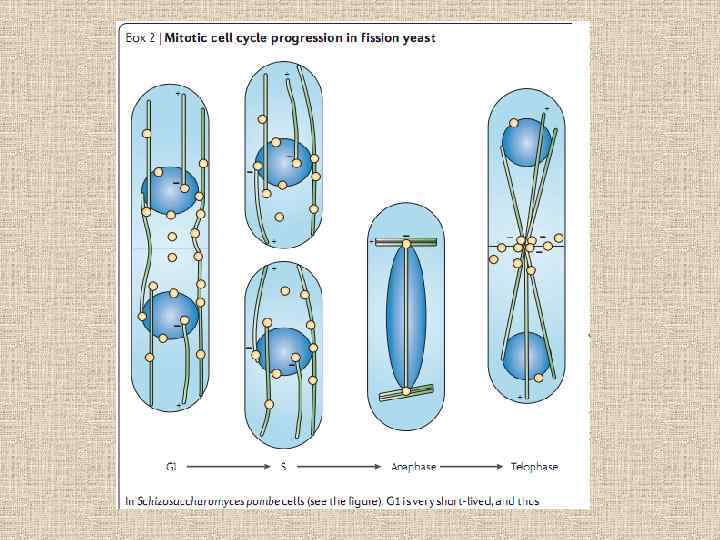
 Особенности митоза у грибов. • Ядерная оболочка не исчезает и не появляется заново, а лишь перетягивается между двумя дочерними ядрами и внутри нее образуется аппарат веретена. Закрытый митоз. Исключения – Ustilago maydis. • Центриоли у грибов отсутствуют, центры организации веретена деления - полярные тела веретена (SPB), имеющие разную организацию. • Нет метафазной пластинки • Особенности поведения ядрышка (таблица)
Особенности митоза у грибов. • Ядерная оболочка не исчезает и не появляется заново, а лишь перетягивается между двумя дочерними ядрами и внутри нее образуется аппарат веретена. Закрытый митоз. Исключения – Ustilago maydis. • Центриоли у грибов отсутствуют, центры организации веретена деления - полярные тела веретена (SPB), имеющие разную организацию. • Нет метафазной пластинки • Особенности поведения ядрышка (таблица)
 Центр организации веретена деления Поведение ядрышка Центриоли Ядрышко Plasmodiophora То же P Blastocladiella То же ? Saprolegnia То же p Achlya То же p Phytophthora То же p Mucor ПТВ p Dipodascus ПТВ di Aspergillus ПТВ di Neurospora ПТВ p Uromyces ПТВ di Coprinus ПТВ d Armillaria ПТВ d Род Physarum p - присутствует во время митоза, d - исчезает в профазу-метафазу, di - выбрасывается в цитоплазму. ПТВ – полярные тела веретена
Центр организации веретена деления Поведение ядрышка Центриоли Ядрышко Plasmodiophora То же P Blastocladiella То же ? Saprolegnia То же p Achlya То же p Phytophthora То же p Mucor ПТВ p Dipodascus ПТВ di Aspergillus ПТВ di Neurospora ПТВ p Uromyces ПТВ di Coprinus ПТВ d Armillaria ПТВ d Род Physarum p - присутствует во время митоза, d - исчезает в профазу-метафазу, di - выбрасывается в цитоплазму. ПТВ – полярные тела веретена
 a prophase nucleus of an ascomycete, Ceratocystis fagacearum early prophase of mitosis") МИТОЗ (профаза) a prophase nucleus of an ascomycete, Ceratocystis fagacearum early prophase of mitosis in a living hyphal cell of an ascomycete, Nectria haematococca
МИТОЗ (профаза) a prophase nucleus of an ascomycete, Ceratocystis fagacearum early prophase of mitosis in a living hyphal cell of an ascomycete, Nectria haematococca
 МИТОЗ (a, d, g - метафаза; b, e, h-анафаза A; c, f, I - анафаза B удлинение центрального веретена за счет действия астральных мкр/тр) middle stages of mitosis in vegetative hyphae of three filamentous fungi —Aspergillus nidulans (a–c), Nectria haematococca (d–f), and Helicobasidium mompa (g–i)—visualized in vivo by phase-contrast videomicroscopy.
