

ХРОМОСОМЫ ЧЕЛОВЕКА.pptx
- Количество слайдов: 44
 ХРОМОСОМЫ ЧЕЛОВЕКА
ХРОМОСОМЫ ЧЕЛОВЕКА
 I. Развитие представлений о хромосомах человека 70 -е годы XIX века - О. Гертвиг и В. Флемминг описали окрашиваемые нитевидные структуры в ядрах клеток эукариот 1879 г. – Дж. Арнольд (J. Arnold) – первая визуализация хромосом человека 1888 г. - В. Вальдейер (W. Waldeyer) назвал эти структуры ХРОМОСОМАМИ (chroмa –цвет; soma – тело) 1904 г. – В. Сеттон (W. Sutton) и Т. Бовери (Th. Boveri)– гипотеза о связи хромосом с элементами наследственности
I. Развитие представлений о хромосомах человека 70 -е годы XIX века - О. Гертвиг и В. Флемминг описали окрашиваемые нитевидные структуры в ядрах клеток эукариот 1879 г. – Дж. Арнольд (J. Arnold) – первая визуализация хромосом человека 1888 г. - В. Вальдейер (W. Waldeyer) назвал эти структуры ХРОМОСОМАМИ (chroмa –цвет; soma – тело) 1904 г. – В. Сеттон (W. Sutton) и Т. Бовери (Th. Boveri)– гипотеза о связи хромосом с элементами наследственности
 I этап - поиск адекватных подходов и разработка методик для проведенияя анализа морфологии хромосом Г. А. Левитский (1878 -1942) - ввел термин «кариотип» в современном его понимании. Автор одного из первых в мире учебников по цитогенетике М. С. Навашин (1896— 1973) - автор исследований по морфологии клеточного ядра, индивидуальной изменчивости хромосом в эволюционном аспекте, цитологии отдаленных гибридов. Изучал динамику клеточного деления и влияния на клетку физических и химических факторов. А. Г. Андрес (совместно с М. С. Навашиным) –впервые в мире провели анализ тонкого морфологического строения десяти наиболее крупных хромосом человека. Автор книги «Введение в кариологию человека» (1934 г. ) П. И. Живаго (1883 -1948) - Основные работы по изучению тонкого строения клеточного ядра. Обнаружил оптическую гетерогенность интерфазных ядер; исследовал строение и функцию ядрышка и показал его большую роль в обменных процессах клетки.
I этап - поиск адекватных подходов и разработка методик для проведенияя анализа морфологии хромосом Г. А. Левитский (1878 -1942) - ввел термин «кариотип» в современном его понимании. Автор одного из первых в мире учебников по цитогенетике М. С. Навашин (1896— 1973) - автор исследований по морфологии клеточного ядра, индивидуальной изменчивости хромосом в эволюционном аспекте, цитологии отдаленных гибридов. Изучал динамику клеточного деления и влияния на клетку физических и химических факторов. А. Г. Андрес (совместно с М. С. Навашиным) –впервые в мире провели анализ тонкого морфологического строения десяти наиболее крупных хромосом человека. Автор книги «Введение в кариологию человека» (1934 г. ) П. И. Живаго (1883 -1948) - Основные работы по изучению тонкого строения клеточного ядра. Обнаружил оптическую гетерогенность интерфазных ядер; исследовал строение и функцию ядрышка и показал его большую роль в обменных процессах клетки.
 II этап – определение модального числа хромосом у человека и детальное описание их морфологии (1956 г. ) J-H. Tjio (1919– 2001) A. Levan (1905– 1998).
II этап – определение модального числа хромосом у человека и детальное описание их морфологии (1956 г. ) J-H. Tjio (1919– 2001) A. Levan (1905– 1998).

 идентифицировал трисомию по хромосоме 21") • 1959 г. – Ж. Лежен (J. Lejeune) идентифицировал трисомию по хромосоме 21 при синдроме Дауна • 1960 г. – Х. Эдвардс (H. Edwards) описал синдром трисомии по хромосоме 18 • 1960 г. – К. Патау (K. Patau) описал синдром трисомии по хромосоме 13 • Рождение медицинской цитогенетики!
