хромосомыПрез.клетка .5.pptx
- Количество слайдов: 47
 • • Хромосомы 1. Строение метафазных хромосом 2. Строение и функция центромерного района 3. Строение и функция теломерного района 4. Типы метафазных хромосом 5. Политенные хромосомы 6. Хромосомы типа ламповых щеток
• • Хромосомы 1. Строение метафазных хромосом 2. Строение и функция центромерного района 3. Строение и функция теломерного района 4. Типы метафазных хромосом 5. Политенные хромосомы 6. Хромосомы типа ламповых щеток
:") Хромосомные наборы разных видов цветковых растений и животных (в скобках указано диплоидное число хромосом): Растения: А скерда зеленая (6), Б рожь (14), В мягкая пшеница (42), Г кувшинка гигантская (224), Д лютик малый (16), Е рябчик царский (30), Ж гаплоидный набор хромосом лука победного (8), З гаплоидный набор хромосом в пыльцевом зерне венечника ресничного (7), И две крупные хромосомы клубнекамыша приморского. Животные: А речной рак (98), Б комар (6), В щука (19), Г курица (78), Д кошка (38), справа метафаза первого мейоза с 33 мя парами хромосом, Е лошадь (66), справа метафаза мейоза с 33 мя парами хромосом, Ж бык (60), слева метафаза первого мейоза с 30 ю парами хромосом, З саламандра (34), И овца (54).
Хромосомные наборы разных видов цветковых растений и животных (в скобках указано диплоидное число хромосом): Растения: А скерда зеленая (6), Б рожь (14), В мягкая пшеница (42), Г кувшинка гигантская (224), Д лютик малый (16), Е рябчик царский (30), Ж гаплоидный набор хромосом лука победного (8), З гаплоидный набор хромосом в пыльцевом зерне венечника ресничного (7), И две крупные хромосомы клубнекамыша приморского. Животные: А речной рак (98), Б комар (6), В щука (19), Г курица (78), Д кошка (38), справа метафаза первого мейоза с 33 мя парами хромосом, Е лошадь (66), справа метафаза мейоза с 33 мя парами хромосом, Ж бык (60), слева метафаза первого мейоза с 30 ю парами хромосом, З саламандра (34), И овца (54).
 Фазы митоза: 1 интерфаза, 2 профаза, 3") • Морфология митотических хромосом. • А) Фазы митоза: 1 интерфаза, 2 профаза, 3 метафаза, 4 анафаза, 5, 6 телофаза • Б) Одна из метафазных хромосом. 1 Сестринские хромосомы (хроматиды); 2 плечи хромосом; 2 плечи хромосом; 3 центромера, первичная перетяжка; 4 вторичная перетяжка, ядрышковый организатор; 5 теломера; 6 пучки микротрубочек • В) Хромосома I человека при дифференциальной окраске: в сегментах (“бэндах”) • локализованы определенные
• Морфология митотических хромосом. • А) Фазы митоза: 1 интерфаза, 2 профаза, 3 метафаза, 4 анафаза, 5, 6 телофаза • Б) Одна из метафазных хромосом. 1 Сестринские хромосомы (хроматиды); 2 плечи хромосом; 2 плечи хромосом; 3 центромера, первичная перетяжка; 4 вторичная перетяжка, ядрышковый организатор; 5 теломера; 6 пучки микротрубочек • В) Хромосома I человека при дифференциальной окраске: в сегментах (“бэндах”) • локализованы определенные
 МЕТАФАЗНЫЕ ХРОМОСОМЫ
МЕТАФАЗНЫЕ ХРОМОСОМЫ
 Типы метафазных хромосом: 1, 6 метацентрические, 2 субметацентрические, 3, 4, 5 акроцентрические, 7 акроцентрическая со вторичной перетяжкой, 8 спутничная. Светлыми кружками обозначены центромеры.
Типы метафазных хромосом: 1, 6 метацентрические, 2 субметацентрические, 3, 4, 5 акроцентрические, 7 акроцентрическая со вторичной перетяжкой, 8 спутничная. Светлыми кружками обозначены центромеры.
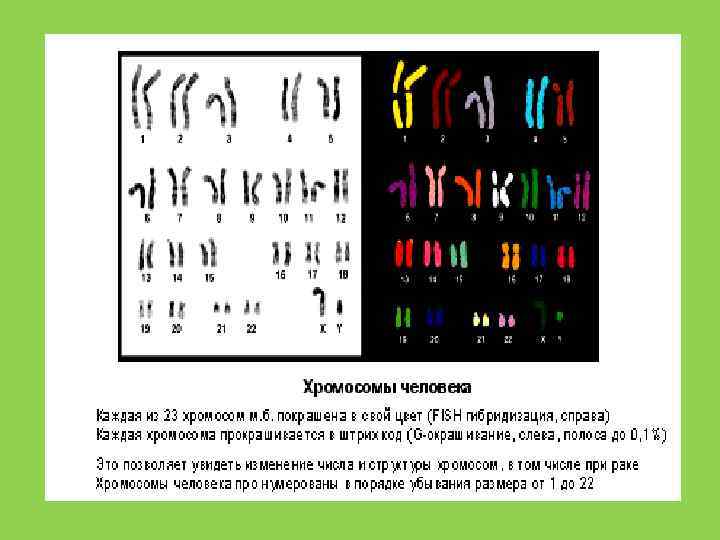
 Рубцов Н. Б. , Карамышева Т. В. Вестн. ВОГи. С, 2000).") Хромосомы человека(FISH –гибридизация) Рубцов Н. Б. , Карамышева Т. В. Вестн. ВОГи. С, 2000).