МИТОЗ (a, d, g - метафаза; b, e, h-анафаза A; c, f, I - анафаза B удлинение центрального веретена за счет действия астральных мкр/тр) middle stages of mitosis in vegetative hyphae of three filamentous fungi —Aspergillus nidulans (a–c), Nectria haematococca (d–f), and Helicobasidium mompa (g–i)—visualized in vivo by phase-contrast videomicroscopy.
 Почкующиеся дрожжи Псевдогифальный рост у Candida
Почкующиеся дрожжи Псевдогифальный рост у Candida
 Анафаза
Анафаза
 Ustilago maydis Aspergillus nidulans
Ustilago maydis Aspergillus nidulans
 Моторные белки и митоз
Моторные белки и митоз
 Образцы деления ядер в многоядерных гифах грибов. Ceratocystis fagacearum Aspergillus nidulans Neurospora crassa, Ashbya gosssypii
Образцы деления ядер в многоядерных гифах грибов. Ceratocystis fagacearum Aspergillus nidulans Neurospora crassa, Ashbya gosssypii
 Aсинхронный митоз у Ashbia gossipia
Aсинхронный митоз у Ashbia gossipia
 Роль циклинов в асинхронном митозе
Роль циклинов в асинхронном митозе
") Микроавтофагия ядра Saccharomyces cerevisiae (Kvam and Goldfarb, 2006)
Микроавтофагия ядра Saccharomyces cerevisiae (Kvam and Goldfarb, 2006)

 профаза метафаза анафаза телофаза общее время 15 -60 1") Продолжительность митоза у грибов (мин) профаза метафаза анафаза телофаза общее время 15 -60 1 -5 2 -5 Род гриба Physarum 1 -5 30 Mucor 100 Saccharomyces 10 -15 Aspergillus 6 -8 Neurospora 10 Fusarum 1. 2 2 Ustilago 12 5 разные гомобазидиом. 0. 2 2. 1 28 5. 5 45 7 -45
Продолжительность митоза у грибов (мин) профаза метафаза анафаза телофаза общее время 15 -60 1 -5 2 -5 Род гриба Physarum 1 -5 30 Mucor 100 Saccharomyces 10 -15 Aspergillus 6 -8 Neurospora 10 Fusarum 1. 2 2 Ustilago 12 5 разные гомобазидиом. 0. 2 2. 1 28 5. 5 45 7 -45
 Время митоза у Fusarium oxysporum
Время митоза у Fusarium oxysporum
, Профаза митоза (6 -7)") Fusarium oxysporum Ядро перед делением (5), Профаза митоза (6 -7)
Fusarium oxysporum Ядро перед делением (5), Профаза митоза (6 -7)
 Fusarium oxysporum Митоз, метафаза
Fusarium oxysporum Митоз, метафаза
 Fusarium oxysporum Митоз, метафаза
Fusarium oxysporum Митоз, метафаза
 Fusarium oxysporum Митоз, анафаза
Fusarium oxysporum Митоз, анафаза
 Fusarium oxysporum Схема митоза
Fusarium oxysporum Схема митоза
 Коньгационный митоз у Auricularia auricular-judae (он занимает 42 мин. : 5 мин. – образование септы, 23 мин. – слияние пряжки и гифы и 13 мин – миграция ядра в гифу).
Коньгационный митоз у Auricularia auricular-judae (он занимает 42 мин. : 5 мин. – образование септы, 23 мин. – слияние пряжки и гифы и 13 мин – миграция ядра в гифу).