• 1959 г. – Ж. Лежен (J. Lejeune) идентифицировал трисомию по хромосоме 21 при синдроме Дауна • 1960 г. – Х. Эдвардс (H. Edwards) описал синдром трисомии по хромосоме 18 • 1960 г. – К. Патау (K. Patau) описал синдром трисомии по хромосоме 13 • Рождение медицинской цитогенетики!
 III этап – появление методов идентификации хромосом и их отдельных районов (конец 60 -х – начало 70 -х годов XX в. ) • 1968 г. – Т. Касперсон (T. Caspersson) применил для исследования метафазных хромосом метод дифференциальной Q-окраски • 1971 г. – М. Дретс (M. Drets) и М. Шау (M. Shaw) использовали GTG-метод дифференциальной окраски хромосом
III этап – появление методов идентификации хромосом и их отдельных районов (конец 60 -х – начало 70 -х годов XX в. ) • 1968 г. – Т. Касперсон (T. Caspersson) применил для исследования метафазных хромосом метод дифференциальной Q-окраски • 1971 г. – М. Дретс (M. Drets) и М. Шау (M. Shaw) использовали GTG-метод дифференциальной окраски хромосом
 А. А. Прокофьева-Бельговская внесла огромный вклад в разработку новых методов анализа хромосом, позволяющих оценить их роль в патологии человека. Цитогенетик с мировым именем, она внесла значительный вклад в исследование проблемы организации эукариотической хромосомы. Основатель советской школы цитогенетики человека и медицинской цитогенетики. На протяжении нескольких десятилетий она была ведущим цитогенетиком нашей страны. «Гетерохроматические А. А. Прокофьева-Бельговская (1903 -1984) районы хромосом» . - Наука – Москва, 1986 г.
А. А. Прокофьева-Бельговская внесла огромный вклад в разработку новых методов анализа хромосом, позволяющих оценить их роль в патологии человека. Цитогенетик с мировым именем, она внесла значительный вклад в исследование проблемы организации эукариотической хромосомы. Основатель советской школы цитогенетики человека и медицинской цитогенетики. На протяжении нескольких десятилетий она была ведущим цитогенетиком нашей страны. «Гетерохроматические А. А. Прокофьева-Бельговская (1903 -1984) районы хромосом» . - Наука – Москва, 1986 г.
 Один из ведущих отечественных медицинских генетиков 70 -х – первой половины 80 -х гг. 20 в. , автор приоритетных исследований в области структурно-функциональной организации хромосом, хромосомных болезней, цитогенетики злокачественных опухолей. Разработал методы исследования линейной структуры хромосомы по неоднородности ее конденсации. Сформулировал положение о характере и природе линейной дифференцированности метафазной хромосомы. Показал роль хромосомных вариантов в патологии развития у человека. А. Ф. Захаров (1928 – 1986). Хромосомы человека. Атлас. – М. , 1982
Один из ведущих отечественных медицинских генетиков 70 -х – первой половины 80 -х гг. 20 в. , автор приоритетных исследований в области структурно-функциональной организации хромосом, хромосомных болезней, цитогенетики злокачественных опухолей. Разработал методы исследования линейной структуры хромосомы по неоднородности ее конденсации. Сформулировал положение о характере и природе линейной дифференцированности метафазной хромосомы. Показал роль хромосомных вариантов в патологии развития у человека. А. Ф. Захаров (1928 – 1986). Хромосомы человека. Атлас. – М. , 1982
 •") IV этап – внедрение и широкое использование молекулярноцитогенетических методов (с 1986 г. ) • 1986 г. - первые молекулярноцитогенетические эксперименты на хромосомах человека – FISH-метод • 1992 г. - разработка метода сравнительной геномной гибриидзации (CGH) A. Kallionieme D. Pinkel