Хромосомы человека(FISH –гибридизация) Рубцов Н. Б. , Карамышева Т. В. Вестн. ВОГи. С, 2000).

 а деконденсированная центромера; б частично") Модель строения кинетохора (по: Zinkowski et al. , 1991) а деконденсированная центромера; б частично конденсированная центромера; в полностью конденсированная центромера. 1 участок ДНК с белковыми сегментами, связывающими микрогрубочки; 2 линкерный участок ДНК; 3 сформированный кинетохор; 4 микротрубочки; 5 плечи хромосом
Модель строения кинетохора (по: Zinkowski et al. , 1991) а деконденсированная центромера; б частично конденсированная центромера; в полностью конденсированная центромера. 1 участок ДНК с белковыми сегментами, связывающими микрогрубочки; 2 линкерный участок ДНК; 3 сформированный кинетохор; 4 микротрубочки; 5 плечи хромосом
 • . Схема локализация центромерных белков и ДНК • 1 хромосома; 2 центромера; 3 микротрубочки; 4 фиброзная корона: CENP E, динеины; 5 внешний слой: CENP E, CENP F; 6 промежуточная зона; 7 внутренний слой: CENP C, ДНК; 8 центромерный гетерохроматин: сателлитная ДНК, CENP B, МСАК, INCENP (когезин) • С
• . Схема локализация центромерных белков и ДНК • 1 хромосома; 2 центромера; 3 микротрубочки; 4 фиброзная корона: CENP E, динеины; 5 внешний слой: CENP E, CENP F; 6 промежуточная зона; 7 внутренний слой: CENP C, ДНК; 8 центромерный гетерохроматин: сателлитная ДНК, CENP B, МСАК, INCENP (когезин) • С
 Политенные хромосомы, выделенные из клеток слюнных желёз личинок дрозофилы ( Мюнтцинг. Генетические исследования, 1963. )
Политенные хромосомы, выделенные из клеток слюнных желёз личинок дрозофилы ( Мюнтцинг. Генетические исследования, 1963. )
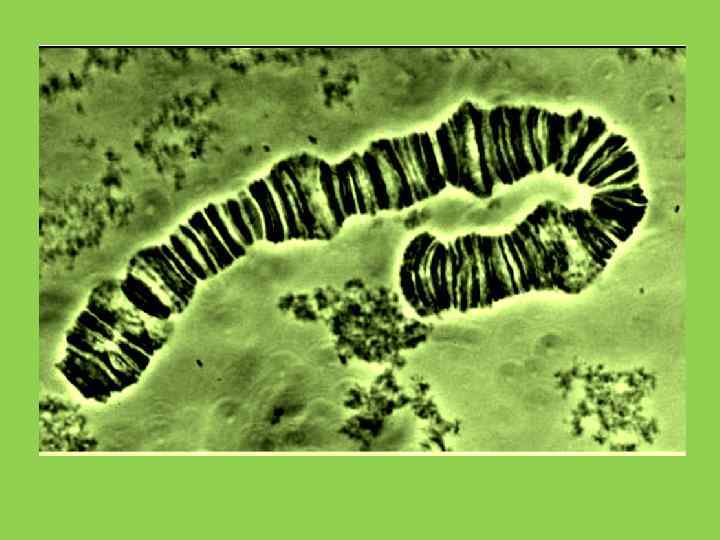
 • Участок политенной диски , имеющие псевдохромомерное строениехромосомы, содержащий диски разной морфологии • А «идеальный диск» ; В, С, Д ; • Е пуффирующий диск
• Участок политенной диски , имеющие псевдохромомерное строениехромосомы, содержащий диски разной морфологии • А «идеальный диск» ; В, С, Д ; • Е пуффирующий диск

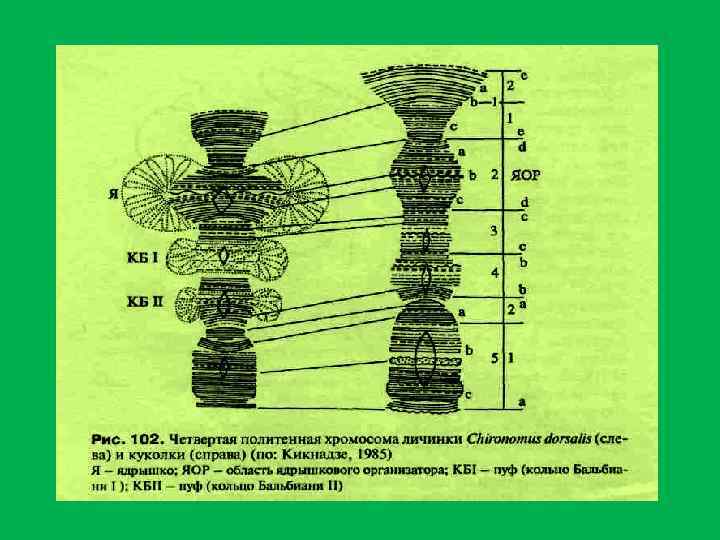
 Изменение рисунка участка хромосомы типа «ламповых щеток» при растяжении его с помощью микроманипулятора. [Из Callan, 1986. Р. 18]
Изменение рисунка участка хромосомы типа «ламповых щеток» при растяжении его с помощью микроманипулятора. [Из Callan, 1986. Р. 18]
. Схематическое") • Хромосомы типа ламповых щеток (Mancino et al. , 1969; Miller, 1973). Схематическое изображение структуры хромосом типа ламповых щеток в овогенезе саламандры [Salamandra salamandra]; L и R — левое и правое плечо хро мосом; —III — I номера хромосом • Хромосомы типа ламповых щеток впервые были обнаружены Рюккертом в 1892 г. •
• Хромосомы типа ламповых щеток (Mancino et al. , 1969; Miller, 1973). Схематическое изображение структуры хромосом типа ламповых щеток в овогенезе саламандры [Salamandra salamandra]; L и R — левое и правое плечо хро мосом; —III — I номера хромосом • Хромосомы типа ламповых щеток впервые были обнаружены Рюккертом в 1892 г. •
 • Ядрышко • 1. Число ядрышек в ядре, их хромосомное происхождение. ДНК ядрышка, строение генов р. РНК • 2. Структура ядрышка • 3. Фибрилярный центр и ядрышковый организатор • 4. Структурные типы ядрышек • 5. Цикл изменения структуры ядрышка в связи с его функциональной активностью • 6. Судьба ядрышка в течение клеточного цикла • 7. Строение и химический состав рибосом. • 8. Морфологическое строение большой и малой субъедициц рибосом. • 9. Функция рибосом. Синтез белка.
