Лекция 5-6. 22,27.02.14.ppt
- Количество слайдов: 96





 Характеристики популяций
Характеристики популяций
 N d. N /dt. N t r d. N /dt. N = b - d = r (Const) d. N/dt= r. N N
N d. N /dt. N t r d. N /dt. N = b - d = r (Const) d. N/dt= r. N N
, мальтузианский параметр b max r max b r d d") Биотический потенциал (Chapmen, 1928), мальтузианский параметр b max r max b r d d min N
Биотический потенциал (Chapmen, 1928), мальтузианский параметр b max r max b r d d min N
 экспоненциально, устанавливается") Ещё одно важное обстоятельство, касающееся экспоненциального роста: В популяции, растущей (или убывающей) экспоненциально, устанавливается стабильное возрастное распределение
Ещё одно важное обстоятельство, касающееся экспоненциального роста: В популяции, растущей (или убывающей) экспоненциально, устанавливается стабильное возрастное распределение
N + r =") d. N /dt. N rmax d. N /dt. N = (-r/K)N + r = = r (-N/K + 1) = r (1 – N/K) N t+1 = Nt er(1 -N/K) К N
d. N /dt. N rmax d. N /dt. N = (-r/K)N + r = = r (-N/K + 1) = r (1 – N/K) N t+1 = Nt er(1 -N/K) К N
 +∞ r 0 -∞ r = ln. R 0 /T +∞ 1 R 0
+∞ r 0 -∞ r = ln. R 0 /T +∞ 1 R 0
 Приближённый способ расчета удельной скорости роста численности r Экспоненциальный рост численности. Почему? Объяснение на следую-щих слайдах Почему за среднее время генерации G численность популяции возрастает примерно в R 0 раз? Потому что R 0, как мы только что показали, – это количество потомков, которые произведены самкой за время ее жизни, дожили до следующего периода размножения и, таким образом, составили следующее поколе-ние. То есть R 0 – это отношение численностей последовательных поколений, Почему равенство для r является приближённым? Потому что в ходе нашего вывода формулы для r непрерывный процесс размножения заменяется дискретным (поколения дискретны!). Но непрерывное равно дискретному только приблизительно!
Приближённый способ расчета удельной скорости роста численности r Экспоненциальный рост численности. Почему? Объяснение на следую-щих слайдах Почему за среднее время генерации G численность популяции возрастает примерно в R 0 раз? Потому что R 0, как мы только что показали, – это количество потомков, которые произведены самкой за время ее жизни, дожили до следующего периода размножения и, таким образом, составили следующее поколе-ние. То есть R 0 – это отношение численностей последовательных поколений, Почему равенство для r является приближённым? Потому что в ходе нашего вывода формулы для r непрерывный процесс размножения заменяется дискретным (поколения дискретны!). Но непрерывное равно дискретному только приблизительно!
 Кому давать орден «Мать-героиня» ? Из лекций А. М. Гилярова
Кому давать орден «Мать-героиня» ? Из лекций А. М. Гилярова
 Кому давать орден «Мать-героиня» ? Популяция, где по пять детей у молодых матерей (в год по ребенку с 18 до 22 лет): R 0 = 2. 5 G = 20 лет Популяция, где по десять детей у более взрослых матерей (в год по ребенку с 31 до 40 лет): R 0 = 5 G = 35. 5 лет Из лекций А. М. Гилярова
Кому давать орден «Мать-героиня» ? Популяция, где по пять детей у молодых матерей (в год по ребенку с 18 до 22 лет): R 0 = 2. 5 G = 20 лет Популяция, где по десять детей у более взрослых матерей (в год по ребенку с 31 до 40 лет): R 0 = 5 G = 35. 5 лет Из лекций А. М. Гилярова
 / 20 =") Кому давать орден «Мать-героиня» ? r 1 = (ln 2. 5) / 20 = -1 0. 916 / 20 = 0. 0458 год r 2 = (ln 5) / 35. 5 = 1. 609 / 35. 5 = 0. 0453 лекций А. М. год-1 Из Гилярова Замечание 1. А почему все-таки правы те, кто присуждали орден за 10 детей? Потому что r 1 может быть реализована только в чреде поколений, то есть если дочери этих матерей будут заводить столько же детей в столь же раннем возрасте (что, конечно, совсем необязательно) Замечание 2. И величина r 1, и величина r 2, конечно, завышены, поскольку не учитывают, что la < 1. (Напомним, что R 0 = b∙Ta∙la. ) Замечание 3 (точнее, вопрос). Почему r 1 > r 2 несмотря на то, что R 0 у молодых матерей в 2 раза меньше? Ответ «потому что время генерации у них меньше» не годится, потому что время генерации у них меньше