IV этап – внедрение и широкое использование молекулярноцитогенетических методов (с 1986 г. ) • 1986 г. - первые молекулярноцитогенетические эксперименты на хромосомах человека – FISH-метод • 1992 г. - разработка метода сравнительной геномной гибриидзации (CGH) A. Kallionieme D. Pinkel
 – предложена система описания хромосом") НОМЕНКЛАТУРА ХРОМОСОМ ЧЕЛОВЕКА • Денверская конференция (1960 г. ) – предложена система описания хромосом • Лондонская конференция (1963 г. ) – официально введено разделение хромосом на 7 морфологических групп (A – G) • Парижская конференция (1971 г. )– идентификация хромосом на основе дифференциальной окраски • V Международный конгресс по генетике человека (Мехико, 1972 г. ) – появление первой официальной номенклатуры хромосом человека - «An International System for Human Cytogenetic Nomenclature» (ISCN, 1978)
НОМЕНКЛАТУРА ХРОМОСОМ ЧЕЛОВЕКА • Денверская конференция (1960 г. ) – предложена система описания хромосом • Лондонская конференция (1963 г. ) – официально введено разделение хромосом на 7 морфологических групп (A – G) • Парижская конференция (1971 г. )– идентификация хромосом на основе дифференциальной окраски • V Международный конгресс по генетике человека (Мехико, 1972 г. ) – появление первой официальной номенклатуры хромосом человека - «An International System for Human Cytogenetic Nomenclature» (ISCN, 1978)
 Группа A Хромосомы 1– 3 Описание Большие, метацентрические, равноплечие B 4, 5 Большие, субметацентрические, плечи сильно отличются по размеру C 6– 12, X Среднего размера; субметацентрические D 13– 15 Среднего размера; акроцентрические, со спутниками E 16– 18 Маленькие; 16 хромосома - метацентрческая, хромосомы 17 и 18 - субметацентрические F 19, 20 Маленькие; метацентрические G 21, 22, Y Маленькие; акроцентрические, со спутниками Маленькая; акроцентрическая, без спутников Аутосомы нумеруются от больших к меньшим, кроме хромосомы 21, которая меньше чем хромосома 22.
Группа A Хромосомы 1– 3 Описание Большие, метацентрические, равноплечие B 4, 5 Большие, субметацентрические, плечи сильно отличются по размеру C 6– 12, X Среднего размера; субметацентрические D 13– 15 Среднего размера; акроцентрические, со спутниками E 16– 18 Маленькие; 16 хромосома - метацентрческая, хромосомы 17 и 18 - субметацентрические F 19, 20 Маленькие; метацентрические G 21, 22, Y Маленькие; акроцентрические, со спутниками Маленькая; акроцентрическая, без спутников Аутосомы нумеруются от больших к меньшим, кроме хромосомы 21, которая меньше чем хромосома 22.
 окрашивание") МЕТАФАЗНЫЕ ПЛАСТИНКИ ХРОМОСОМ ЧЕЛОВЕКА Увеличение 10 х100 Сплошное (рутинное) окрашивание
МЕТАФАЗНЫЕ ПЛАСТИНКИ ХРОМОСОМ ЧЕЛОВЕКА Увеличение 10 х100 Сплошное (рутинное) окрашивание
,") ДИФФЕРЕНЦИАЛЬНОЕ ОКРАШИВАНИЕ ХРОМОСОМ 1. Методы, выявляющие поперечную исчерченность (чередование светлых и темных поперечных полос), специфичную для каждой хромосомы – Q, G и R-окрашивание. 2. Методы, селективно окрашивающие определенные участки хромосом – C, T, Ag-NOR и др.
ДИФФЕРЕНЦИАЛЬНОЕ ОКРАШИВАНИЕ ХРОМОСОМ 1. Методы, выявляющие поперечную исчерченность (чередование светлых и темных поперечных полос), специфичную для каждой хромосомы – Q, G и R-окрашивание. 2. Методы, селективно окрашивающие определенные участки хромосом – C, T, Ag-NOR и др.
 ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА
ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА
 QF QFQ QFH GT GTG GTW GAG CB CBG RF RFA RH RHG Q- флуоресцентное окрашивание Q-флуоресцентное окрашивание с акрихином Q-флуоресцентное окрашивание с Hoechst 33258 G-окрашивание с красителем Гимза (Giemsa) G-окрашивание с трипсином и красителем Гимза G-окрашивание с трипсином и красителем Райта (Wright) G-окрашивание с солями уксусной кислоты и красителем Гимза С-окрашивание с гидроксидом бария и красителем Гимза R- флуоресцентное окрашивание R-флуоресцентное окрашивание с акридином орнажевым R-окрашивание при нагревании с красителем Гимза
QF QFQ QFH GT GTG GTW GAG CB CBG RF RFA RH RHG Q- флуоресцентное окрашивание Q-флуоресцентное окрашивание с акрихином Q-флуоресцентное окрашивание с Hoechst 33258 G-окрашивание с красителем Гимза (Giemsa) G-окрашивание с трипсином и красителем Гимза G-окрашивание с трипсином и красителем Райта (Wright) G-окрашивание с солями уксусной кислоты и красителем Гимза С-окрашивание с гидроксидом бария и красителем Гимза R- флуоресцентное окрашивание R-флуоресцентное окрашивание с акридином орнажевым R-окрашивание при нагревании с красителем Гимза
 GTG -окраска
GTG -окраска
 Q -окраска
Q -окраска
 C -окраска
C -окраска
 ГЕТЕРОХРОМАТИН vs ЭУХРОМАТИН n n Эухроматин n Менее конденсированные районы хромосом n Транскрипционно активен Гетерохроматин n Плотные, компактные (конденсированные) районы хромосом, интенсивно окрашивается. n Обычно выявляются в зоне первичной перетяжки (центромера) n Транскрипционно неактивен (в целом) n Поздно реплицируется в S-фазе митоза (асинхронно с эухроматином)
ГЕТЕРОХРОМАТИН vs ЭУХРОМАТИН n n Эухроматин n Менее конденсированные районы хромосом n Транскрипционно активен Гетерохроматин n Плотные, компактные (конденсированные) районы хромосом, интенсивно окрашивается. n Обычно выявляются в зоне первичной перетяжки (центромера) n Транскрипционно неактивен (в целом) n Поздно реплицируется в S-фазе митоза (асинхронно с эухроматином)
 гетерохроматин • II. Гетерохроматизированные эухроматические районы (факультативный") Два вида гетерохроматина • I. Структурный (конституциональный) гетерохроматин • II. Гетерохроматизированные эухроматические районы (факультативный гетерохроматин)
Два вида гетерохроматина • I. Структурный (конституциональный) гетерохроматин • II. Гетерохроматизированные эухроматические районы (факультативный гетерохроматин)
 гетерохроматин n n Стабильно выявляется в хромосомах Генетически инертный (не содержит") I. Структурный (конституциональный) гетерохроматин n n Стабильно выявляется в хромосомах Генетически инертный (не содержит активно транскрибируемых генов)
I. Структурный (конституциональный) гетерохроматин n n Стабильно выявляется в хромосомах Генетически инертный (не содержит активно транскрибируемых генов)
 ТИПЫ СТРУКТУРНОГО ГЕТЕРОХРОМАТИНА Ø Центромерный гетерохроматин Ø Гетерохроматин вторичных перетяжек (1 qh, 9 qh, 16 qh) Ø Гетерохроматин коротких плеч акроцентрических хромосом Ø Гетерохроматин хромосомы Y (Yqh)
ТИПЫ СТРУКТУРНОГО ГЕТЕРОХРОМАТИНА Ø Центромерный гетерохроматин Ø Гетерохроматин вторичных перетяжек (1 qh, 9 qh, 16 qh) Ø Гетерохроматин коротких плеч акроцентрических хромосом Ø Гетерохроматин хромосомы Y (Yqh)
 16 qh+ увеличение длины гетерохроматического района длинного плеча") НОМЕНКЛАТУРА ВАРИАНТОВ ХРОМОСОМ Варибельность длины (размеров) 16 qh+ увеличение длины гетерохроматического района длинного плеча хромосомы 16 Yqh+ увеличение гетерохроматического района длинного плеча хромосомы Y. увеличение размеров спутников на коротком плече 21 рs+ хромосомы 21 увеличение длины спутничной нити на коротком 22 рstk+ плече хромосомы 22. увеличение азмера ентромерного етерохроматина р ц г 13 cenh+mat хромосомы 13, унаследованное от матери 9 qh+, 15 cenh+ 21 ps+ увеличение длины гетерохроматического района длинного плеча хромосомы 9, увеличение центромерного гетерохроматина в хромосоме 15 и большие спутники на коротком плече хромосомы 21. Центромерный гетерохроматин (h), спутничные нити (stk) спутник (s)
