
Хромосомная теория наследственности. Метод цитогенетики. План Занятия: 1.







 и американского цитолога")














 Соматический кроссинговер впервые был выявлен у дрозофилы")



 сцеплении генов 26 Р: AaBb")



:")








 – комплекс ДНК с белками (гистоновыми и негистоновыми) Строение")



, образующий несколько сортов гамет и")

 arm long (q-) arm centromere chromatid telomere telomere")

,")


 –Н 1, Н2А, Н2В, Н3, Н4")

 Вид")

, соединенных друг с другом в")
 — участок ДНК, в районе который")



- локализован в прицентромерных и теломерных районах. Это постоянно выявляемые участки, состоит")
 Хроматин Эухроматин (слабо конденсированный,")


")

 Гистон")
![Расположение молекул гистонов в нуклеосоме [Lewin, 2000. Р. 582]](https://present5.com/presentacii-2/20171208\14410-topic_4.ppt\14410-topic_4_69.jpg "Расположение молекул гистонов в нуклеосоме [Lewin, 2000. Р. 582]")










 Гистон Н1 связывается с ДНК между корами. Компактизация – 40 раз.")

 Нуклеосомы (11 нм) 30 нм Укладка типа «соленоида» 300 нм")




 и его студент Бертрам открыли в 1948 году")





 пятна.")



 раздел генетики, изучающий закономерности наследственности и изменчивости на уровне клетки и субклеточных")



. Фитогемагглютинин является углеводом обыкновенной")


, механический центр хромосомы,")

 Отделение лейкоцитов Добавление стимулятора митоза – ФГА (фитогемагглютинина) 72 часа")

")
 Субметацентрические (Sm) Субакроцентрические (Sа) Акроцентрические (А) Телоцентрические (Т)")
 используется при классификации хромосом. Центромера")
![Типы метафазных хромосом [МакКьюсик, 1967. С. 25]](https://present5.com/presentacii-2/20171208\14410-topic_4.ppt\14410-topic_4_112.jpg "Типы метафазных хромосом [МакКьюсик, 1967. С. 25]")






. Нерасхождение")
 Парижская (дифференциальная окраска) Q, G, R, C и др.")


: крупные хромосомы, отличимые друг от друга; центромеры находятся")
: хромосомы средних размеров, трудно отличимые друг от друга; центромеры")
: короткие хромосомы. Группа F (19-20): мелкие, центромеры посередине. Группа")






 Treated with temperature Giemsa stain is used)")

 leucocytes chromosomes")
 PI (propidium iodide) Quinacrine Ethidium bromide")


 и суббенды")





 кольцо ацентрические фрагменты")





![Гены болезней, картированные в первой хромосоме [McKusick, 1992. - Из:В.П.Пузырев,1996.С. 25 ].](https://present5.com/presentacii-2/20171208\14410-topic_4.ppt\14410-topic_4_152.jpg "Гены болезней, картированные в первой хромосоме [McKusick, 1992. - Из:В.П.Пузырев,1996.С. 25 ].")



 Анализ родословных (генеалогический) Генетики")





 политенные хромосомы (в слюнных железах")



 хромосомы Репликация ДНК не сопровождается делением клетки, что приводит к накоплению вновь")