Кому давать орден «Мать-героиня» ? r 1 = (ln 2. 5) / 20 = -1 0. 916 / 20 = 0. 0458 год r 2 = (ln 5) / 35. 5 = 1. 609 / 35. 5 = 0. 0453 лекций А. М. год-1 Из Гилярова Замечание 1. А почему все-таки правы те, кто присуждали орден за 10 детей? Потому что r 1 может быть реализована только в чреде поколений, то есть если дочери этих матерей будут заводить столько же детей в столь же раннем возрасте (что, конечно, совсем необязательно) Замечание 2. И величина r 1, и величина r 2, конечно, завышены, поскольку не учитывают, что la < 1. (Напомним, что R 0 = b∙Ta∙la. ) Замечание 3 (точнее, вопрос). Почему r 1 > r 2 несмотря на то, что R 0 у молодых матерей в 2 раза меньше? Ответ «потому что время генерации у них меньше» не годится, потому что время генерации у них меньше
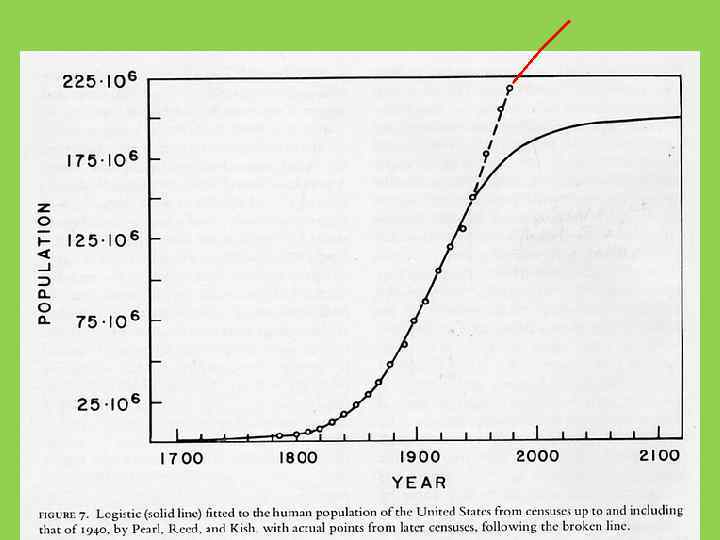
 Логистический (S-образный, сигмоидный) рост замкнутой популяции (r линейно убывает вслед") ОСНОВНЫЕ УРАВНЕНИЯ (ЛОГИСТИЧЕСКИЙ РОСТ) Логистический (S-образный, сигмоидный) рост замкнутой популяции (r линейно убывает вслед за N пересекая ось в точке N=K) Для непрерывного размножения: d. N/dt = r. N (1 - N/K) (Ферхюльст – Пирл) N t = K/(1+be-rt); b = (K-No)/No Для дискретного: Nt+1= N t. R / (1 + *Ntc) где К- плотность насыщения, c – параметр запаздывания, =(R-1)/K
ОСНОВНЫЕ УРАВНЕНИЯ (ЛОГИСТИЧЕСКИЙ РОСТ) Логистический (S-образный, сигмоидный) рост замкнутой популяции (r линейно убывает вслед за N пересекая ось в точке N=K) Для непрерывного размножения: d. N/dt = r. N (1 - N/K) (Ферхюльст – Пирл) N t = K/(1+be-rt); b = (K-No)/No Для дискретного: Nt+1= N t. R / (1 + *Ntc) где К- плотность насыщения, c – параметр запаздывания, =(R-1)/K
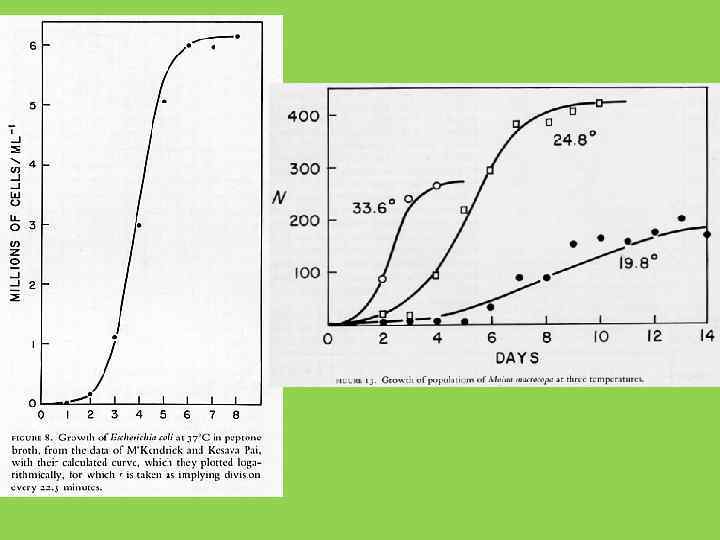
 S – образный, близкий к логистическому, рост популяций может наблюдаться при самых разных механизмах ограничения плотности: 1) нехватки пищи (дафнии, водоросли, бактерии) 2) накопление продуктов метаболизма (дрожжи) 3) каннибализм (мучные жуки Tribolium) 4) поведенческие механизмы (домовые мыши в эксперименте) … и т. д.
S – образный, близкий к логистическому, рост популяций может наблюдаться при самых разных механизмах ограничения плотности: 1) нехватки пищи (дафнии, водоросли, бактерии) 2) накопление продуктов метаболизма (дрожжи) 3) каннибализм (мучные жуки Tribolium) 4) поведенческие механизмы (домовые мыши в эксперименте) … и т. д.
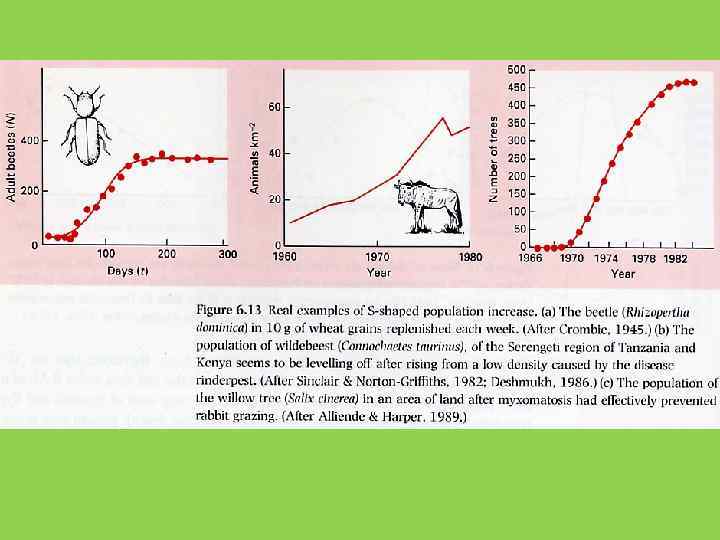
 Модель логистического роста не более, чем одна из множества моделей, описывающих S-образный рост
Модель логистического роста не более, чем одна из множества моделей, описывающих S-образный рост
 (Left) The relationships between population growth rates (pgr) and size (N)") (A to C) (Left) The relationships between population growth rates (pgr) and size (N) with (right) their associated population time series. . The data come from three insect populations (A) тля - Acyrthosiphon pisum, (B) бабочка павлиний глаз - Inachis io, (C) совка - Xylena vetusta
(A to C) (Left) The relationships between population growth rates (pgr) and size (N) with (right) their associated population time series. . The data come from three insect populations (A) тля - Acyrthosiphon pisum, (B) бабочка павлиний глаз - Inachis io, (C) совка - Xylena vetusta
 Где θ – показатель, характеризующий форму зависимости удельной скорости популяционного роста r 0 от плотности
Где θ – показатель, характеризующий форму зависимости удельной скорости популяционного роста r 0 от плотности
 Разные формы зависимости удельной скорости популяционного роста r 0 от плотности популяции. Линейная зависимость при θ=1, вогнутая при θ<1
Разные формы зависимости удельной скорости популяционного роста r 0 от плотности популяции. Линейная зависимость при θ=1, вогнутая при θ<1
 млекопитающие, (B)") Значения показателя тета для четырех таксономических группа из имеющейся базы данных: (A) млекопитающие, (B) птицы, (C) рыбы, (D) насекомые. Больше +1 – выпуклая, меньше +1 - вогнутая
Значения показателя тета для четырех таксономических группа из имеющейся базы данных: (A) млекопитающие, (B) птицы, (C) рыбы, (D) насекомые. Больше +1 – выпуклая, меньше +1 - вогнутая
, показано, что скорость популяционного роста высока") На 1780 сериях наблюдений (млекопитающие, птицы, рыбы, насекомые), показано, что скорость популяционного роста высока при низких значениях плотности, но она очень быстро падает при возрастании плотности. Зависимость описывается вогнутой кривой. Отсюда следует, что большинство популяций в природе существуют при плотностях, близких к ёмкости среды (величина K).