• Ядрышко • 1. Число ядрышек в ядре, их хромосомное происхождение. ДНК ядрышка, строение генов р. РНК • 2. Структура ядрышка • 3. Фибрилярный центр и ядрышковый организатор • 4. Структурные типы ядрышек • 5. Цикл изменения структуры ядрышка в связи с его функциональной активностью • 6. Судьба ядрышка в течение клеточного цикла • 7. Строение и химический состав рибосом. • 8. Морфологическое строение большой и малой субъедициц рибосом. • 9. Функция рибосом. Синтез белка.
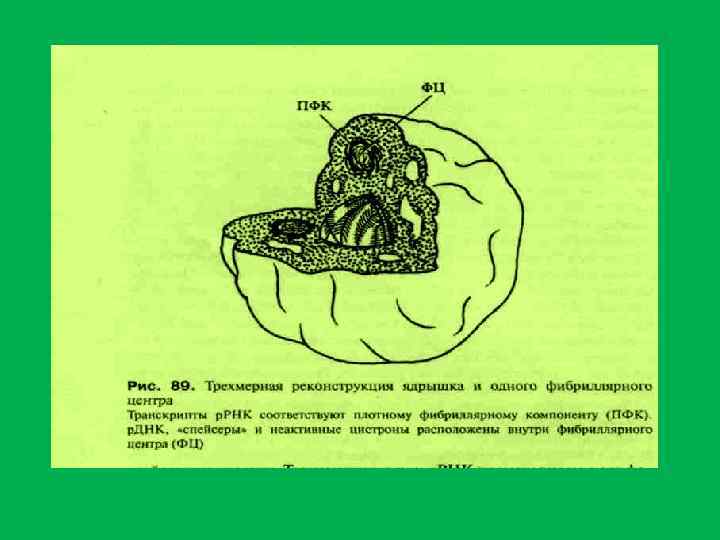
; 2 фибриллярные центры; 3 плотный фибриллярный компонент;") Схема компонентов ядрышка 1 гранулярный компонент (нуклеолонема); 2 фибриллярные центры; 3 плотный фибриллярный компонент; 4 околоядрышковый хроматин
Схема компонентов ядрышка 1 гранулярный компонент (нуклеолонема); 2 фибриллярные центры; 3 плотный фибриллярный компонент; 4 околоядрышковый хроматин
 Рис. Структурные типы ядрышек 1 ретикулярный; 2 компактный; 3 вакуолярный; 4 кольцевидный; 5 сегрегированный
Рис. Структурные типы ядрышек 1 ретикулярный; 2 компактный; 3 вакуолярный; 4 кольцевидный; 5 сегрегированный
 • Схема активации и инактивации ядрышкового организатора • 1 начало активации «елочки» появляются по периферии одного крупного ФЦ (ЯОР хромосомы); 2, 3 ~ дальнейшая активация приводит к образованию множественных ФЦ; 4 полная активация; 5 полная инактивация