14410-topic_4.ppt
- Количество слайдов: 170
Хромосомная теория наследственности. Метод цитогенетики.
План Занятия: 1. Хромосомная теория наследственности. 2. Типы хромосом. 3. Организация хроматина 4. Половой хроматин. 5. Понятие кариотипа и идиограммы.. 6.Методы изучения кариотипа человека. 3. Денверская и Парижская классификации хромосом человека.
Термин «хромосома» предложил в 1888 г. немецкий морфолог Вильгельм Вальдейер для обозначения внутриядерных структур эукариотической клетки, окрашиваемых основными красителями (от греч. хрома — цвет, краска, и сома — тело) 3
Роль хромосом в передаче наследственной информации была доказана благодаря: Определению групп сцепления признаков, которые соответствовали числу хромосом Построению генетических, а потом и цитологических карт хромосом Открытию генетического определения пола 4
Соответствие законов Менделя поведению хромосом Г. Мендель считал, что признаки передаются из поколения в поколение как отдельные «факторы», расположенные в половых клетках Г. Мендель не знал места расположения этих факторов в клетках, так как в то время не было известно о существовании хромосом неизвестной была роль ядра в процессе размножения не были открыты процессы митоза и мейоза 5
К началу XX ст. углубленное изучение поведения хромосом в ходе деления клетки, при созревании половых клеток, при оплодотворении и раннем развитии зародыша выявило строго закономерные изменения их организации 6
Это привело немецкого цитолога и эмбриолога Теодора Бовери (1902—1907) и американского цитолога Уолтера Сэттона (1902—1903) к установлению связи наследственного материала с хромосомами, что легло в основу хромосомной теории наследственности 7
В частности, они отметили: хромосомы существуют как гомологичные пары (Мендель считал, что наследственные факторы существуют в парах) гомологичные хромосомы распределяются при мейозе таким образом, что гаметы получают только одну хромосому из пары (Мендель считал, что при образовании гамет в каждую гамету попадает только один фактор из пары) 8
хромосомы различных гомологичных пар группируются случайным образом при мейозе и распределяются по гаметам независимо от любой другой пары (Мендель считал, что факторы каждой пары являются независимыми от любой другой пары при их распределении по гаметам) 9
10 - гомологичные хромосомы от двоих родителей встречаются в результате слияния мужской и женской гамет, при этом восстанавливается диплоидное число у потомков (Мендель считал, что мужские и материнские факторы смешиваются во время оплодотворения)
В 1902 г. Бовери и Сэттон сформулировали основные положения хромосомной теории наследственности – учения о локализации наследственных факторов в хромосомах: гены расположены в хромосомах в каждой гомологичной хромосоме находится по одному аллельному гену гомологичные хромосомы и вместе с ними аллельные гены объединяются в зиготе во время оплодотворения 11
Молодая наука генетика попала в кризисную ситуацию! Сэттон в 1903 г., пытаясь объяснить противоречие, допустил, что каждая хромосома содержит больше, чем 1 ген, и все гены, расположенные в одной хромосоме, наследуются вместе Но он не смог подтвердить это экспериментально! 12
Все эти протиречия легли в основу нового открытия, сделанного профессором экспериментальной зоологии Колумбийского университета в Нью-Йорке – Томасом Хантом Морганом Он сумел объединить данные статистики и результаты исследования процессов, происходящих в клетках 13 Томас Хант Морган (1866-1945)
Морган начал генетические эксперименты в 1909 г. Объектом исследований была избрана плодовая мушка - Drosophila melanogaster очень плодовита (25 поколений в год ) от яйца до взрослой особи - 10 дней мало хромосом (4 пары) имеет много признаков (цвет глаз, форма крыльев, окраска тела) 14
Некоторые мутации
Морган выявил то же самое «взаимное притяжение генов», которое наблюдали в 1906 г. Бэтсон и Пеннет Было установлено, что наследственные признаки дрозофилы можно разделить на три связанные между собой группы, которые наследуются как единое целое Морган назвал этот феномен сцеплением генов 16
Как цитолог он знал о существовании в клетках дрозофилы трех больших хромосом Существует и четвертая, небольшая хромосома В 1914 г. Герман Джозеф Мёллер, работавший у Моргана, открыл четвертую группу генов (очень малочисленную) 17
Таким образом Морган установил, что гены действительно находятся в хромосомах. Гены, принадлежащие к одной группе, в следующих поколениях неожиданно оказывались в различных группах 18 Морган сделал предположение, что происходит обмен генетическим материалом между различными хромосомами. Ему даже удалось наблюдать этот процесс под микроскопом: две хромосомы сближались и перекрещивались, обмениваясь фрагментами. Этот процесс получил название кроссинговера
Кроссинговер Если сцепление является неполным, то в профазе мейоза I в стадии пахитены может происходить обмен идентичными участками гомологичных хромосом – кроссинговер (перекрест) Это явление изучалось Т. Морганом на дрозофилах Кроссинговер – это обмен гомологичными участками гомологичных хромосом, который может сопровождаться обменом аллельных генов 19
Кроссинговер может происходить в одном или в нескольких участках хромосом Кроссинговер – генетически обусловленный процесс. У большинства организмов кроссинговер характерен как для самок, так и для самцов, но у некоторых (например, дрозофилы) он происходит только у гомогаметного пола (самок) 20
Кроссинговер
Типы кроссинговера Различают кроссинговер: мейотический митотический (соматический) Соматический кроссинговер впервые был выявлен у дрозофилы Куртом Стерном. Происходит со значительно меньшей частотой, чем при мейозе. Генетического значения не имеет. Индивидуумы с митотическим кроссинговером - мозаики 22