НОМЕНКЛАТУРА ВАРИАНТОВ ХРОМОСОМ Варибельность длины (размеров) 16 qh+ увеличение длины гетерохроматического района длинного плеча хромосомы 16 Yqh+ увеличение гетерохроматического района длинного плеча хромосомы Y. увеличение размеров спутников на коротком плече 21 рs+ хромосомы 21 увеличение длины спутничной нити на коротком 22 рstk+ плече хромосомы 22. увеличение азмера ентромерного етерохроматина р ц г 13 cenh+mat хромосомы 13, унаследованное от матери 9 qh+, 15 cenh+ 21 ps+ увеличение длины гетерохроматического района длинного плеча хромосомы 9, увеличение центромерного гетерохроматина в хромосоме 15 и большие спутники на коротком плече хромосомы 21. Центромерный гетерохроматин (h), спутничные нити (stk) спутник (s)
 n n Конденсированный и поэтому генетически инактивированный эухроматин") II. Гетерохроматизированные эухроматические районы (факультативный гетерохроматин) n n Конденсированный и поэтому генетически инактивированный эухроматин Представлен только в одной в двух гомологичных хромосом (при инактивации одной из хромосом X – тельца Барра) или в отдельных районах хромосом, близких к району структурного гетерохроматина при хромосомных перестройках
II. Гетерохроматизированные эухроматические районы (факультативный гетерохроматин) n n Конденсированный и поэтому генетически инактивированный эухроматин Представлен только в одной в двух гомологичных хромосом (при инактивации одной из хромосом X – тельца Барра) или в отдельных районах хромосом, близких к району структурного гетерохроматина при хромосомных перестройках
 • Бэнд - участок хромосомы, отличающийся от соседних по интенсивности окраски,") ХРОМОСОМНЫЕ БЭНДЫ (BANDS) • Бэнд - участок хромосомы, отличающийся от соседних по интенсивности окраски, при использовании соответствующего метода дифференциально го окрашивания.
ХРОМОСОМНЫЕ БЭНДЫ (BANDS) • Бэнд - участок хромосомы, отличающийся от соседних по интенсивности окраски, при использовании соответствующего метода дифференциально го окрашивания.
 «… Бэнды отражают структурно-функциональную организацию генома, обусловливающую регуляцию репликации ДНК, репарацию, транскрипцию и генетическую рекомбинацию. Бэнды представляют собой крупные структуры, которые могут включать сотни генов. Молекулярной основой дифференциального окрашивания хромосом является нуклеотидный и белковый состав и функциональная организация соответствующих бэндам участков генома. G-позитивные бэнды (R-негативные) содержат АТбогатую, поздно реплицирующуюся ДНК, бедную генами G-негативные бэнды (R-позитивные) GC–богаты, обогащены генами, их ДНК рано реплицируется» . ISCN, 2009
«… Бэнды отражают структурно-функциональную организацию генома, обусловливающую регуляцию репликации ДНК, репарацию, транскрипцию и генетическую рекомбинацию. Бэнды представляют собой крупные структуры, которые могут включать сотни генов. Молекулярной основой дифференциального окрашивания хромосом является нуклеотидный и белковый состав и функциональная организация соответствующих бэндам участков генома. G-позитивные бэнды (R-негативные) содержат АТбогатую, поздно реплицирующуюся ДНК, бедную генами G-негативные бэнды (R-позитивные) GC–богаты, обогащены генами, их ДНК рано реплицируется» . ISCN, 2009
 - постоянные и отчетливые морфологические особенности хромосом. Делят хромосомы на") Четко видимые ориентиры (landmarks) - постоянные и отчетливые морфологические особенности хромосом. Делят хромосомы на районы. К этим ориентирам относятся концы хромосомных плеч, центромеры и определенные, наиболее четко видимые бэнды. Нумерация бэндов и районов осуществляется в направлении от центромеры к теломере. 10
Четко видимые ориентиры (landmarks) - постоянные и отчетливые морфологические особенности хромосом. Делят хромосомы на районы. К этим ориентирам относятся концы хромосомных плеч, центромеры и определенные, наиболее четко видимые бэнды. Нумерация бэндов и районов осуществляется в направлении от центромеры к теломере. 10
 • первая цифра - номер хромосомы, в которой локализован данный бэнд; • второй символ (p или q) - плечо хромосомы; • третий символ – номер района, в состав которого входит бэнд; • четвертый символ – номер бэнда в составе района. 1 р31 - первый бэнд, локализованный в третьем районе короткого плеча хромосомы 1.