• Схема активации и инактивации ядрышкового организатора • 1 начало активации «елочки» появляются по периферии одного крупного ФЦ (ЯОР хромосомы); 2, 3 ~ дальнейшая активация приводит к образованию множественных ФЦ; 4 полная активация; 5 полная инактивация
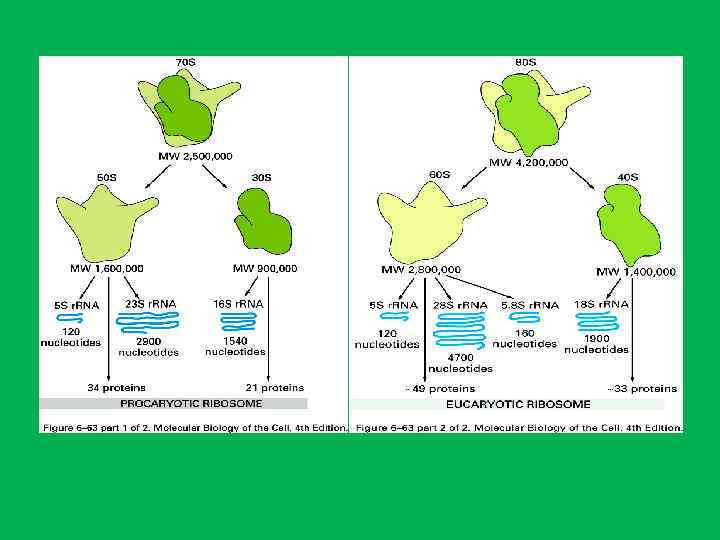
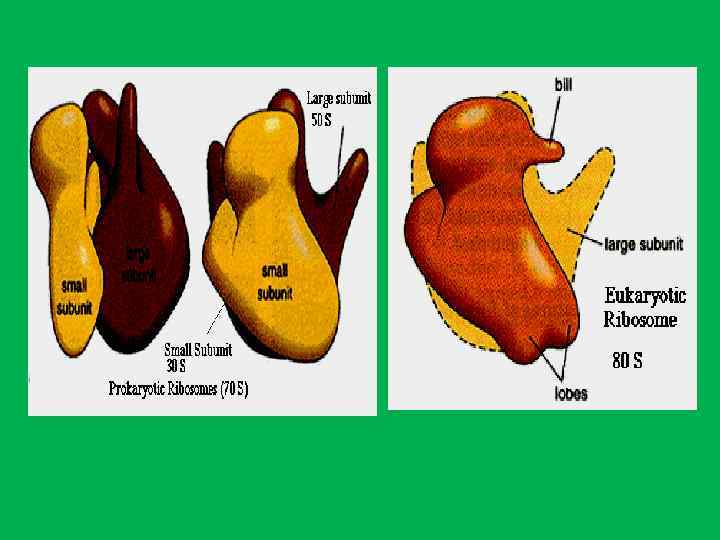
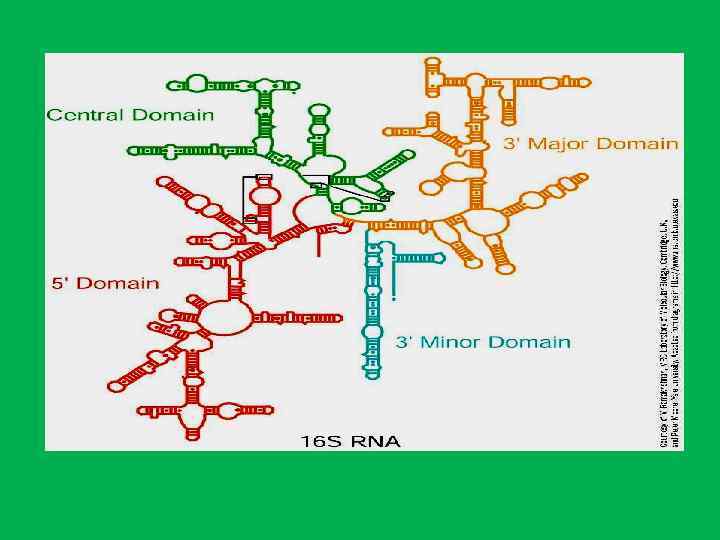
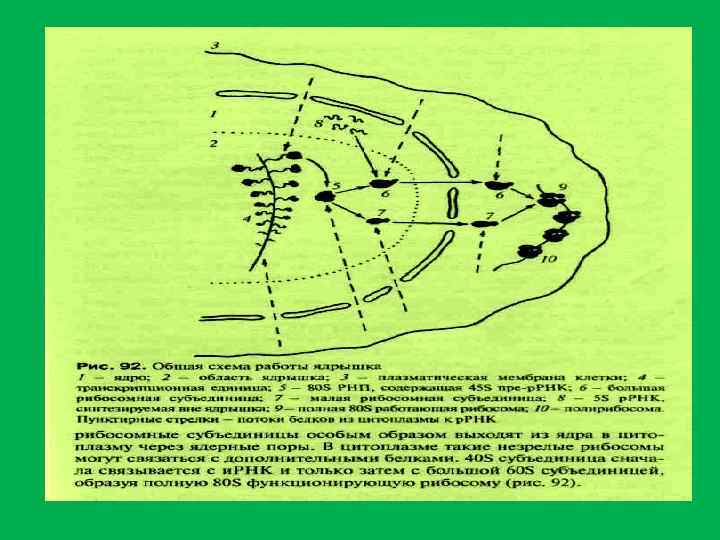
 • Белки ядрышка составляют 60% от сухой массы • 1. Белки рибосом • 2. белки , связанные с транскрипцией р. РНК (РНК полимераза I, факторы транскрипции топоизомеразы, метилазы, нуклеазы, протеинкиназы, фосфатазы ) • РНК полимераза и факторы транскрипции р. РНК располагаются в фибрилярных центрах (ФЦ), а также в плотном фибриллярном компоненте (ПФК). • Специфический для ядрышка белок фибрилларин(34 к. Да) , располагается в плотном фибриллярном компоненте, где он осуществляет процессинг пре р. РНК. И остаточном ядрышковом матриксе. • Белок С 23 (110 к. Да)или нуклеолин локализуется в области ПФК и в ФЦ и в зонах ядрышковых организаторов митотических хромосом • Белок В 23 (нуклеофозин, 37 к. Да) – участвует в прмежуточных и терминальных стадиях синтеза рибосом и в транспорте пре рибосом.