В зависимости от количества появившихся хиазм: одиночный кроссинговер – образуется только одна хиазма, что приводит к обмену только одним участком ДНК гомологичных хромосом. Это наиболее распространенный тип кроссинговера двойной кроссинговер - образуются две хиазмы. Этот тип кроссинговера приводит к обмену двумя участками ДНК гомологичных хромосом множественный кроссинговер - образуется более, чем две хиазмы между несестринскими хроматидами гомологичных хромосом. Далее они могут быть классифицированы как тройные (3 хиазмы), четвертные (4 хиазмы) и т.д. 23
24 А a B b Гены Несцепленные Сцепленные (в разных хромосомах) (в одной хромосоме) Полное (абсолютное) Неполное сцепление сцепление А a B b А a B b 30 000 генов 46 хромосом
Наследование несцепленных генов 25 Р: AaBb x aabb G: F1: AaBb, Aabb, aaBb, aabb А a B b a a b b Анализирующее скрещивание 25% 25% 25% 25% 100% 25% 25% 25% 25%
Наследование при полном (абсолютном) сцеплении генов 26 Р: AaBb x aabb G: F1: AaBb, aabb А a B b a a b b Анализирующее скрещивание 50% 50% 100% 50% 50% B b А a
Наследование при неполном сцеплении генов 27 Р: AaBb x aabb G: А a B b a a b b Анализирующее скрещивание 100% A A a a B B b b A B a b A a b B A a b B некр некр 10% кросс10% 40% 40%
F1: AaBb, Aabb, aaBb, aabb 40% 10% 10% 40% некр. кросс. кросс. некр. Некроссоверные >>> Кроссоверные гаметы гаметы 28
Определение частоты кроссинговера Установлено, что вероятность кроссинговера между определенными сцепленными генами зависит от расстояния между ними в хромосоме Т. Морган установил, что чем дальше расположены гены в хромосоме, тем более вероятным является кроссинговер Для близко расположенных генов кроссинговер менее вероятен Частота кроссинговера выражается в процентах кроссоверных (рекомбинантных) генотипов, полученных после анализирующего скрещивания 29
Частота кроссинговера (расстояние между генами): число кроссоверных организмов ЧК = * 100% общее число потомков 30
Эта частота строго пропорциональна расстоянию между сцепленными генами и измеряется в морганидах 1 морганида соответствует 1% рекомбинантных гамет или генотипов, полученных при анализирующем скрещивании 31
F1: AaBb, Aabb, aaBb, aabb 80 20 20 80 некр. кросс. кросс. некр. Расстояние между генами = 20+20 * 100% = 20% = 20 морганид 80+20+20+80 32
Митотическая активность ткани – это отношение числа клеток, находящихся в митозе, к общему числу клеток исследуемой ткани, которое выражается либо в процентах (%), либо в промилле (‰), т. е. в числе митозов на 1000 клеток. Показатель митотической активности называется митотическим индексом М I =M/N ×1000 MI (‰), где М – число клеток в митозе, N – число клеток в зоне деления.
Факторы, влияющие на частоту кроссинговера Температура – высокая и низкая температура повышает частоту Рентгеновские лучи – повышают Возраст – снижает Мутагены – повышают или снижают Пол – у самцов дрозофилы кроссинговер не происходит, у самцов млекопитающих интенсивность кроссинговера незначительна Расстояние между генами Пищевые эффекты – Са и Mg могут повышать или снижать Центромерные эффекты – гены в области центромеры менее способны к кроссинговеру 34
Значение кроссинговера Кроссинговер — широко распространенное явление Происходит практически у всех организмов, которые размножаются половым путем Этот процесс является молекулярной основой комбинативной изменчивости В результате рекомбинации генов могут появляться новые полезные признаки и их сочетания. Поэтому кроссинговер имеет большое значение для выживания и размножения Этот процесс также увеличивает генетическое разнообразие потомков, что очень важно для приспособления и эволюции Определение частоты кроссинговера лежит в основе картирования генов хромосом, то есть определения места расположения разных генов в хромосоме 35
Хромосомная теория сцепления Моргана и Кастла Гены, проявляющие сцепление, расположены в одной и той же хромосоме Гены расположены в линейной последовательности в хромосоме, то есть сцепление генов – линейное Расстояние между сцепленными генами обратно пропорционально силе сцепления Сцепленные гены остаются в своей комбинации во время наследования Гены, локализованные в одной хромосоме, передаются вместе (сцепленно) и составляют одну группу сцепления Количество групп сцепления равно гаплоидному числу хромосом Так, у мухи дрозофилы 8 хромосом — 4 группы сцепления, у каждого человека 46 хромосом — 23 группы сцепления (но теоретически 24 группы сцепления) 36
ДНК бактерий тоже иногда называется бактериальной хромосомой. Она кольцевая и лишена гистонов.
Хромосома бактерий имеет небольшое количество белков Оболочка клетки Точка начала репликации ДНК ДНК ДНК-связывающие белки ДНК в виде двойной спирали 20% белков Роль белков- компактизация и прикрепление ДНК к оболочке бактерии
Хромосома (греч. – «окрашенное тело») – комплекс ДНК с белками (гистоновыми и негистоновыми) Строение хромосом эукариот ДНК гистоны хромосома
Хромосомы – структура ДНК и белка, которая находится в ядре клетки. Клетки человека содержат 46 хромосом в 23 парах – одна из каждой пары получена от одного родителя. Хромосомные пары 1 – 22 называют аутосомами. 23 пара половые хромосомы (X и Y): Нормальная женщина имеет XX хромосомы, а нормальный мужчина XY. Ген серповидноклеточной анемии (хромосома 11) Ген цистического фиброза (хромосома 7)
СТРОЕНИЕ ХРОМОСОМЫ Хромосома состоит из двух хроматид. На хромосоме имеется первичная перетяжка – центромера. Центромера делит хромосому на короткое и длинное плечо. Конец хромосомы называется теломером. 1—хроматида; 2—центромера; 3—короткое плечо; 4—длинное плечо
Хромосома – хранение, воспроизведение и передача наследственной информации при размножении клеток и организмов Короткое плечо хромосомы обозначается р, а длинное – q. Каждое плечо хромосомы разделяется на районы, нумеруемые от центромеры к теломере (1-4). Полосы внутри районов нумеруются по порядку от центромеры. Если локализация гена точно известна, для ее обозначения используют индекс полосы. Например, ген, кодирующий фермент эстеразу Д, обозначается 13р14 и означает 4 полоса 1 района короткого плеча 13 хромосомы.