На 1780 сериях наблюдений (млекопитающие, птицы, рыбы, насекомые), показано, что скорость популяционного роста высока при низких значениях плотности, но она очень быстро падает при возрастании плотности. Зависимость описывается вогнутой кривой. Отсюда следует, что большинство популяций в природе существуют при плотностях, близких к ёмкости среды (величина K).
 N R c Случайные колебания Предельные циклы Затухающие Монотонно затухающие Nt+1= Nt R / (1+a*Ntc) время
N R c Случайные колебания Предельные циклы Затухающие Монотонно затухающие Nt+1= Nt R / (1+a*Ntc) время
 r-стратегия К-стратегия «отбор на количество» «отбор на качество» ,") СТРАТЕГИИ ПОПУЛЯЦИИ (Mc. Arthur, 1967) r-стратегия К-стратегия «отбор на количество» «отбор на качество» , повышение Основные признаки r-видов: устойчивости • высокая плодовитость, Основные признаки К-видов: • короткое время жизни, • низкая плодовитость, • обычно малые размеры • значительная особей (у растений мелкие продолжительность жизни, семена), • крупные размеры особей и • низкая энергия поддержания, семян, мощные корневые системы, • высокие траты энергии на размножение, • высокая конкурентоспособность, • Высокая скорость устойчивость на занимаемой расселения, территории, • низкая • высокая специализация конкурентоспособность образа жизни. • забота о потомстве
СТРАТЕГИИ ПОПУЛЯЦИИ (Mc. Arthur, 1967) r-стратегия К-стратегия «отбор на количество» «отбор на качество» , повышение Основные признаки r-видов: устойчивости • высокая плодовитость, Основные признаки К-видов: • короткое время жизни, • низкая плодовитость, • обычно малые размеры • значительная особей (у растений мелкие продолжительность жизни, семена), • крупные размеры особей и • низкая энергия поддержания, семян, мощные корневые системы, • высокие траты энергии на размножение, • высокая конкурентоспособность, • Высокая скорость устойчивость на занимаемой расселения, территории, • низкая • высокая специализация конкурентоспособность образа жизни. • забота о потомстве
 Jack of all trades, master of none
Jack of all trades, master of none
 is a situation that involves losing one quality or aspect") A trade-off (or tradeoff) is a situation that involves losing one quality or aspect of something in return for gaining another quality or aspect
A trade-off (or tradeoff) is a situation that involves losing one quality or aspect of something in return for gaining another quality or aspect
 Характеристика r-стратегия K–стратегия Численность популяции Очень изменчива, может быть больше K Обычно близка к K Оптимальный тип местообитания или климата Изменчивый и(или) непредсказуемый Более-менее постоянный, предсказуемый Смертность Обычно катастрофическая Небольшая Размер популяции Изменчивый во времени, неравновесный Конкуренция Часто слабая Онтогенетические особенности Быстрое развитие, раннее размножение Небольшие размеры Единственное размножение Много потомков Короткая жизнь (менее 1 года) Способность к Быстрое и широкое Относительно постоянный, равновесный Обычно острая Относительно медленное развитие Позднее размножение Крупные размеры Многократное размножение Мало потомков Долгая жизнь (больше 1 года)