• Белки ядрышка составляют 60% от сухой массы • 1. Белки рибосом • 2. белки , связанные с транскрипцией р. РНК (РНК полимераза I, факторы транскрипции топоизомеразы, метилазы, нуклеазы, протеинкиназы, фосфатазы ) • РНК полимераза и факторы транскрипции р. РНК располагаются в фибрилярных центрах (ФЦ), а также в плотном фибриллярном компоненте (ПФК). • Специфический для ядрышка белок фибрилларин(34 к. Да) , располагается в плотном фибриллярном компоненте, где он осуществляет процессинг пре р. РНК. И остаточном ядрышковом матриксе. • Белок С 23 (110 к. Да)или нуклеолин локализуется в области ПФК и в ФЦ и в зонах ядрышковых организаторов митотических хромосом • Белок В 23 (нуклеофозин, 37 к. Да) – участвует в прмежуточных и терминальных стадиях синтеза рибосом и в транспорте пре рибосом.
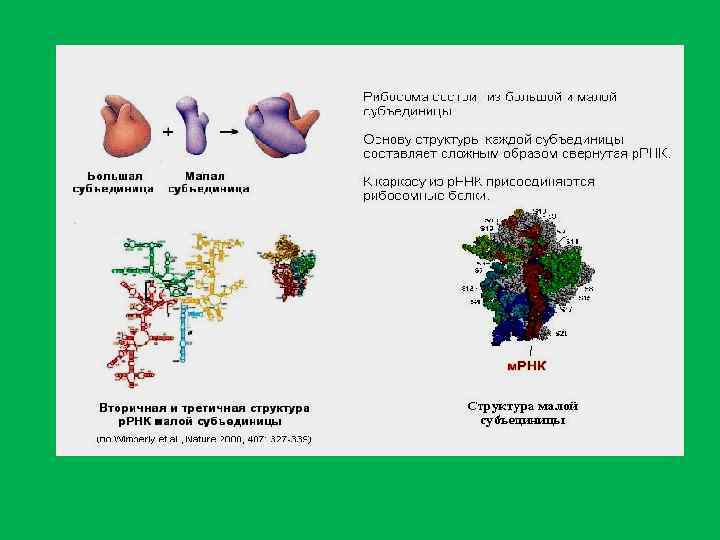
 Сравнение контуров рибосомных субчас тиц и их изолированных высокополимерных РНК в компактной форме по данным электронной мик роскопии: вверху большая S) субчастица и ее 23 S РНК. Внизу малая (30 S) субчастица и ее 16 S РНК (В. Д. Васильев, Институт белка РАН, Пущино
Сравнение контуров рибосомных субчас тиц и их изолированных высокополимерных РНК в компактной форме по данным электронной мик роскопии: вверху большая S) субчастица и ее 23 S РНК. Внизу малая (30 S) субчастица и ее 16 S РНК (В. Д. Васильев, Институт белка РАН, Пущино
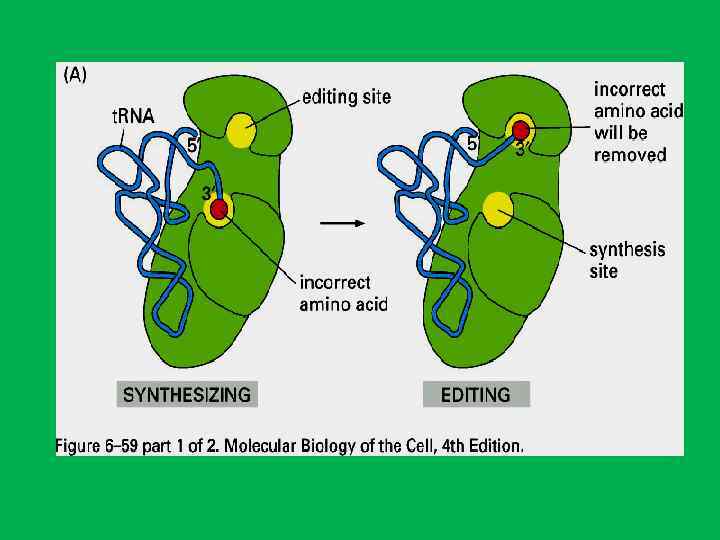
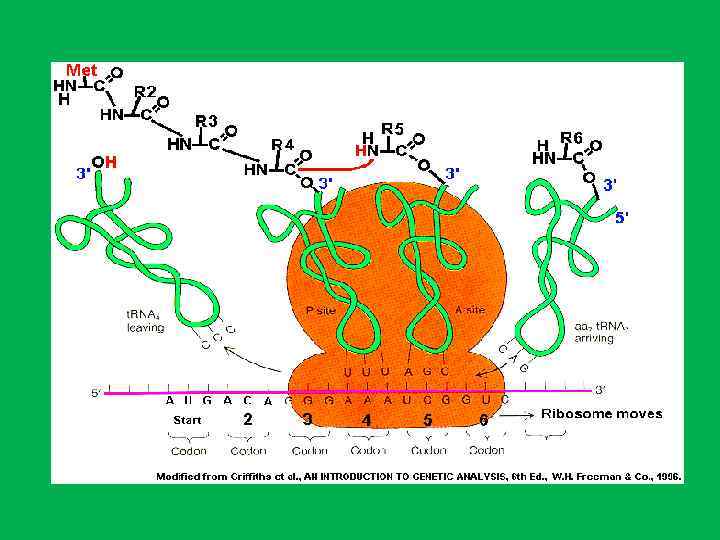
 и двух т. РНК") Размещение основных функциональных лигандов цепи м. РНК (обозначена синим цветом) и двух т. РНК (зеленые) в рибосоме. Контуры рибосомы даны в соответствии с последними данными криоэлектронной микроскопии. Полость между субчастицами является главным функциональным карманом рибосомы, здесь размещаются две молекулы т. РНК. Молекулы т. РНК (эминоацил т. РНК и пептидил т. РНК) связаны с м. РНК своими антикодоновыми верхушками и с пептидил трансферазным центром в основании центрального выступа большой субчастицы своими акцепторными концами, несущими аминокислотные остатки. В процессе трансляции цепь м. РНК сканируется рибосомой от 5' конца (голова цепи) к З' концу (хвост цепи), и т. РНК сменяются в зависимости от нуклеотидных комбинаций, находящихся в каждый данный момент на рибосоме. Согласно представленной модели, цепь м. РНК движется сквозь рибосому через шею малой субчастицы и выходит в зазор между центральным и левым боковыми выступами большой субчастицы