Гетерозигота - организм, имеющий разные аллели одного гена (Аа), образующий несколько сортов гамет и дающий расщепление при разведении “в себе”. Гемизигота - организм, содержащий только один аллель данного гена (ХУ). Встречается у гетерогаметного пола по генам, не имеющим гомолога в У-хромосоме, или по генам, расположенным только в У-хромосоме.
Пара гомологичных хромосом нумеруется одинаково и гены находятся в одних и тех же локусах локус (положение гена) аллель Различные структурные варианты одного гена называют аллельными генами или аллелями Аллельные гены расположены в одной и той же точке хромосом одной пары (гомологичных хромосом). Такую точку называют локус Разные аллели одного гена определяют различные варианты признаков – фены.
Chromosome short (p-) arm long (q-) arm centromere chromatid telomere telomere
При изучении кариотипа определяют следующие морфометрические характеристики хромосом Lа – абсолютная длина хромосомы в мкм; Lр – длина короткого плеча; Lg – длина длинного плеча. Iв – плечевой индекс, Iс – центромерный индекс, Lr – относительная длина хромосомы, Ih - процент гетерохроматиновой зоны, Is – индекс спирализации.
По значению плечевого индекса определяется форма хромосом. При Iв 1-1,9 хромосома называется равноплечей (метацентрической), 2-4,9 – слабонеравноплечей (субметацентрической), 5 и более – акроцентрической или резко неравноплечей.
Аутосомы Аутосомами у живых организмов с хромосомным определением пола называют парные хромосомы, одинаковые у мужских и женских организмов. Аутосомы обозначают порядковыми номерами.
Половые хромосомы Половые хромосомы - хромосомы раздельнополых организмов, в которых расположены гены, определяющие пол и сцепленные с полом признаки организма. В хромосомном наборе клеток млекопитающих и человека особи женского пола имеют две одинаковые (тип ХХ), а мужского пола - неодинаковые (тип ХY) половые хромосомы. У бабочек, птиц, некоторых пресмыкающихся и земноводных - обратное соотношение.
Хромосома эукариот ДНК -50-40% Белки-50-60% : гистоновые (основные) –Н 1, Н2А, Н2В, Н3, Н4 Негистоновые (кислые) – более 100 видов Полисахариды Липиды Ионы металлов
Хромосомы к клетке в зависимости от фазы клеточного цикла бывают: Интерфазные, активные Митотические, неактивные интерфаза митоз
Интерфазные хромосомы – слабо упакованы и готовы к использованию (репликации, транскрипции и др.) Вид интерфазного ядра под микроскопом. Хромосомы активна
Метафазная хромосома видна в микроскоп и неактивна
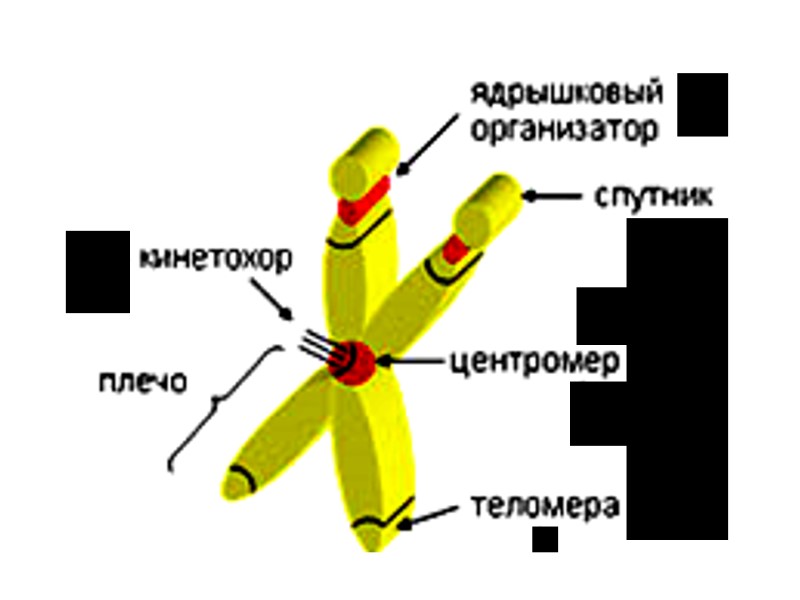
Метафазная хромосома состоит из двух продольных нитей хроматид (дезоксирибонуклеопротеида), соединенных друг с другом в области первичной перетяжки – центромеры, которая делит тело хромосомы на два плеча. Участок каждого плеча вблизи центромеры – проксимальный, а удаленый от нее – дистальный, концевые отделы которых – теломеры – препятствуют соединению концевых участков хромосом. При потере теломер происходят хромосомные перестройки. Некоторые хромосомы могут иметь вторичные перетяжки, отделяющие от тела участок – спутник.
ЦЕНТРОМЕРА (от центр + греч. meros — часть) — участок ДНК, в районе который соединяет хроматиды. ХРОМАТИДА (от греч. chroma - цвет, краска + eidos - вид) — часть хромосомы, которая состоит из молекулы ДНК, соединенной с белками. Хромосомы состоят из хроматина – комплекса ДНК с многочисленными белками.
Компактизация ДНК. ДНК в ядре клеток человека~1 м Cредний d ядра ~10 мкм Длина ДНК в 1 хромосоме человека ~4 см Длина метафазной хромосомы ~4 мкм. ДНК метафазных хромосом человека компактизована по длине в 104 раз
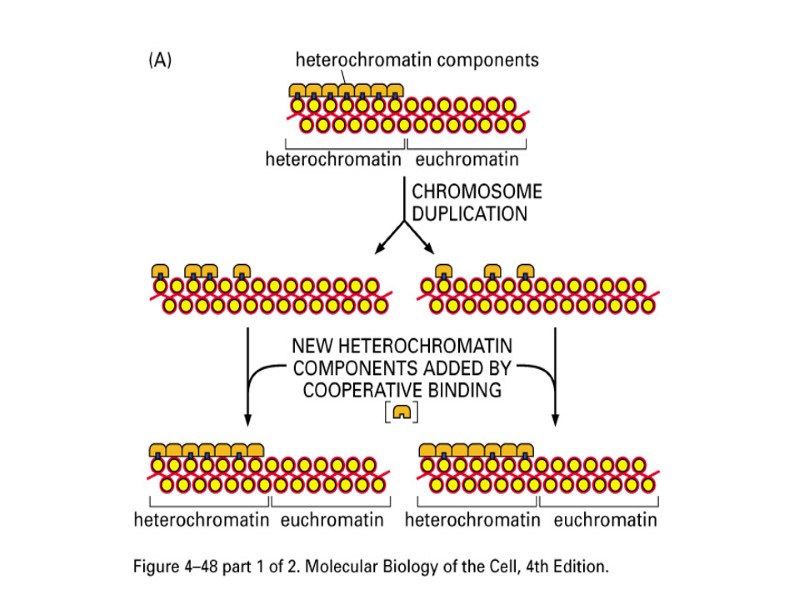
В состав хроматина входят ДНК, РНК, белки, неорганические ионы. При делении клетки ДНК спирализуется, и хроматин преобразуется в хромосомы. ХРОМАТИН - это основное вещество интерфазного ядра. Разные участки хроматина различаются по степени конденсации, или упаковки в пространстве. Эухроматин слабо конденсирован и содержит активно функционирующие гены. Гетерохроматин сильно конденсирован и содержит нефункционирующие гены и участки ДНК, не содержащие гены. Участки гетерохроматина окрашиваются красителями сильнее, чем участки эухроматина и в микроскоп выглядят более темными. При делении клетки хроматин, конденсируясь, приобретает вид плотных палочковидных структур, особенно хорошо видимых в метафазу митоза.
Гетерохроматин и Эухроматин Конденсированое состояние Функционально неактивный Вид плотных глыбок, интенсивно окрашивается красителями Низкая транскрибируемость Факультативный и конститутивный Низкая степень пострансляционных модификаций гистонов Меньшая компактизация ДНК Функционально активные участки хромосом Деконденсированные, светлые участки Локализация активно транскрибирующихся генов
Гетерохроматин 1.Конститутивный (структурный)- локализован в прицентромерных и теломерных районах. Это постоянно выявляемые участки, состоит из умеренно и многократно повторяющихся п.н. 2.Факультативный – участки обратимой компактизации эухроматиновых районов. Пример: одна из двух Х хромосом человека.
Хроматин – комплекс ДНК и белков (гистонов и негистонов) Хроматин Эухроматин (слабо конденсированный, активный) Гетерохроматин (сильно конденсированный, неактивный) Факультативный (содержит гены, не активные в данной клетке в данное время) Конститутивный (структурный) (структурный) не содержит генов ядро ядрышко эухроматин гетерохроматин
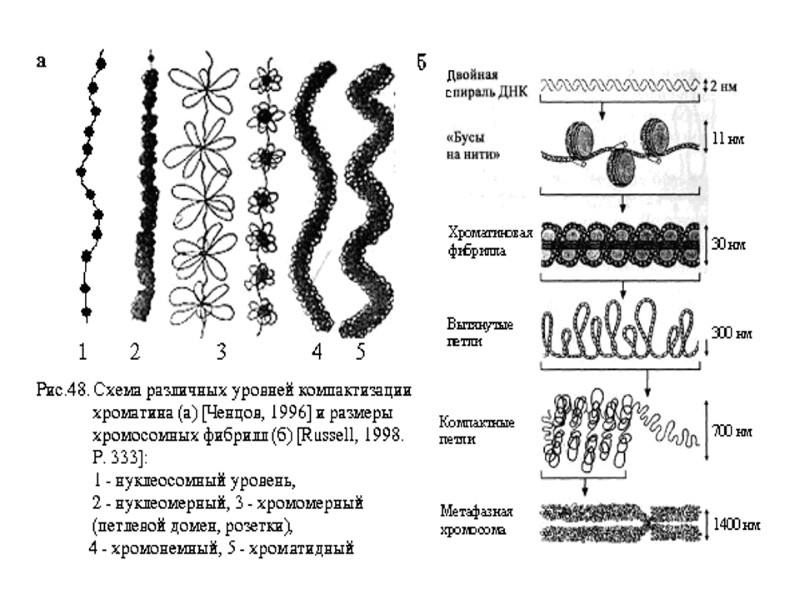
ДНК – 2 нм Нуклеосома – 11 нм Фибрилла 30 нм «соленоид» - 300 нм Нить – 700 нм Метафазная хромосома – 1400 нм активная неактивная ДНК 2 нм Нуклеосома 11 нм Фибрилла 30 нм Фибрилла 300 нм Фибрилла 700 нм Метафазная хромосома 1400 нм
Компактизация -7000 раз Компактизация -3500 раз Компактизация -1000 раз