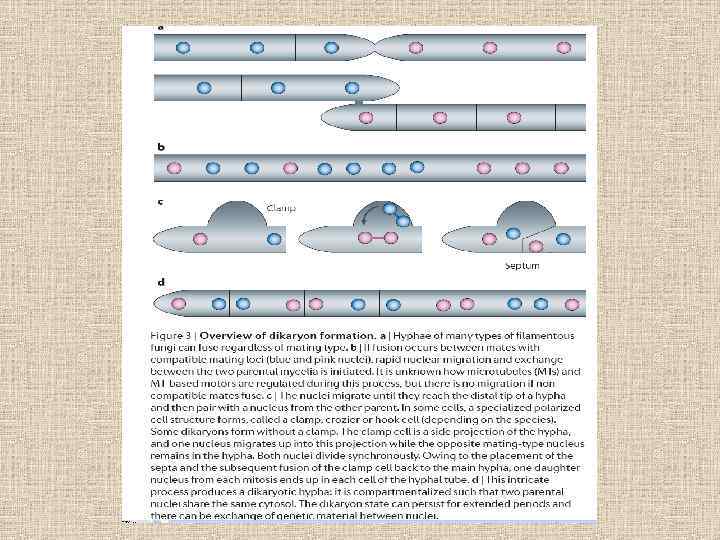
 • Баланс родительских генотипов в гетерокарионах варьирует около 1/1 в зависимости от условий окружающей среды. Работа на Heterobasidion parviporum показала, что индивидуальные ядра находятся под действием отбора. Кроме того, ядра с разными генотипами или разными эпигенетическими состояниями также различны в скорости репликации и миграции,
• Баланс родительских генотипов в гетерокарионах варьирует около 1/1 в зависимости от условий окружающей среды. Работа на Heterobasidion parviporum показала, что индивидуальные ядра находятся под действием отбора. Кроме того, ядра с разными генотипами или разными эпигенетическими состояниями также различны в скорости репликации и миграции,
 • Давно известно, что ядра в дикарионах могут общаться: обмен генетическим материалом и соматическая рекомбинация встречается между генотипами. На положение двух ядер в дикарионе влияют специфические гены, которые экспрессируются в этих ядрах. Например, у S. commune, когда ядра в дикарионе близко расположены (<2 микрон между ними), то экспрессируются гидрофобин-кодирующие гены, отличные от тех, которые экспрессируются при большей дистанции между ядрами. Таким образом, точное положение родительских ядер в дикарионе может быть деталью транскрипционной программы клетки.
• Давно известно, что ядра в дикарионах могут общаться: обмен генетическим материалом и соматическая рекомбинация встречается между генотипами. На положение двух ядер в дикарионе влияют специфические гены, которые экспрессируются в этих ядрах. Например, у S. commune, когда ядра в дикарионе близко расположены (<2 микрон между ними), то экспрессируются гидрофобин-кодирующие гены, отличные от тех, которые экспрессируются при большей дистанции между ядрами. Таким образом, точное положение родительских ядер в дикарионе может быть деталью транскрипционной программы клетки.
 митоза и последовательность событий наблюдаемых в живых гифах Auricularia auricular-judae (Lǚ,") Время (мин. ) митоза и последовательность событий наблюдаемых в живых гифах Auricularia auricular-judae (Lǚ, Mc. Laughlin, 1995) интерв ал среднее ± откл. № Одно ядро двигается в пряжку, ядрышко исчезает 4 - 12 11, 3 ± 2, 0 11 Ядрышки появляются 3 -6 4, 5 ± 0, 86 9 Септа инициируется 5 -7 4, 6 ± 1, 70 9 Септа полностью сформирована 3 -6 4, 3 – 1, 03 9 Септа сливается с нижележащей клеткой 21 - 24 22, 7 ± 1, 1 3 Восстановление двуядерности 11 - 17 14 ± 2, 0 3 событие
Время (мин. ) митоза и последовательность событий наблюдаемых в живых гифах Auricularia auricular-judae (Lǚ, Mc. Laughlin, 1995) интерв ал среднее ± откл. № Одно ядро двигается в пряжку, ядрышко исчезает 4 - 12 11, 3 ± 2, 0 11 Ядрышки появляются 3 -6 4, 5 ± 0, 86 9 Септа инициируется 5 -7 4, 6 ± 1, 70 9 Септа полностью сформирована 3 -6 4, 3 – 1, 03 9 Септа сливается с нижележащей клеткой 21 - 24 22, 7 ± 1, 1 3 Восстановление двуядерности 11 - 17 14 ± 2, 0 3 событие