Характеристика r-стратегия K–стратегия Численность популяции Очень изменчива, может быть больше K Обычно близка к K Оптимальный тип местообитания или климата Изменчивый и(или) непредсказуемый Более-менее постоянный, предсказуемый Смертность Обычно катастрофическая Небольшая Размер популяции Изменчивый во времени, неравновесный Конкуренция Часто слабая Онтогенетические особенности Быстрое развитие, раннее размножение Небольшие размеры Единственное размножение Много потомков Короткая жизнь (менее 1 года) Способность к Быстрое и широкое Относительно постоянный, равновесный Обычно острая Относительно медленное развитие Позднее размножение Крупные размеры Многократное размножение Мало потомков Долгая жизнь (больше 1 года)
 N r-стратегия К-стратегия Заселение неблагоприят ной ниши время
N r-стратегия К-стратегия Заселение неблагоприят ной ниши время
 Виоленты (силовики) – энергично развиваясь, они захватывают") жизненные стратегии растений (по Л. Г. Раменскому) Виоленты (силовики) – энергично развиваясь, они захватывают территорию и удерживают ее за собой, подавляя соперников полнотой использования ресурсов среды аналоги К-стратегии. Патиенты (выносливцы) – в борьбе за выживание они берут не энергией жизнедеятельности и роста, а своей выносливостью к крайним условиям (ксерофиты – типчак, тенелюб – кислица) - переходная L-стратегия. Эксплеренты(заполняющие) – имеют очень низкую конкурентную мощность, но зато они способны очень быстро захватывать освобождающиеся территории, выполняя промежутки между сильными растениями, аналоги R-стратегии
жизненные стратегии растений (по Л. Г. Раменскому) Виоленты (силовики) – энергично развиваясь, они захватывают территорию и удерживают ее за собой, подавляя соперников полнотой использования ресурсов среды аналоги К-стратегии. Патиенты (выносливцы) – в борьбе за выживание они берут не энергией жизнедеятельности и роста, а своей выносливостью к крайним условиям (ксерофиты – типчак, тенелюб – кислица) - переходная L-стратегия. Эксплеренты(заполняющие) – имеют очень низкую конкурентную мощность, но зато они способны очень быстро захватывать освобождающиеся территории, выполняя промежутки между сильными растениями, аналоги R-стратегии
 Эксплерент Виолент (С)") Сравнительная характеристика типов стратегий растений Признак Тип стратегии (Раменский - Грайм) Эксплерент Виолент (С) Патиент (S) (R) Абиотические Благоприятн Неблагоприят Благоприятн условия среды ые ные ые Наличие Нет Есть нарушений Уровень конкуренции Высокий Низкий Высокий между особями
Сравнительная характеристика типов стратегий растений Признак Тип стратегии (Раменский - Грайм) Эксплерент Виолент (С) Патиент (S) (R) Абиотические Благоприятн Неблагоприят Благоприятн условия среды ые ные ые Наличие Нет Есть нарушений Уровень конкуренции Высокий Низкий Высокий между особями
 «Треугольник» Дж. Грайма
«Треугольник» Дж. Грайма
 Эффект “bottleneck”в динамике популяции
Эффект “bottleneck”в динамике популяции
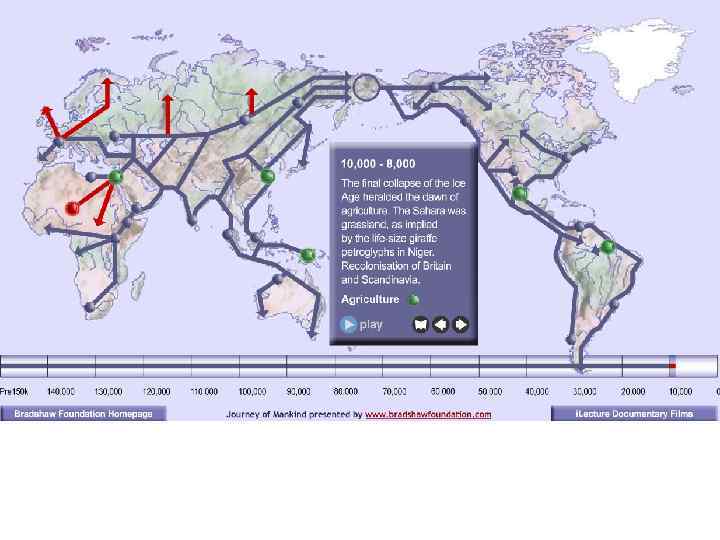
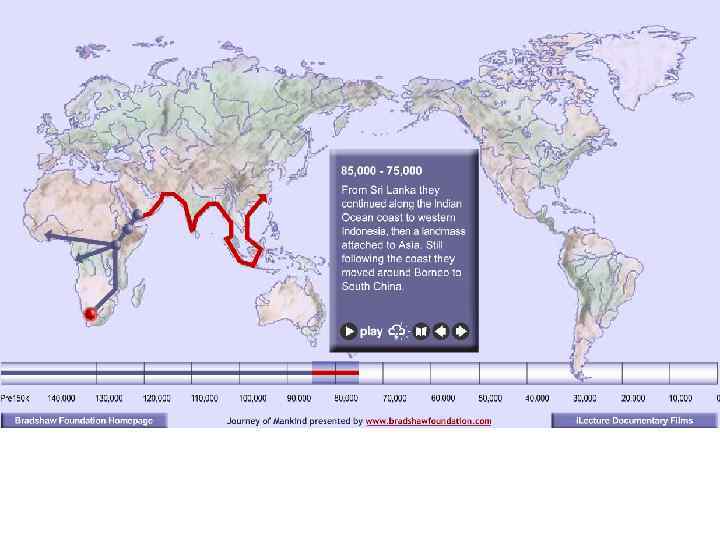
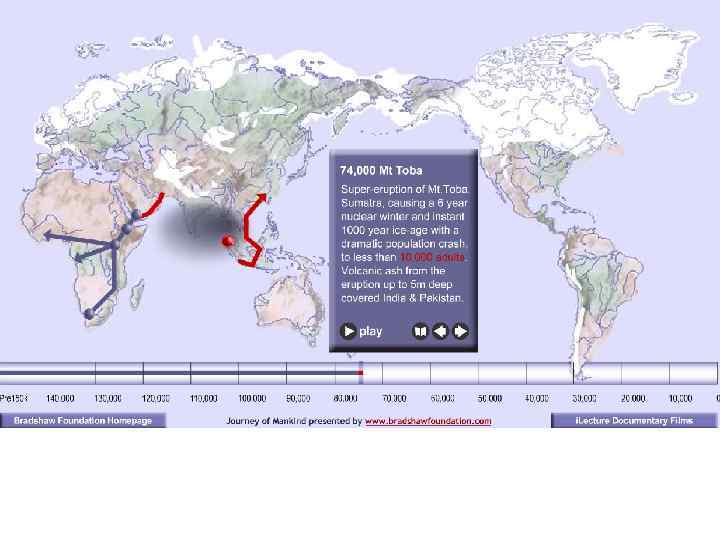
 Серия слайдов, показывающая расселение Homo sapiens из восточных районов Центральной Африки
Серия слайдов, показывающая расселение Homo sapiens из восточных районов Центральной Африки
 Так какая же стратегия поддержания численности лучше?
Так какая же стратегия поддержания численности лучше?