Строение нуклеосомы ДНК Кор из 8 молекул гистонов 2 х (Н2а, Н2b, H3, H4) Гистон Н1 линкер Нуклеосома: кор- гистоновый октамер и ДНК (146 п.н.). Компактизация в 6-7 раз.
Структурной единицей хроматина является нуклеосома – комплекс из четырех пар гистоновых белков, вокруг которого намотано около двух витков молекулы ДНК. В одной хромосоме находится только одна молекула ДНК, которая намотана на тысячи гистоновых комплексов.
Нуклеосома. d~11 нм 147 п.о. ДНК на один гистоновый октамер ( 1.7 оборота) Гистон Н1: 20п.о. линкерной ДНК
Расположение молекул гистонов в нуклеосоме [Lewin, 2000. Р. 582]
Гистоны Arg -богатые: Н3 и Н4,наиболее консервативные из всех известных белков. Lys:умеренно обогащенныe лизином Н2А и Н2В, сильно богатые лизином гистоны группы H1 Каждый из 4х гистонов содержит центральную глобулярную часть, С и N-концевые участки
Модификация гистонов В регуляции активности генов
Нуклеосома – это октамер гистонов и 1,7 витка суперспирали фрагмента ДНК длиной 146 пар оснований В целом нуклеосомная упаковка ограничивает узнавание последовательности ДНК факторами транскрипции, репликации и рекомбинации По отдельным генам нуклеосомы распределяются неслучайным образом - позиционирование Позиционирование нуклеосом на промоторе может быть фактором регуляции транскрипции, как +, так и -
Ремоделирование нуклеосом – это изменение их связывания с ДНК. Осуществляют белки Перемещение нуклеосом Изменение расстояния между нуклеосомами Удаление гистонов Сборка гистонов
Ковалентная модификация включает – Ацетилирование – деацетилирование Фосфорилирование – дефосфорилирование Метилирование убиквитирование Метилирование лизина по положению 9 в гистоне Н3 – подавление транскрипции, часто это сочетается с метилированием ДНК по цитозину Ацетилирование лизина по положению 9 в гистоне Н3 в сочетании с метилированием лизина в положении 4– подавление транскрипции
Метилирование Arg,Lys Фосфорилирование Модификация гистонов
Убиквитинирование. Убиквитин: 76 ак, кислый белок H2A и H2B : внутриклеточные процессы- инициация и элонгация транскрипции, инактивация генов и репарации ДНК
Ацетилирование гистонов
Связь с метилированием ДНК
Гистоновый код- Аминокислотный остаток – разные АМК могут Подвергаться модификации – Лиз, Арг, Сер, Тре 2. Модифицирующие ферменты – их много 3. Белки, воспринимающие модификацию гистонов Комбинация всех этих факторов – и есть гистоновый код Обеспечивают активацию и сайленсинг генов, очевидно помогая факторам транскрипции
Хроматиновая фибрилла (соленоидный) Гистон Н1 связывается с ДНК между корами. Компактизация – 40 раз.
30nm фибриллы Соленоидная модель. Модель суперспирали Кросс - линкерная модель. Ленточная модель Хромонема -100nm: упаковка более 10,000 п.о.ДНК Гистоны H2AZ Хромосомы
белковый каркас (остов, скаффолд) Нуклеосомы (11 нм) 30 нм Укладка типа «соленоида» 300 нм
Метафазные хромосомы
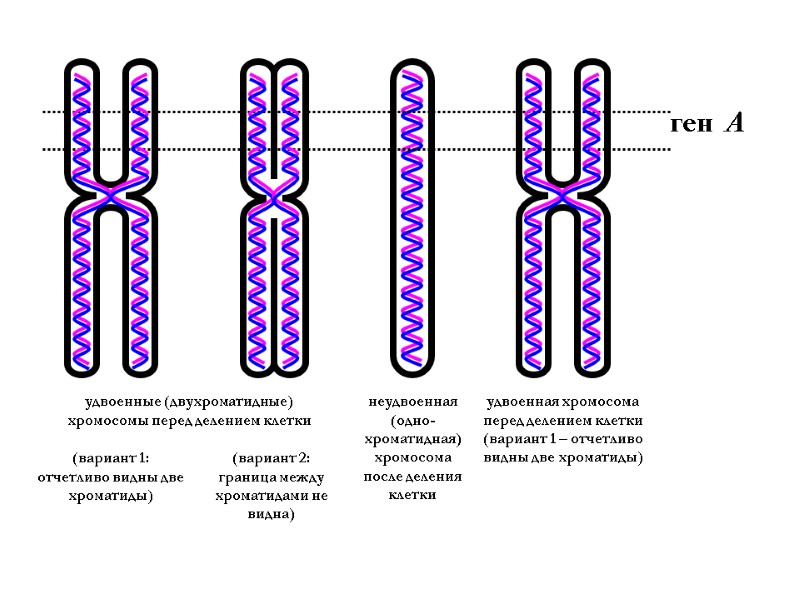
Концы каждой хромосомы – это теломеры. У человека ДНК теломерного участка представляет собой многократно повторяющуюся нуклеотидную последовательность 5' ТТАГГГ 3' в одной из нуклеотидных цепей ДНК. После каждого акта репликации и деления клетки происходит укорочение теломерных участков хромосом. Во время профазы и метафазы митоза каждая хромосома состоит из двух одинаковых хроматид – одинаковых копий материнской хромосомы, образовавшихся после репликации ДНК.
ФУНКЦИИ ХРОМОСОМ Хромосомы – хранители генетической информации. Регулируют процессы в клетке путем синтеза первичной структуры белка, иРНК, рРНК.
ДИПЛОИДНЫЙ НАБОР ХРОМОСОМ КОМАР – 6 ОКУНЬ – 28 ПЧЕЛА – 32 СВИНЬЯ – 38 МАКАК-РЕЗУС –42 КРОЛИК - 44 КРОЛИК – 44 ЧЕЛОВЕК – 46 ШИМПАНЗЕ – 48 БАРАН – 54 ОСЕЛ – 62 ЛОШАДЬ – 64 КУРИЦА - 78
Канадский ученый Барр (1908 – 1995) и его студент Бертрам открыли в 1948 году в ядрах нервных клеток кошек структуру, названную тельце Барра, или Х-половой хроматин
Тельце Барра представлено инактивированной Х хромосомой и находится у женщин в соматических клетках Тельце Барра
Тельце Барра – пример факультативного гетерохроматина, можно видеть в соматических клетках женского организма млекопитающих
Несмотря на то, что женские особи млекопитающих имеют две Х–хромосомы, а мужские – только одну, экспрессия генов Х-хромосомы происходит на одном и том же уровне у обоих полов. Это обусловлено тем, что в каждой клетке самки функционирует лишь одна из двух Х –хромосом. Вторая Х–хромосома на ранних стадиях эмбрионального развития (в период имплантации) инактивируется и превращается в генетически инертное тельце Барра (половой хроматин). При этом в разных клетках отцовская и материнская Х–хромосомы выключаются случайно. Женские особи, гетерозиготные по генам половых хромосом, представляют собой генетические мозаики.
Исследование полового хроматина – тельца Барра У пациента берется соскоб эпителия ротовой полости Помещается на предметное стекло Окрашивается Рассматривается Дешевый экспресс метод определения числа Х хромосом: клетка ядро
Ген, отвечающий за инактивацию Х-хромосомы, на ней же и лежит. Которая из Х-хромосом будет инактивирована, по-видимому, процесс случайный. У черепаховых кошек ген черной и желтой окраски лежит в Х-хромосоме. Распределение пятен абсолютно непредсказуемо.
Широко известный пример проявления такой мозаичности –черепаховые кошки, имеющие черные и желтые (рыжие) пятна. В данном случае окраска определяется аллелями гена O, локализованного на Х-хромосоме: аллель O определяет желтую (рыжую) окраску меха, аллель o –черную. Черепаховые кошки гетерозиготны по этому гену, их генотип –Oo. Желтые и черные пятна у них развиваются в результате случайной инактивации Х–хромосомы с аллелем O или o в разных клетках на ранних этапах эмбриогенеза. Коты могут быть только гемизиготами OY (рыжие) или oY (черные). У гетерозиготных черепаховых кошек (самок) Оо наблюдается кодоминирование, а у гомозиготных котов (самцов) OY или oY –моноаллельное наследование. Коты черепаховую окраску имеют редко, только если в яйцеклетке будет 2 X-хромосомы