Размещение основных функциональных лигандов цепи м. РНК (обозначена синим цветом) и двух т. РНК (зеленые) в рибосоме. Контуры рибосомы даны в соответствии с последними данными криоэлектронной микроскопии. Полость между субчастицами является главным функциональным карманом рибосомы, здесь размещаются две молекулы т. РНК. Молекулы т. РНК (эминоацил т. РНК и пептидил т. РНК) связаны с м. РНК своими антикодоновыми верхушками и с пептидил трансферазным центром в основании центрального выступа большой субчастицы своими акцепторными концами, несущими аминокислотные остатки. В процессе трансляции цепь м. РНК сканируется рибосомой от 5' конца (голова цепи) к З' концу (хвост цепи), и т. РНК сменяются в зависимости от нуклеотидных комбинаций, находящихся в каждый данный момент на рибосоме. Согласно представленной модели, цепь м. РНК движется сквозь рибосому через шею малой субчастицы и выходит в зазор между центральным и левым боковыми выступами большой субчастицы
 и большой (внизу) субчастицах рибосомы. Цепь м. РНК") Расположение функциональных центров на малой (вверху) и большой (внизу) субчастицах рибосомы. Цепь м. РНК связана с малой субчастицей в районе ее "шеи" и протягивается через этот м. РНК связывающий центр в ходе трансляции от 5' конца (направлен вниз и вправо) к З' концу (обращен вверх и влево). Две молекулы г. РНК занимают А- и Р участки на малой субчастице, будучи связаны своими антикодонами с двумя смежными кодонами м. РНК в районе "шеи" малой субчастицы. Пептидилтрансферазный центр (РТС) распо ложен в районе "шеи" (в борозде под центральным выступом) большой субчастицы. Когда субчастицы ассоциированы, акцепторные концы двух т. РИК с их аминоацильным и пептидильным остатками взаимодействуют с пептид и лтрансфераз ным центром большой субчастицы. Факторы элонгации EFJ и EF 2 связываются в районе палочкообразного бокового выступа большой субчастицы (на рисунке направлен на зрителя) со стороны, обращенной к малой субчастице
Расположение функциональных центров на малой (вверху) и большой (внизу) субчастицах рибосомы. Цепь м. РНК связана с малой субчастицей в районе ее "шеи" и протягивается через этот м. РНК связывающий центр в ходе трансляции от 5' конца (направлен вниз и вправо) к З' концу (обращен вверх и влево). Две молекулы г. РНК занимают А- и Р участки на малой субчастице, будучи связаны своими антикодонами с двумя смежными кодонами м. РНК в районе "шеи" малой субчастицы. Пептидилтрансферазный центр (РТС) распо ложен в районе "шеи" (в борозде под центральным выступом) большой субчастицы. Когда субчастицы ассоциированы, акцепторные концы двух т. РИК с их аминоацильным и пептидильным остатками взаимодействуют с пептид и лтрансфераз ным центром большой субчастицы. Факторы элонгации EFJ и EF 2 связываются в районе палочкообразного бокового выступа большой субчастицы (на рисунке направлен на зрителя) со стороны, обращенной к малой субчастице
 ПОЛИСОМА
ПОЛИСОМА
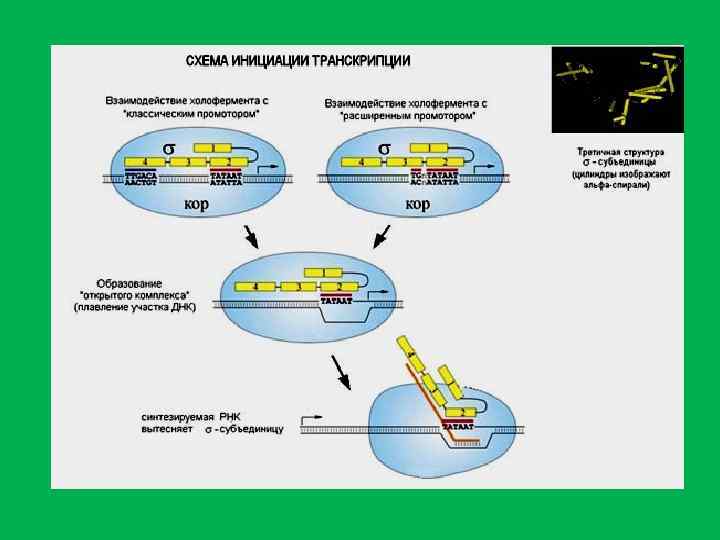
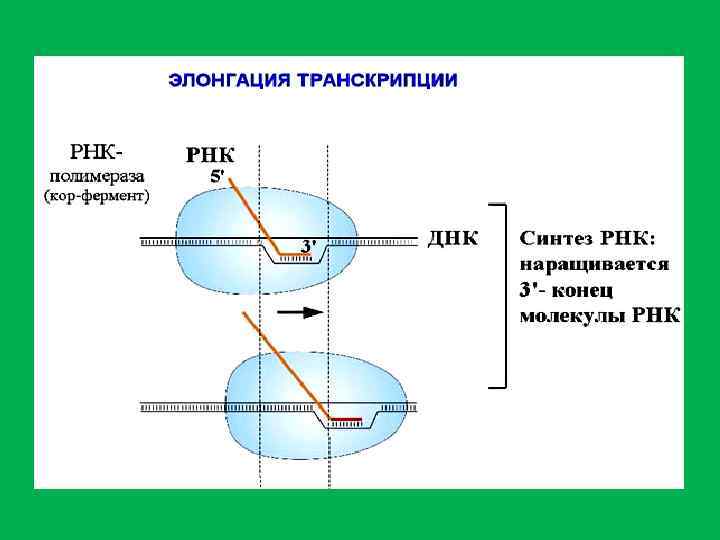
; элетронная микрофотография транскрипции (по V. Foe).") Две возможные модели транскирипции (А и Б); элетронная микрофотография транскрипции (по V. Foe).