 Кривая уязвимости как функция годовой плодовитости и положение на ней некоторых видов. Описывается уравнением имеет вид: L = 1/[1 + exp(-1. 52 + 2. 10 B)]. Чем больше плодовитость, тем меньше вероятность оказаться под угрозой вымирания (по Л. В. Полищуку, 2009). Для видов с годовой плодовитостью выше 2. 9 дочерей /год вероятность оказаться под угрозой вымирания меньше 0. 01
Кривая уязвимости как функция годовой плодовитости и положение на ней некоторых видов. Описывается уравнением имеет вид: L = 1/[1 + exp(-1. 52 + 2. 10 B)]. Чем больше плодовитость, тем меньше вероятность оказаться под угрозой вымирания (по Л. В. Полищуку, 2009). Для видов с годовой плодовитостью выше 2. 9 дочерей /год вероятность оказаться под угрозой вымирания меньше 0. 01
 кривые дожития кривые выживания lx – кривые
кривые дожития кривые выживания lx – кривые
 Число выживших логарифмическая шкала ВОЗРАСТНЫЕ КОГОРТЫ 1000 10 1 Тип II. Некоторые птицы, беспозвоночн Тип III. ые (гидра), Некоторые насекомые, растения и водные беспозвоночные, рептилии моллюски, рыбы, растения Тип I. Человек, крупные млекопитаю щие с заботой о потомстве Возраст 0
Число выживших логарифмическая шкала ВОЗРАСТНЫЕ КОГОРТЫ 1000 10 1 Тип II. Некоторые птицы, беспозвоночн Тип III. ые (гидра), Некоторые насекомые, растения и водные беспозвоночные, рептилии моллюски, рыбы, растения Тип I. Человек, крупные млекопитаю щие с заботой о потомстве Возраст 0
 Кривая типа I – выпуклая – характерна для человека и для большинства млекопитающих. Однако обратите внимание, что для человека и тем более для млекопитающих, живущих в дикой природе, в отличие от идеального типа I начальный этап кривой выживания характеризуется заметным спадом, соответствующим младенческой смертности. Кривая типа III – вогнутая – характерна для рыб, выметывающих многочисленную икру, морских донных беспозвоночных с планктонной личинкой, паразитов (для паразитов даже говорят о «законе большого числа яиц» ). Кривая типа II – линейная (в полулогарифмическом масштабе!), или диагональная. Ее считают характерной для птиц. Фактически это просто промежуточный случай (между типом I и типом II). Однако он очень важен теоретически, поскольку эта кривая характеризует безвыборочную (неизбирательную, не зависящую от возраста) элиминацию. Кривая этого типа используется во многих экологических моделях. Кроме того, она нередко встречается на практике.
Кривая типа I – выпуклая – характерна для человека и для большинства млекопитающих. Однако обратите внимание, что для человека и тем более для млекопитающих, живущих в дикой природе, в отличие от идеального типа I начальный этап кривой выживания характеризуется заметным спадом, соответствующим младенческой смертности. Кривая типа III – вогнутая – характерна для рыб, выметывающих многочисленную икру, морских донных беспозвоночных с планктонной личинкой, паразитов (для паразитов даже говорят о «законе большого числа яиц» ). Кривая типа II – линейная (в полулогарифмическом масштабе!), или диагональная. Ее считают характерной для птиц. Фактически это просто промежуточный случай (между типом I и типом II). Однако он очень важен теоретически, поскольку эта кривая характеризует безвыборочную (неизбирательную, не зависящую от возраста) элиминацию. Кривая этого типа используется во многих экологических моделях. Кроме того, она нередко встречается на практике.
 Возрастная когорта (угол 45 о т. к. она вымирает за время равное макс продолжительности жизни) X(τ, t) численность t (время) (максимальная продолжительность жизни) τ (продолжительность жизни) Мгновенная развертка возрастного состава популяции
Возрастная когорта (угол 45 о т. к. она вымирает за время равное макс продолжительности жизни) X(τ, t) численность t (время) (максимальная продолжительность жизни) τ (продолжительность жизни) Мгновенная развертка возрастного состава популяции
 Выяснилось, что в течении первых 70 суток жизни смертность макрели Scomber scombrus составила 99. 9994 % (100% - число отложенных яиц) Из миллиона икринок получалось 6 личинок, достигших длины 25 -39 мм
Выяснилось, что в течении первых 70 суток жизни смертность макрели Scomber scombrus составила 99. 9994 % (100% - число отложенных яиц) Из миллиона икринок получалось 6 личинок, достигших длины 25 -39 мм
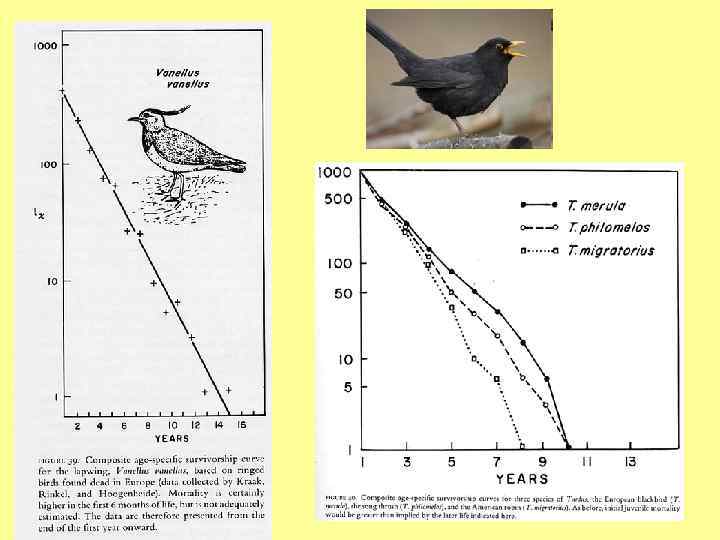
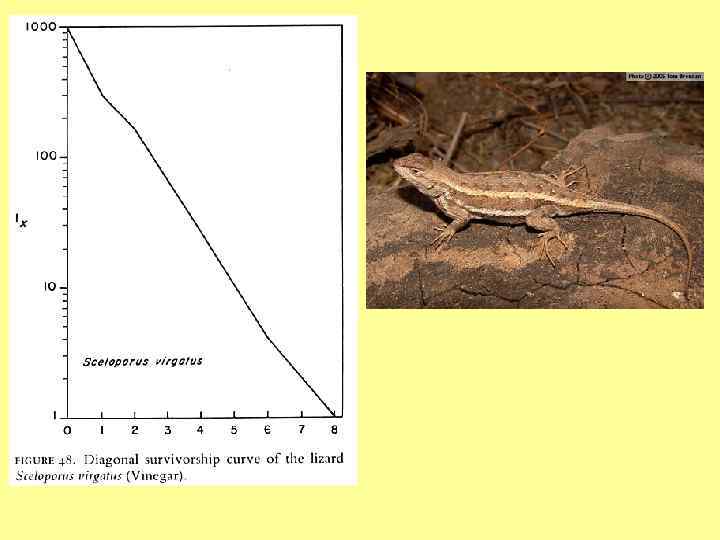
") Диагональные lx кривые (смертность, не зависящая от возраста)
Диагональные lx кривые (смертность, не зависящая от возраста)
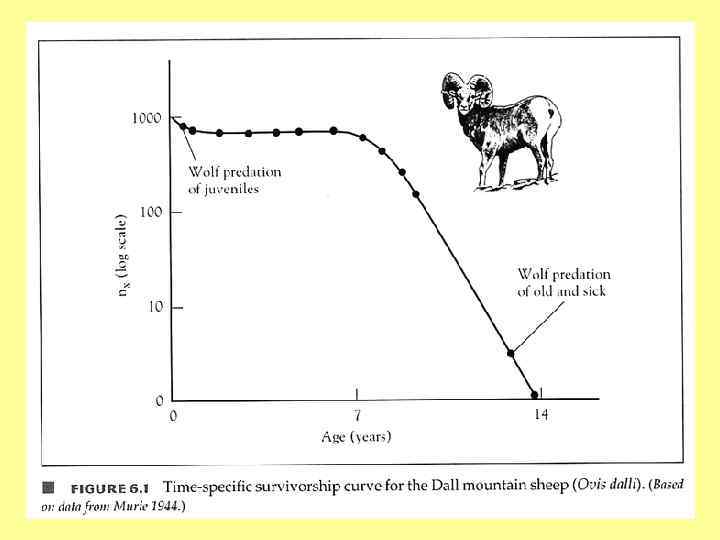