• первая цифра - номер хромосомы, в которой локализован данный бэнд; • второй символ (p или q) - плечо хромосомы; • третий символ – номер района, в состав которого входит бэнд; • четвертый символ – номер бэнда в составе района. 1 р31 - первый бэнд, локализованный в третьем районе короткого плеча хромосомы 1.
 Бэнд 1 р31: суббэнды 1 р31. 1; 1 р31. 2; 1 р31. 3, 1 р36. 33 - дистальный элемент суббэнда 1 р36. 3 300 - 400 - 550 - 700 - 850 - бэндов
Бэнд 1 р31: суббэнды 1 р31. 1; 1 р31. 2; 1 р31. 3, 1 р36. 33 - дистальный элемент суббэнда 1 р36. 3 300 - 400 - 550 - 700 - 850 - бэндов
 300 бэндов 550 бэндов 700 -бэндов
300 бэндов 550 бэндов 700 -бэндов
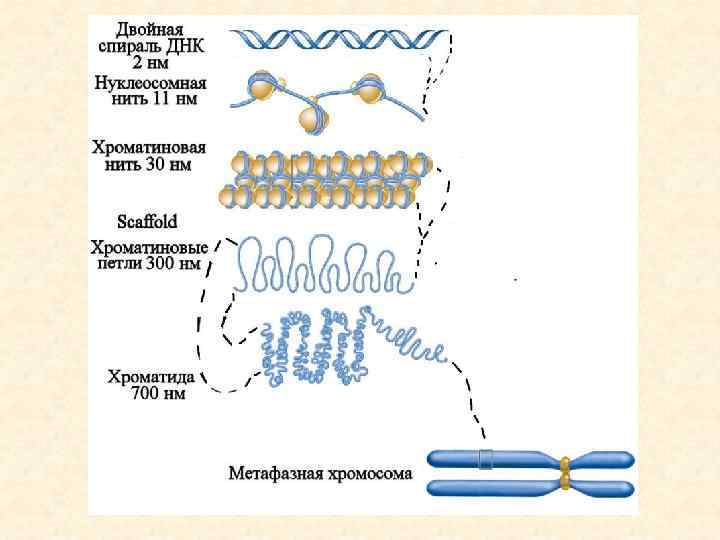
 II. Структурно-функциональная организация хромосом ТЕЛОМЕРА Белки кинетохора ЦЕНТРОМЕРА ТЕЛОМЕРА Ø Хромосома содержит длинную линейную молекулу ДНК Ø Гены локализованы между центромерным и теломерными районами (от нескольких сотен до нескольких тысяч различных генов) Ø Каждая хромосома содержит центромеру, в которой формируется сайт узнавания белков кинетохора Ø Теломера содержит специализированные последовательности, которые локализованы на обоих концах линейной хромосомы Ø Повторяющиеся последовательности в основном локализованы в центромерных и теломерных районах, но могут быть диспергированы в других районах хромосом Гены Повторяющиеся последовательности
II. Структурно-функциональная организация хромосом ТЕЛОМЕРА Белки кинетохора ЦЕНТРОМЕРА ТЕЛОМЕРА Ø Хромосома содержит длинную линейную молекулу ДНК Ø Гены локализованы между центромерным и теломерными районами (от нескольких сотен до нескольких тысяч различных генов) Ø Каждая хромосома содержит центромеру, в которой формируется сайт узнавания белков кинетохора Ø Теломера содержит специализированные последовательности, которые локализованы на обоих концах линейной хромосомы Ø Повторяющиеся последовательности в основном локализованы в центромерных и теломерных районах, но могут быть диспергированы в других районах хромосом Гены Повторяющиеся последовательности
 Три основных типа ДНК- последовательностей • Уникальная или неповторяющаяся • Умеренно повторяющаяся • Высоко повторяющаяся
Три основных типа ДНК- последовательностей • Уникальная или неповторяющаяся • Умеренно повторяющаяся • Высоко повторяющаяся
: § встречается один или несколько раз в") Ø Уникальная или неповторяющаяся (6070% ядерной ДНК): § встречается один или несколько раз в геноме, § включает структурные гены и внутригенные области ØУмеренно повторяющаяся: § повторенные сотни и тысячи раз в геноме; § включает: - гены р. РНК и гистонов -мобильные элементы (short interspersed elements (SINES), long interspersed elements (LINES)