Две возможные модели транскирипции (А и Б); элетронная микрофотография транскрипции (по V. Foe).
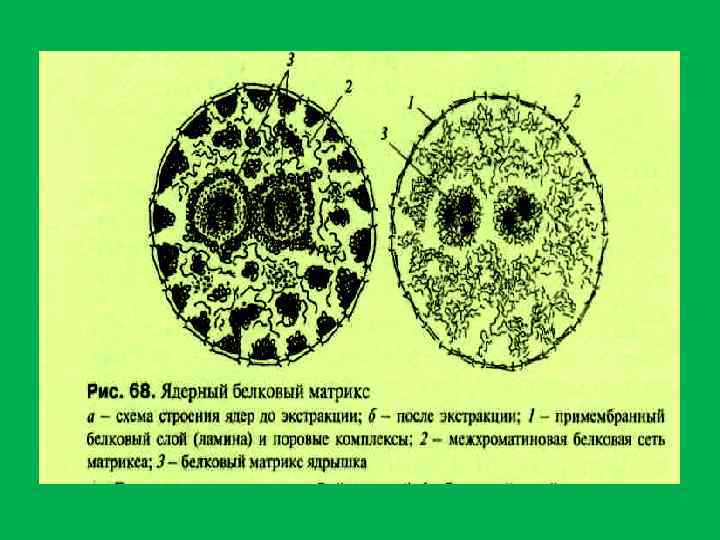
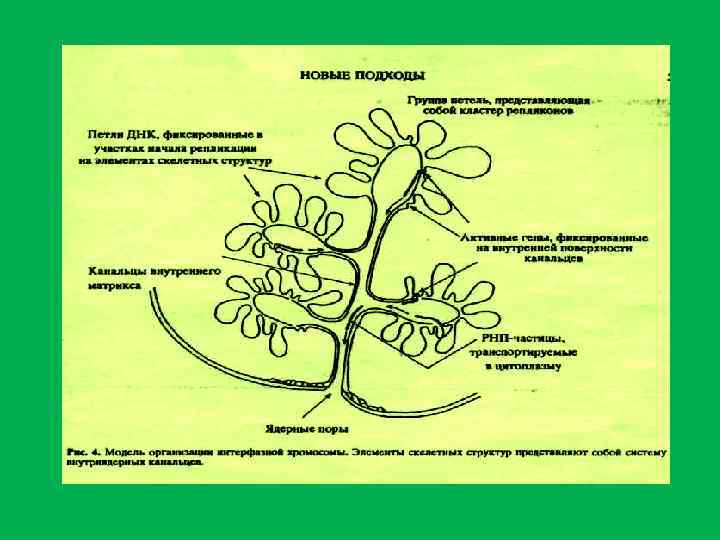
 • Ядерный матрикс состоит из трех компонентов: • 1. Периферического белкового сетчатого слоя ламины, 2. внутренней , или интерхроматиновой, сети (остов), 3. остаточного ядрышка • . Ламина – это тонкий фиброзный слой, расположенный под внутренней мембраной ядерной оболочки. В ее состав Входят также компексы ядерных пор , которые погружены в фиброзный слой. Ламина поддерживает морфологическую целостность ядра . • 2. После экстракции хроматина в ядре выявляется внутриядерный остов , или сеть. Он представлен рыхлой фиброзной сетью, которая располагается между хроматином • 3. «остаточное» ядрышко – плотная структура , повторяющая по своей форме ядрышко, также состоит из плотно уложенных фибрилл. • Основным компонентом ядерного матрикса является белок(88 98%%): • а). ламины А, В, С. • в). в состав внутриядерного остова входит ряд белков. Принимающих участие в доменной организации ДНК в интерфазном ядре при образовании хромомерной формы упаковки хроматина.
• Ядерный матрикс состоит из трех компонентов: • 1. Периферического белкового сетчатого слоя ламины, 2. внутренней , или интерхроматиновой, сети (остов), 3. остаточного ядрышка • . Ламина – это тонкий фиброзный слой, расположенный под внутренней мембраной ядерной оболочки. В ее состав Входят также компексы ядерных пор , которые погружены в фиброзный слой. Ламина поддерживает морфологическую целостность ядра . • 2. После экстракции хроматина в ядре выявляется внутриядерный остов , или сеть. Он представлен рыхлой фиброзной сетью, которая располагается между хроматином • 3. «остаточное» ядрышко – плотная структура , повторяющая по своей форме ядрышко, также состоит из плотно уложенных фибрилл. • Основным компонентом ядерного матрикса является белок(88 98%%): • а). ламины А, В, С. • в). в состав внутриядерного остова входит ряд белков. Принимающих участие в доменной организации ДНК в интерфазном ядре при образовании хромомерной формы упаковки хроматина.