 http: //www. alaskanalpinetreks. com/Image. Gallery/Wildlife_3/Da ll. Sheep. Ram. Standing. On.") Баран Далла (Ovis dalli) http: //www. alaskanalpinetreks. com/Image. Gallery/Wildlife_3/Da ll. Sheep. Ram. Standing. On. Rock. html Из лекций А. М. Гилярова Adolph Murie (1899 – 1974)
Баран Далла (Ovis dalli) http: //www. alaskanalpinetreks. com/Image. Gallery/Wildlife_3/Da ll. Sheep. Ram. Standing. On. Rock. html Из лекций А. М. Гилярова Adolph Murie (1899 – 1974)
 Обложка классической работы Адольфа Мури, в которой он привел данные по смертности барана Далла в разных возрастах, послужившие основой для построения кривой выживания В работе Мури в главе 3 приводятся материалы по повозрастной смертности барана Далла (возраст умерших животных определялся по годовым кольцам прироста на поверхности рога; самые ранние кольца находятся на дистальном конце рога). Всего имеются данные по 608 черепам, однако только у 464 из них сохранились рога. У остальных 144 экземпляров возраст мог быть определен лишь приблизительно (по стертости зубов). Эти звери относились к старшим возрастам (≥ 9 лет) и для упрощения расчетов в Книга полностью выложена на сайте последующей таблице не учтены. (в открытом доступе): http: //www. nps. gov/history/online_books/fauna 5/fauna. htm
Обложка классической работы Адольфа Мури, в которой он привел данные по смертности барана Далла в разных возрастах, послужившие основой для построения кривой выживания В работе Мури в главе 3 приводятся материалы по повозрастной смертности барана Далла (возраст умерших животных определялся по годовым кольцам прироста на поверхности рога; самые ранние кольца находятся на дистальном конце рога). Всего имеются данные по 608 черепам, однако только у 464 из них сохранились рога. У остальных 144 экземпляров возраст мог быть определен лишь приблизительно (по стертости зубов). Эти звери относились к старшим возрастам (≥ 9 лет) и для упрощения расчетов в Книга полностью выложена на сайте последующей таблице не учтены. (в открытом доступе): http: //www. nps. gov/history/online_books/fauna 5/fauna. htm
 http: //scienceblogs. com/tetrapodzoology/2007/06/return_ to_the_world_of_sheep. php http: //www. wildnatureimages. com/Dall's. Sheep. Pictures. htm
http: //scienceblogs. com/tetrapodzoology/2007/06/return_ to_the_world_of_sheep. php http: //www. wildnatureimages. com/Dall's. Sheep. Pictures. htm
 (построена по материалам анализа 464 черепов)") Когортная таблица выживания для барана Далла (Ovis dalli) (построена по материалам анализа 464 черепов) x, годы mx nx nx(1000) lx dx=qx Lx Tx ex 0+ 33 464 1000 1. 000 0. 071 0. 964 7. 117 1+ 88 431 929 0. 204 0. 834 6. 153 6. 624 2+ 7 343 739 0. 020 0. 732 5. 319 7. 195 3+ 8 336 724 0. 024 0. 716 4. 587 6. 335 4+ 7 328 707 0. 021 0. 699 3. 872 5. 477 5+ 18 321 692 0. 056 0. 672 3. 172 4. 586 6+ 28 303 653 0. 092 0. 623 2. 500 3. 828 7+ 29 275 593 0. 105 0. 561 1. 877 3. 167 8+ 42 246 530 0. 171 0. 485 1. 316 2. 482 9+ 47 204 440 0. 230 0. 389 0. 831 1. 890 10+ 67 157 338 0. 427 0. 266 0. 442 1. 306 11+ 56 90 194 0. 622 0. 134 0. 176 0. 906 12+ 32 34 73 0. 073 0. 941 0. 039 0. 042 0. 574 13+ 1 2 4 0. 004 0. 500 0. 003 0. 750 14+ 1 1 2 0. 002 1. 000
Когортная таблица выживания для барана Далла (Ovis dalli) (построена по материалам анализа 464 черепов) x, годы mx nx nx(1000) lx dx=qx Lx Tx ex 0+ 33 464 1000 1. 000 0. 071 0. 964 7. 117 1+ 88 431 929 0. 204 0. 834 6. 153 6. 624 2+ 7 343 739 0. 020 0. 732 5. 319 7. 195 3+ 8 336 724 0. 024 0. 716 4. 587 6. 335 4+ 7 328 707 0. 021 0. 699 3. 872 5. 477 5+ 18 321 692 0. 056 0. 672 3. 172 4. 586 6+ 28 303 653 0. 092 0. 623 2. 500 3. 828 7+ 29 275 593 0. 105 0. 561 1. 877 3. 167 8+ 42 246 530 0. 171 0. 485 1. 316 2. 482 9+ 47 204 440 0. 230 0. 389 0. 831 1. 890 10+ 67 157 338 0. 427 0. 266 0. 442 1. 306 11+ 56 90 194 0. 622 0. 134 0. 176 0. 906 12+ 32 34 73 0. 073 0. 941 0. 039 0. 042 0. 574 13+ 1 2 4 0. 004 0. 500 0. 003 0. 750 14+ 1 1 2 0. 002 1. 000
 Величины, входящие в таблицу выживания x – возраст, точнее, возрастной интервал или класс; 0+ означает интервал от 0 до 1 года (ягнята-сеголетки), 1+ означает интервал от 1 до 2 лет (годовики) и т. д. mx – число погибших в интервале от 0 до 1 года (x = 0+), от 1 до 2 лет (x = 1+) и т. д. В случае барана Далла это данные полевых учетов, которые служат основой для всех последующих расчетов nx – число выживших на начало возрастного интервала x+, или восстановленная численность когорты Вы легко можете проверить эти уравнения по таблице выживания на предыдущем слайде! и т. д.