Пример. Рыжая кошка скрещивается с черным котом.В их потомстве все коты были рыжие, а кошки имели черепаховую окраску. При скрещивании гибридов первого поколения между собой в их потомстве наблюдалось расщепление –1 часть кошек рыжие :1 часть кошек черепаховые : 1 часть котов рыжие : 1 часть котов черные.
Гибридологический анализ Характер расщепления по цвету шерсти зависит от пола особей. Можно предположить, что ген О локализован в Х-хромосоме. Тогда генотип исходной кошки –ОО, поскольку у нее отсутствуют черные пятна и, соответственно, отсутствует аллель о. Генотип исходного кота –оY, поскольку он гомозиготен, и у него проявляется только аллель о.
Этапы цитогенетического исследования Изучение митотических хромосом – цитогенетика.
Цитогенетика (cytogenetics) раздел генетики, изучающий закономерности наследственности и изменчивости на уровне клетки и субклеточных структур (главным образом хромосом).
Применение цитогенетического метода позволяет изучать нормальную морфологию хромосом и кариотипа, определять генетический пол организма, диагностировать различные хромосомные болезни, связанные с изменением числа хромосом или с нарушением структуры хромосом, изучать процессы мутагенеза на уровне хромосом и кариотипа. Метод широко применяется в медико-генетическом консультировании для целей пренатальной диагностики хромосомных болезней.
Цитогенетический метод Изучение количества и формы хромосом при помощи микроскопа Для изучения кариотипа обычно используют лейкоциты периферической крови, клетки красного костного мозга и некоторые другие клетки.
Цитотогенетический метод
Одним из стимуляторов деления является вещество растительного происхождения фитогемагглютинин (ФГА). Фитогемагглютинин является углеводом обыкновенной фасоли Phaseolus vulgaris, способный агглютинировать эритроциты. Фитогемагглютинин является сильным митогеном – веществом, стимулирующим деление клеток путем митоза.
Под влиянием ФГА клетки начинает делиться путем митоза. Затем в культуральную среду с делящимися клетками добавляют колхицин. Это алкалоид растительного происхождения, обычно получаемый из безвременника (зимовника) осеннего (Colchicum autumnale) или других представителей семейства лилейные. Колхицин препятствует образованию микротрубочек из белка тубулина.
Изучение хромосом Хромосомы заметны только в момент деления. С целью изучения хромосом применяют колхицин, который тормозит стадию анафазы, разрушая нити веретена деления. Хроматиды не расходятся к полюсам клетки и хромосомы имеют вид Х-образных фигур.
Терминология Кинетохор (от греч. kinetós — движущийся и choros — место), механический центр хромосомы, к которому прикрепляются нити веретена деления клетки. Центромера - это область в хромосоме на стадии метафазы митоза, которая менее плотно упакована, образует первичную перетяжку, разделяющую хромосому на два плеча.
Добавление гипотонического раствора – клетки разбухают Х ХХ х х при раскапывании от удара о стекло хромосомы разлетаются в стороны – образуется метафазная пластинка Затем препарат фиксируют и окрашивают Х Х
Кровь (или другой материал) Отделение лейкоцитов Добавление стимулятора митоза – ФГА (фитогемагглютинина) 72 часа Добавление колхицина – блокатора микротрубочек Деление лимфоцитов тормозится на стадии метафазы
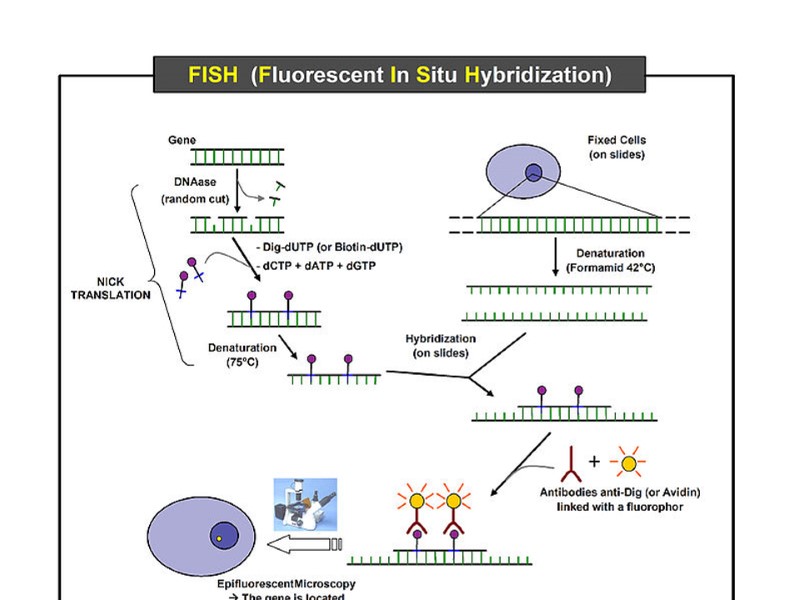
Виды окраски хромосом Рутинная, появилась в 50-х годах ХХ века. (Денверская классификация поделила все хромосомы человека на 7 групп по размеру и форме) Дифференциальная, появилась в конце 60-х годов (G, R, Q и С методы).Парижская конференция закрепила за каждой хромосомой номер, ввела обозначения для мутаций. FISH - метод, был разработан в 90-х годах и дал еще больше возможностей для диагностики.
Виды метафазных хромосом согласно Денверской классификации Метацентрическая, субметацентрическая, акроцентрическая, телоцентрическая, со спутником (первичная перетяжка) вторичная перетяжка короткое плечо (р) длинное плечо (q)
Классификация хромосом Метацентрические хромосомы (М) Субметацентрические (Sm) Субакроцентрические (Sа) Акроцентрические (А) Телоцентрические (Т) Хромосомы типа (М) называют равноплечими. Хромосомы типа (Sm, Sа) называют неравноплечими. Хромосомы типа (А, Т) называют палочковидными.
Классификация хромосом Положение центромеры (перетяжки) используется при классификации хромосом. Центромера
Типы метафазных хромосом [МакКьюсик, 1967. С. 25]
large metacentric small metacentric large acrocentric small acrocentric the rest – submetacentric chromosomes
14 8 10 14 14 + 8 + 14 + 10 = 46 Total number of chromosomes: 46
Кариотип — это совокупность всех хромосом диплоидного набора клетки, который характеризуется количеством хромосом и особенностями строения каждой хромосомы. Каждый вид растений и животных имеет свой видоспецифичный кариотип. Для изучения хромосом используют метод кариотипирования. На рисунке (а) представлена метафазная пластинка хромосом человека. На рисунке (b) представлена раскладка хромосом человека (с учетом размера хромосом, расположения центромеры).
Для нормального кариотипа характерно следующее: присутствует нормальное количество хромосом, все хромосомы представлены парами гомологичных друг другу хромосом, каждая хромосома имеет нормальное строение: характерное для нее расположение центромеры, соотношение и строение плеч, отсутствуют хромосомные мутации.
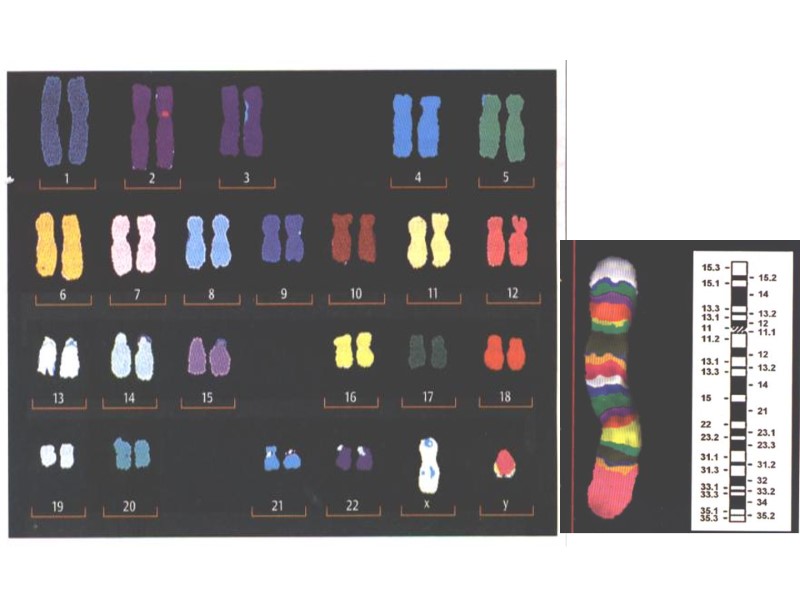
Кариограмма – это изображение всех хромосом диплоидного набора клетки, которые распределены по группам и расположены друг за другом в порядке уменьшения размеров с учетом индивидуальных особенностей каждой хромосомы.
ВСЕ ХРОМОСОМЫ ЧЕЛОВЕКА
Нерасхождение хромосом в мейозе дает нарушение во всех клетках потомка (генеративная мутация). Нерасхождение при митозе дает нарушение только в потомстве данной клетки (соматическая мутация). Такой организм называется «мозаик».