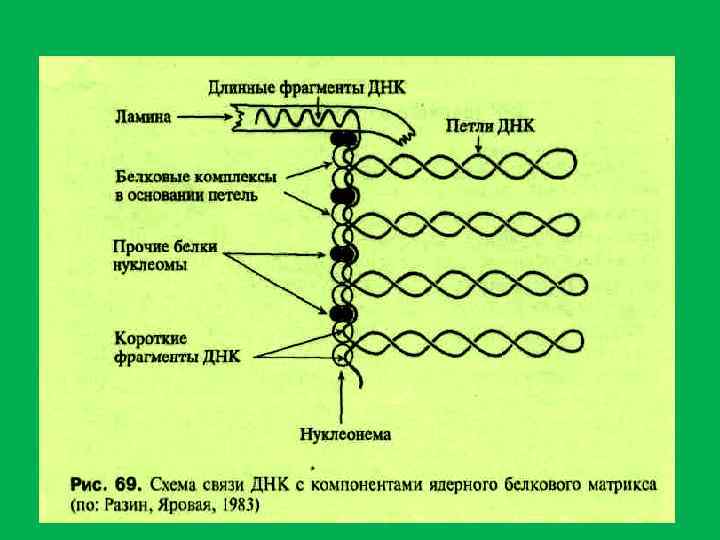
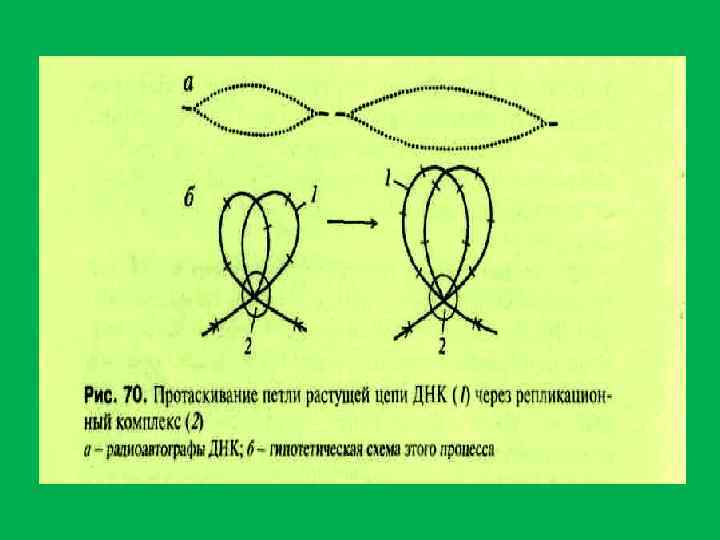
. В") • ДНК ядерного белкового матрикса (составляет менее 1% от всей днк ядра). В ядрах существует от 60 000 до 125 000 участков ДНК, защищенных от действия нуклеаз. • 1. группа. высокомолекулярные фрагменты размером около 10 т. пн. (0, 02%) • Эти фрагменты обогащены сателитной ДНК и связаны с ламиной. 2 группа – представлена небольшими фрагментами ДНК(120 140 н. п. ). Они располагаются между участками ДНК длиной около 50 т. п. н. (вероятно петли основной массы хроматина). Зоны связывания ДНК с матриксом (MAR или. SAR)содержат примерно 200 п. н. и располагаются друг от друга на растоянии 5 112 т. п. н. Ядерный матрикс связан с репликацией ДНК. В ядерном матриксе обнаружена ДНК полимераза, ДНК праймаза, ДНК лигаза, ДНК топоизомераза. II. В состав ядерного матрикса входит около 1% РНК(м. РНК. р. РНК. т. РНК, мя. РНК) На ядерном матриксе закреплены транскрипционные комплексы и при транскрипции м. ДНК как бы пртаскивается через эти комплексы, содержащие РНК полимеразу II.
• ДНК ядерного белкового матрикса (составляет менее 1% от всей днк ядра). В ядрах существует от 60 000 до 125 000 участков ДНК, защищенных от действия нуклеаз. • 1. группа. высокомолекулярные фрагменты размером около 10 т. пн. (0, 02%) • Эти фрагменты обогащены сателитной ДНК и связаны с ламиной. 2 группа – представлена небольшими фрагментами ДНК(120 140 н. п. ). Они располагаются между участками ДНК длиной около 50 т. п. н. (вероятно петли основной массы хроматина). Зоны связывания ДНК с матриксом (MAR или. SAR)содержат примерно 200 п. н. и располагаются друг от друга на растоянии 5 112 т. п. н. Ядерный матрикс связан с репликацией ДНК. В ядерном матриксе обнаружена ДНК полимераза, ДНК праймаза, ДНК лигаза, ДНК топоизомераза. II. В состав ядерного матрикса входит около 1% РНК(м. РНК. р. РНК. т. РНК, мя. РНК) На ядерном матриксе закреплены транскрипционные комплексы и при транскрипции м. ДНК как бы пртаскивается через эти комплексы, содержащие РНК полимеразу II.