Ø Уникальная или неповторяющаяся (6070% ядерной ДНК): § встречается один или несколько раз в геноме, § включает структурные гены и внутригенные области ØУмеренно повторяющаяся: § повторенные сотни и тысячи раз в геноме; § включает: - гены р. РНК и гистонов -мобильные элементы (short interspersed elements (SINES), long interspersed elements (LINES)
: § повторенные сотни тысяч или даже миллионы раз") ØВысоко повторяющаяся (25 -28% ядерной ДНК): § повторенные сотни тысяч или даже миллионы раз в геноме ; § каждая копия относительно короткая (от нескольких до нескольких сотен нуклеотидов в длину); § включает: -сателлитная ДНК структурного гетерохроматина (центромерные, теломерные районы хромосом) -Alu-повторы
ØВысоко повторяющаяся (25 -28% ядерной ДНК): § повторенные сотни тысяч или даже миллионы раз в геноме ; § каждая копия относительно короткая (от нескольких до нескольких сотен нуклеотидов в длину); § включает: -сателлитная ДНК структурного гетерохроматина (центромерные, теломерные районы хромосом) -Alu-повторы
 ЦЕНТРОМЕРНЫЙ РАЙОН ХРОМОСОМЫ Белки кинетохора ЦЕНТРОМЕРА Центромера - это структура, обеспечивающая удержание хромосом, правильность выстраивания хромосом в метафазной пластинке и их прикрепление к веретену; участок, ответственный за контроль наступления анафазы
ЦЕНТРОМЕРНЫЙ РАЙОН ХРОМОСОМЫ Белки кинетохора ЦЕНТРОМЕРА Центромера - это структура, обеспечивающая удержание хромосом, правильность выстраивания хромосом в метафазной пластинке и их прикрепление к веретену; участок, ответственный за контроль наступления анафазы
 ТЕЛОМЕРНЫЙ РАЙОН ХРОМОСОМЫ теломера ← Основной функцией теломер является сохранение целостности хромосомы и обеспечение ее полной репликации. теломера ←
ТЕЛОМЕРНЫЙ РАЙОН ХРОМОСОМЫ теломера ← Основной функцией теломер является сохранение целостности хромосомы и обеспечение ее полной репликации. теломера ←
n, заканчивающиеся 3’-однонитевым") СТРОЕНИЕ ТЕЛОМЕРЫ • Теломеры представляют собой в основном двунитевые некодирующие повторы (ТТАГГГ)n, заканчивающиеся 3’-однонитевым участком • Размер двунитевого участка варьирует от 3 до 20 т. п. н. , однонитевого – от 100 до 200 пар оснований «Теломеры состоят из сателлитных повторенных единиц TTAGGG и интенсивно окрашиваются методом Т-бэндинга» . (ISCN, 2009)
СТРОЕНИЕ ТЕЛОМЕРЫ • Теломеры представляют собой в основном двунитевые некодирующие повторы (ТТАГГГ)n, заканчивающиеся 3’-однонитевым участком • Размер двунитевого участка варьирует от 3 до 20 т. п. н. , однонитевого – от 100 до 200 пар оснований «Теломеры состоят из сателлитных повторенных единиц TTAGGG и интенсивно окрашиваются методом Т-бэндинга» . (ISCN, 2009)