Классификация хромосом Денверская (7 групп) Парижская (дифференциальная окраска) Q, G, R, C и др. Идиограмма
В 1960 году была предложена Денверская классификация хромосом Которая учитывает размер и форму хромосом, положение центромеры, наличие вторичных перетяжек и спутников. 23 пары хромосом делят на 7 групп от А до G. Важно учитывать центромерный индекс (ЦИ), который отражает отношение (в %) длины короткого плеча к длине всей хромосомы.
К группе А относят 1-3 пары хромосом. Это большие метацентрические и субметацентрические хромосомы, их ЦИ составляет 38-49. К группе В относят 4 и 5 пары, большие субметацентрические хромосомы, ЦИ 24-30. К группе С относят 6-12 пары, среднего размера субметацентрические хромосомы, ЦИ 27-35, а также половую Х хромосому. К группе D (13-15 пары), акроцентрические, ЦИ около 15. К группе Е (16-18 пары), относительно короткие метацентрические или субметацентрические, ЦИ 26-40. К группе F (19-20 пары), короткие, субметацентрические, ЦИ 36-46. К группе G (21-22 пары, Y-хромосомы) маленькие, акроцентрические, ЦИ 13-33.
Классификация хромосом Группа А (1-3 пары): крупные хромосомы, отличимые друг от друга; центромеры находятся в середине. Группа В (4-5): крупные хромосомы, мало отличимые друг от друга; центромеры смещены к концам.
Классификация хромосом Группа С (6-12): хромосомы средних размеров, трудно отличимые друг от друга; центромеры смещены к концам. С хромосомами этой группы сходна Х-хромосома. Группа D (13-15): хромосомы средних размеров, центромеры почти полностью смещены к концам.
Классификация хромосом Группа Е (16-18): короткие хромосомы. Группа F (19-20): мелкие, центромеры посередине. Группа G (21-22): самые маленькие, центромеры на концах. С хромосомами группы G сходна У-хромосома.
Парижская классификация хромосом человека 1971г. В основе данной классификации лежат методы специальной, дифференциальной их окраски, при которой в каждой хромосоме выявляется характерный только для нее порядок чередования поперечных темных и светлых сегментов. В настоящее время для идентификации хромосом в соответствии с номенклатурой ISCN-1995 (парижская номенклатура) все чаще используется дифференциальное окрашивание, которое на хромосомах дает полосы поперечной исчерченности, благодаря которым можно более точно идентифицировать пары гомологов.
Обозначение полиморфизма хромосом человека
Систематические изображение G – сегментов хромосом человека и система их обозначения согласно решениям Парижской конференции в 1971г. Цифрами обозначены номер хромосом; X и Y - половые хромосомы ; р – короткое, q – длинное плечи хромосом
Различные типы сегментов обозначают по методам, с помощью которых они выявляются более четко. Q-cсегмент – это участки хромосом, флюоресцирующие после окрашивания акрихин-ипритом; G- сегменты выявляются при окрашивании красителем Гимза (Q и G сегменты идентичны). R сегменты окрашиваются только после тепловой денатурации. Эти методы позволяют четко определять хромосомы внутри групп.
G-окрашивание
Q-окрашивание photo scheme
R-staining (reversed to G) Treated with temperature Giemsa stain is used)
Т-staining acridine orange+temperature
Giemsa staining (= solid or conventional staining) leucocytes chromosomes
Fluorochromes in genetics DAPI (4',6-diamidino-2-phenylindole) PI (propidium iodide) Quinacrine Ethidium bromide
Идиограмма Это систематизированный кариотип, в котором хромосомы располагаются по мере убывания их величины. Точно расположить хромосомы по величине удается не всегда, так как некоторые пары хромосом имеют близкие размеры.
Total Genes On Chromosome: 723 373 genes in region marked red, 20 are shown FZD2 AKAP10 ITGB4 KRTHA8 WD1 SOST MPP3 MLLT6 STAT3 BRCA1 breast cancer 1, early onset GFAP NRXN4 NSF NGFR CACNB1 HOXB9 HTLVR ABCA5 CDC6 ITGB3 Chromosome 17 source: Human Genome Project Genes are arranged in linear order on chromosomes
Плечи делят на районы (бенды) и суббенды
FISH -метод – Fluorescent in situ hybridization дал еще больше возможностей
Цветная исчерченность хромосом человека a – метафазная пластинка; b – pаскладка хромосом.
FISH-метод позволяет лучше распознавать хромосомные перестройки, чем одноцветная окраска
Исследование кариотипа (кариотипирование = цитогенетическое исследование позволяет диагностировать хромосомные и геномные мутации Хромосомные мутации – изменение строения хромосом. Геномные мутации – изменение числа хромосом
p q p q p q p q Нормальное расхождение в анафазе Изохромосомы (isochromosome) – аномальная хромосома, которая образуется на стадии анафазы деления клеток, когда центромер делится горизонтально, а не продольно. В результате образуются одна хромосома с двумя длинными плечами и одна с двумя короткими.
Образование кольцевой хромосомы (r) кольцо ацентрические фрагменты
Хромосомные карты Генетические – где лежит какой ген Цитологические – по окраске Физические – основаны на точном расстоянии между генами Рестрикционные – вид физической карты, на которой указаны расстояния между соседними сайтами расщепления ДНК определенной рестриктазой (разрезающим ферментом) Карты часто бывают комбинированные 1 сМ (сентиморган = морганида) – единица расстояния между генами, при которой вероятность кроссинговера равна 1%, (соответствует примерно 1 мегабазе) Гаплоидный геном человека составляет примерно 3 300 000 000 баз, т.е. 3300 сМ
Генетические и физические карты хромосом Генетическое картирование основывается на использовании генетических методов для построения карт, указывающих позиции генов и других последовательностей в геноме. 148 КАРТИРОВАНИЕ ХРОМОСОМ – ОПРЕДЕЛЕНИЕ ПОЛОЖЕНИЯ ГЕНА СООТВЕТСТВУЮЩЕГО ПРИЗНАКА в ИЗВЕСТНОМ ЛОКУСЕ КОНКРЕТНОЙ ХРОМОСОМЫ; ВАЖНА ИЗБИРАТЕЛЬНАЯ ОКРАСКА ХРОМОСОМ (ЗАПИСЬ 22q11.2 – ГЕН РАСПОЛОЖЕН во 2-м субсегменте 1-го сегмента 1-го района длинного плеча хромосомы 22):