Величины, входящие в таблицу выживания x – возраст, точнее, возрастной интервал или класс; 0+ означает интервал от 0 до 1 года (ягнята-сеголетки), 1+ означает интервал от 1 до 2 лет (годовики) и т. д. mx – число погибших в интервале от 0 до 1 года (x = 0+), от 1 до 2 лет (x = 1+) и т. д. В случае барана Далла это данные полевых учетов, которые служат основой для всех последующих расчетов nx – число выживших на начало возрастного интервала x+, или восстановленная численность когорты Вы легко можете проверить эти уравнения по таблице выживания на предыдущем слайде! и т. д.
 – то же, что nx, только из расчета") Величины, входящие в таблицу выживания nx(1000) – то же, что nx, только из расчета на начальную численность когорты n 0 = 1000 особей lx = nx/n 0 – доля особей, доживших до возраста x; график этой функции есть кривая выживания dx (в других обозначениях qx) – удельная смертность особей в когорте, или удельная повозрастная смертность
Величины, входящие в таблицу выживания nx(1000) – то же, что nx, только из расчета на начальную численность когорты n 0 = 1000 особей lx = nx/n 0 – доля особей, доживших до возраста x; график этой функции есть кривая выживания dx (в других обозначениях qx) – удельная смертность особей в когорте, или удельная повозрастная смертность
 ex – средняя ( «ожидаемая» ) продолжительность жизни") Величины, входящие в таблицу выживания (3) ex – средняя ( «ожидаемая» ) продолжительность жизни после достижения возраста x; например, e 0 - средняя продолжительность жизни новорожденных особей. Средняя продолжительность жизни новорожденных особей равна площади под кривой выживания Lx и Tx – вспомогательные величины для определения ex, то есть для расчета площади под кривой выживания Из лекций Л. В. Полищука и т. д.
Величины, входящие в таблицу выживания (3) ex – средняя ( «ожидаемая» ) продолжительность жизни после достижения возраста x; например, e 0 - средняя продолжительность жизни новорожденных особей. Средняя продолжительность жизни новорожденных особей равна площади под кривой выживания Lx и Tx – вспомогательные величины для определения ex, то есть для расчета площади под кривой выживания Из лекций Л. В. Полищука и т. д.
 1.") Графики, которые можно построить на основе таблицы выживания Кривая выживания (в арифметическом масштабе) 1. 000 l (x ) 0. 800 0. 600 Lx – это площадь 0. 400 0. 200 0. 000 0 2 4 6 8 10 Возраст, годы 12 14 16 указанного прямоугольника. Сумма Lx дает площадь под кривой выживания, то есть T 0 = e 0. Обратите внимание, что e 0 = 7. 1 лет из таблицы выживания численно примерно соответствует площади под кривой lx (фигуру под кривой можно приблизительно считать прямоугольным треугольником)
Графики, которые можно построить на основе таблицы выживания Кривая выживания (в арифметическом масштабе) 1. 000 l (x ) 0. 800 0. 600 Lx – это площадь 0. 400 0. 200 0. 000 0 2 4 6 8 10 Возраст, годы 12 14 16 указанного прямоугольника. Сумма Lx дает площадь под кривой выживания, то есть T 0 = e 0. Обратите внимание, что e 0 = 7. 1 лет из таблицы выживания численно примерно соответствует площади под кривой lx (фигуру под кривой можно приблизительно считать прямоугольным треугольником)
") Бараны Размерность площади под кривой выживания: «барано-годы» (в общем случае «особи × возраст» ) Площадь под кривой выживания Барано-годы Годы Из лекций А. М. Гилярова (с
Бараны Размерность площади под кривой выживания: «барано-годы» (в общем случае «особи × возраст» ) Площадь под кривой выживания Барано-годы Годы Из лекций А. М. Гилярова (с