 ФЕРМЕНТ ТЕЛОМЕРАЗА ОБЕСПЕЧИВАЕТ РЕПЛИКАЦИЮ ТЕЛОМЕРНОЙ ДНК Ø Резкое уменьшение активности теломеразы и укорочение теломер обычно сопутствует клеточной дифференцировке Ø Часто наблюдается зависимость размера теломеры от возраста человека (возраст-зависимое прогрессивное укорочение теломер) Феномен репликативного старения клеток, вызванного концевой недорепликацией ДНК, был предсказан А. Оловниковым в 1971 г. За биохимическое доказательство его идеи Э. Блэкберн (E. Blackburn), К. Грейдер (C. Greider) и Д. Шостак (J. Szostak) были удостоены в 2009 г. Нобелевской премии по физиологии и медицине. Ø Нарушение организации теломерных районов приводит к хромосомным перестройкам
ФЕРМЕНТ ТЕЛОМЕРАЗА ОБЕСПЕЧИВАЕТ РЕПЛИКАЦИЮ ТЕЛОМЕРНОЙ ДНК Ø Резкое уменьшение активности теломеразы и укорочение теломер обычно сопутствует клеточной дифференцировке Ø Часто наблюдается зависимость размера теломеры от возраста человека (возраст-зависимое прогрессивное укорочение теломер) Феномен репликативного старения клеток, вызванного концевой недорепликацией ДНК, был предсказан А. Оловниковым в 1971 г. За биохимическое доказательство его идеи Э. Блэкберн (E. Blackburn), К. Грейдер (C. Greider) и Д. Шостак (J. Szostak) были удостоены в 2009 г. Нобелевской премии по физиологии и медицине. Ø Нарушение организации теломерных районов приводит к хромосомным перестройкам
 локализованы в коротких плечах акроцентрических хромосом") ЯДРЫШКО-ОБРАЗУЮЩИЕ РАЙОНЫ ХРОМОСОМ • Ядрышко-образующие районы хромосом (ЯОР) локализованы в коротких плечах акроцентрических хромосом человека 13, 14, 15, 21 и 22. На препаратах метафазных хромосом активные ЯО-районы выявляются как вторичные перетяжки и окрашиваются азотнокислым серебром
ЯДРЫШКО-ОБРАЗУЮЩИЕ РАЙОНЫ ХРОМОСОМ • Ядрышко-образующие районы хромосом (ЯОР) локализованы в коротких плечах акроцентрических хромосом человека 13, 14, 15, 21 и 22. На препаратах метафазных хромосом активные ЯО-районы выявляются как вторичные перетяжки и окрашиваются азотнокислым серебром
") Окраска азотнокислым серебром (Ag-NOR)
Окраска азотнокислым серебром (Ag-NOR)
, кодирующие 18 S и 28 S рибосомную РНК") В ЯОР локализованы гены (р. ДНК), кодирующие 18 S и 28 S рибосомную РНК (р. РНК) – участие в транскрипции РНК. Функция ЯОР состоит в формировании сложной фибриллярной системы ядрышка, в которую поступают продукты активности генов р. РНК. В среднем на геном человека приходится около 400 -500 копий генов р. РНК, Однако их общее число в геноме разных индивидов может значительно варьировать (от 200 до 700).
В ЯОР локализованы гены (р. ДНК), кодирующие 18 S и 28 S рибосомную РНК (р. РНК) – участие в транскрипции РНК. Функция ЯОР состоит в формировании сложной фибриллярной системы ядрышка, в которую поступают продукты активности генов р. РНК. В среднем на геном человека приходится около 400 -500 копий генов р. РНК, Однако их общее число в геноме разных индивидов может значительно варьировать (от 200 до 700).
 46, XY 46, XX
46, XY 46, XX