Физическое картирование использует молекулярно-биологические методы для непосредственного исследования молекул ДНК и создания карт, обозначающих позиции определенных последовательностей, в том числе генов 149
На генетической карте хромосом обозначают: номер группы сцепления, сокращенные названия генов, расстояние между генами С помощью генетических карт можно предвидеть характер наследования признаков, определяемых нанесенными генами: теоретически расчитывать вероятность их расщепления у потомков 150 Физические карты - более точны!
Методы физического картирования хромосом Большое число методов физического картирования хромосом основано на 3 главных подходах: Рестрикционное картирование, устанавливающее относительные позиции на молекуле ДНК последовательностей узнавания для рестрикционных эндонуклеаз Флуоресцентная in situ гибридизация (FISH), при которой расположение маркера картируется гибридизацией образца, который содержит маркер с интактными хромосомами Картирование последовательностями маркерных сайтов (STS-картирование), при котором позиции коротких последовательностей картируются ПЦР и/или гибридизационным анализом фрагментов генома 151
Гены болезней, картированные в первой хромосоме [McKusick, 1992. - Из:В.П.Пузырев,1996.С. 25 ].
Карта хромосомы 9
Карта хромосомы 21 и митохондриального генома
Последовательности, распознаваемые разными рестриктазами EcoRI Г ААТТЦ ЦТТАА Г SmaI ЦЦЦ ГГГ ГГГ ЦЦЦ ДНК разрезают рестриктазами и подвергают электрофорезу. Рестрикционная карта - вид физической карты, на которой указаны расстояния между соседними сайтами расщепления ДНК определенной рестриктазой.
Методы создания хромосомных карт Гибридологический (по результатам скрещиваний. Т.Морган) Анализ родословных (генеалогический) Генетики соматических клеток* ДНК-зондов* Будет обсуждаться позже
В данное время для картирования генов хромосом человека применяются также другие методы: Биохимические методы — сравнение аминокислотных последовательностей белков и нуклеотидной последовательности ДНК отдельных хромосом Цитологические методы — сопоставление изменений морфологии хромосомного участка с характерным фенотипом, анализ «ломких» участков хромосом Молекулярно-генетические методы и др. 157
Благодаря картированию удалось установить локализацию многих генов в хромосомах человека Гены располагаются в хромосомах довольно скученно, образуя группы, между которыми могут находиться большие незанятые участки В различных хромосомах находится различное число генов Наибольшее число генов - свыше 3 000 в наибольшей из аутосом - первой В Х-хромосоме их известно свыше 1400 158
Хромосома 1 Хромосома 4
Х-хромосома
Аллели группы крови по системе АВ0, находятся в 9-й хромосоме группы крови по системе MN - во 2-й системы резус-фактора (Rh) - в 1-й хромосоме главного комплекса гистосовместимости – в 6-й хромосоме Свыше 200 генов напрямую унаследованы нами от бактерий Повторяющиеся последовательности ДНК, которые ранее считались лишними, могут оказаться «черным ящиком» эволюции и рассказать о предыдущих 800 млн. годах развития органического мира 161
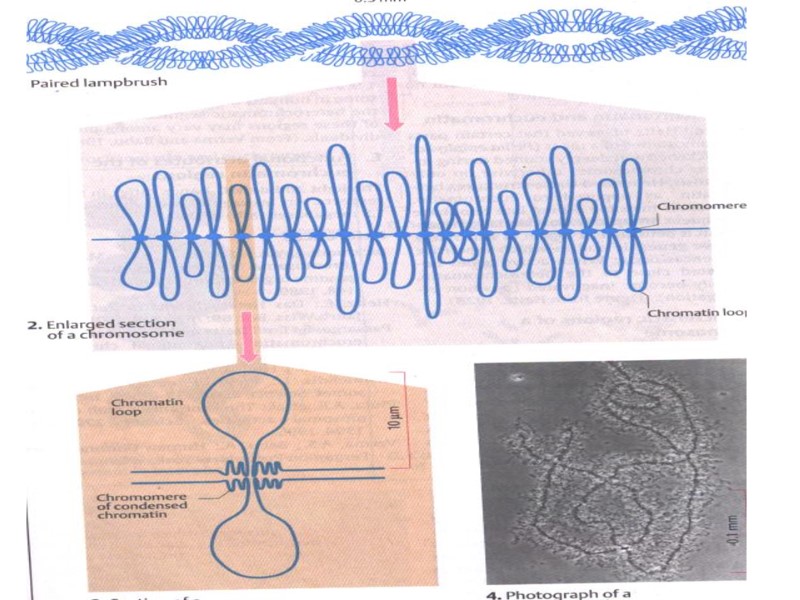
Необычные виды хромосом «ламповые щетки» (найдены в овоцитах амфибий) политенные хромосомы (в слюнных железах личинок двукрылых)
Гигантские хромосомы Это модели для изучения генома. Хромосомы типа «ламповых щеток» - длина их в 30 раз больше длины обычных. Имеют петли. Образуются во время мейоза, в конце мейоза возвращаются в обычное состояние. Изучены у амфибий.
Хромосомы типа ламповых щеток В них активны гены, обеспечивающие образование желтка для развития зародыша
Гигантские хромосомы Политенные хромосомы- образуются в некоторых клетках в результате максимальной деспирализации и многократной репликации без последующего расхождения хромосом (эндомитоза). Политения (от поли... и лат. taenia — повязка, лента), наличие в ядре некоторых соматических клеток гигантских многонитчатых (политенных) хромосом, превышающих в сотни раз обычные. П. приводит к значительному увеличению плоидности ядер
Политенные (многонитчатые) хромосомы Репликация ДНК не сопровождается делением клетки, что приводит к накоплению вновь построенных нитей ДНК. Большое количество копий генов на политенных хромосомах позоляет синтезировать больше нужных личинке белков. 1 — диски; 2 — междисковые участки; 3 — пуф, образовавшийся за счет деконденсации хроматина диска
Политенные хромосомы Диски – участки более плотно упакованного хроматина. Каждый диск – самостоятельная функциональная единица, содержащая от одного до нескольких генов. Во время экспрессии активные диски вздуваются и образуют пуфы. Пуфы появляются и исчезают в определенной последовательности на разных стадиях онтогенеза.
Активные участки -пуфы Неактивные - диски Политенные хромосомы дрозофилы
БИБЛИОГРАФИЧЕСКИЙ СПИСОК Основной. Смирнов В.Г. Цитогенетика. – М.:Высшая школа, 1991. Абрамова А.И., Устинова Е.И. Цитология растений. М.,Колос, 1980. Жимулев И.Ф. Общая и молекулярная генетика. Изд-во Новосиб.ун-та, 2002. Босток К., Самнер Э. Хромосома эукариотической клетки. М., Мир, 1981. 5. Генетика, биохимия и цитология мейоза. М., Наука, 1982. Дополнительный. Макаров В.Б., Сафронов В.В. Цитогенетические методы анализа хромосом. – М., Наука, 1978. Дарлингтон С.., Лакур Л.Ф. Хромосомы. Методы работы. М.,Атомиздат, 1980. Ригер Р.Михаэлис А. Генетический и цитогенетический словарь. М.Мир, 1967. Ченцов Ю.С. Общая цитология. М.,МГУ, 1984. Эмбриология поктрытосеменных растений с основами цитологии. М.МГУ,1965. Паушева В.П. Практикум по цитологии растений. 4-е изд., М.: Агропромиздат, 1988. Эмбриология растений. М.: Агропромиздат, 1990. Ячевская Г.Л., Иванова С.В., Наумов А.А. Особенности мейоза при отдаленной гибридизации. М.: Изд-во МСХА, 1990. Жученко А.А., Король А.Б. Рекомбинации в эволюции и селекции. М.: Наука, 1985. Билич Г., Катинас Г.С., Назарова Л.В. Цитология, Санкт-Петербург, 1999..