 Удельная смертность") Графики, которые можно построить на основе таблицы выживания (на примере барана Далла) Удельная смертность особей в когорте (удельная повозрастная смертность) 1. 000 d(x) Хищничество волков на старых и больных особях 0. 800 0. 600 Хищничеств о волков на годовиках ? 0. 400 0. 200 0. 000 0 5 Из лекций Л. В. Полищука 10 15
Графики, которые можно построить на основе таблицы выживания (на примере барана Далла) Удельная смертность особей в когорте (удельная повозрастная смертность) 1. 000 d(x) Хищничество волков на старых и больных особях 0. 800 0. 600 Хищничеств о волков на годовиках ? 0. 400 0. 200 0. 000 0 5 Из лекций Л. В. Полищука 10 15
 Гипотетическая равновесная популяция с 6 ВК. mx –") ПОЛНАЯ ДЕМОГРАФИЧЕСКАЯ ТАБЛИЦА (по Пианка, 1980) Гипотетическая равновесная популяция с 6 ВК. mx – число потомков одной особи за возрастной период х, lx – доля особей начальной когорты доживших до возраста х, Сумма mx по возрастам х – валовая рождаемость (М), Сумма произведений mx на lx по возрастам х – чистая скорость роста R 0, Сумма произведений mx на lx и на возраст х, отнесенное к R 0 - среднее время генерации (Т), сумма выживаемостей lx всех возрастов начиная с возраста х, отнесенная к lx- ожидаемая продолжительность жизни возраста ex Возраст (Х) годы l. X m. X l X m. X x l X m. X e. X 0 1. 0. 0. 3. 4 1 . 8 . 2 . 16 3. 2 . 6 . 3 . 18 . 36 2. 67 3 . 4 1. 2 2. 5 4 . 6 . 24 . 96 1. 5 5 . 2 . 1 . 02 . 1 1. 6 0. 0 0. 0. СУММЫ: 2. 2 (М) 1. 00 (R 0) 2. 78 (Т)
ПОЛНАЯ ДЕМОГРАФИЧЕСКАЯ ТАБЛИЦА (по Пианка, 1980) Гипотетическая равновесная популяция с 6 ВК. mx – число потомков одной особи за возрастной период х, lx – доля особей начальной когорты доживших до возраста х, Сумма mx по возрастам х – валовая рождаемость (М), Сумма произведений mx на lx по возрастам х – чистая скорость роста R 0, Сумма произведений mx на lx и на возраст х, отнесенное к R 0 - среднее время генерации (Т), сумма выживаемостей lx всех возрастов начиная с возраста х, отнесенная к lx- ожидаемая продолжительность жизни возраста ex Возраст (Х) годы l. X m. X l X m. X x l X m. X e. X 0 1. 0. 0. 3. 4 1 . 8 . 2 . 16 3. 2 . 6 . 3 . 18 . 36 2. 67 3 . 4 1. 2 2. 5 4 . 6 . 24 . 96 1. 5 5 . 2 . 1 . 02 . 1 1. 6 0. 0 0. 0. СУММЫ: 2. 2 (М) 1. 00 (R 0) 2. 78 (Т)
 яйца взрослые нимфы mx Т lx 0 0 10 R 0 = 30, 9 20 30 l x mx 40 50 60 дни Данные по динамике популяционного роста человеческой вши
яйца взрослые нимфы mx Т lx 0 0 10 R 0 = 30, 9 20 30 l x mx 40 50 60 дни Данные по динамике популяционного роста человеческой вши
 Homo sapiens Древний Рим данные в основном за первые четыре столетия Нашей Эры
Homo sapiens Древний Рим данные в основном за первые четыре столетия Нашей Эры
 Женщины Мужчины 40 100 лет") 1000 Число выживших из 1000 рожденных (логарифми ческая шкала) Женщины Мужчины 40 100 лет Возраст
1000 Число выживших из 1000 рожденных (логарифми ческая шкала) Женщины Мужчины 40 100 лет Возраст
 1000 Число выживших из 1000 рожденных Девочки Мальчики 985 10 дней 20 дней Возраст
1000 Число выживших из 1000 рожденных Девочки Мальчики 985 10 дней 20 дней Возраст
 Возрастная когорта (угол 45 о т. к. она вымирает за время равное макс продолжительности жизни) X(τ, t) численность t (время) (максимальная продолжительность жизни) τ (продолжительность жизни) Мгновенная развертка возрастного состава популяции
Возрастная когорта (угол 45 о т. к. она вымирает за время равное макс продолжительности жизни) X(τ, t) численность t (время) (максимальная продолжительность жизни) τ (продолжительность жизни) Мгновенная развертка возрастного состава популяции
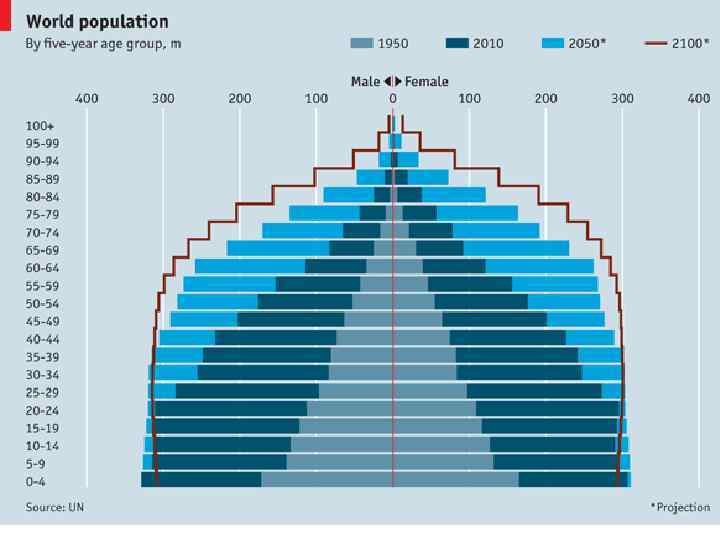
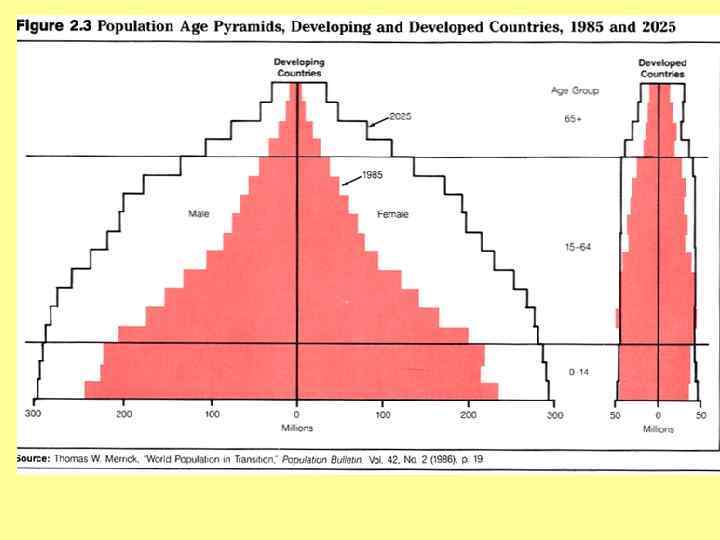
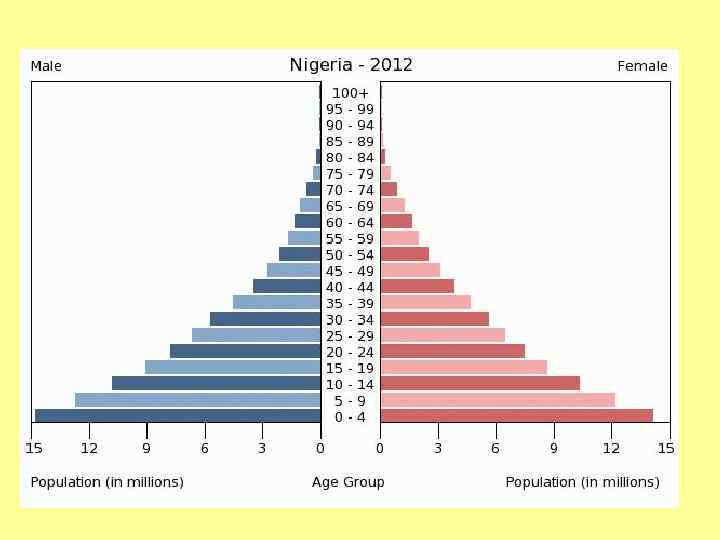
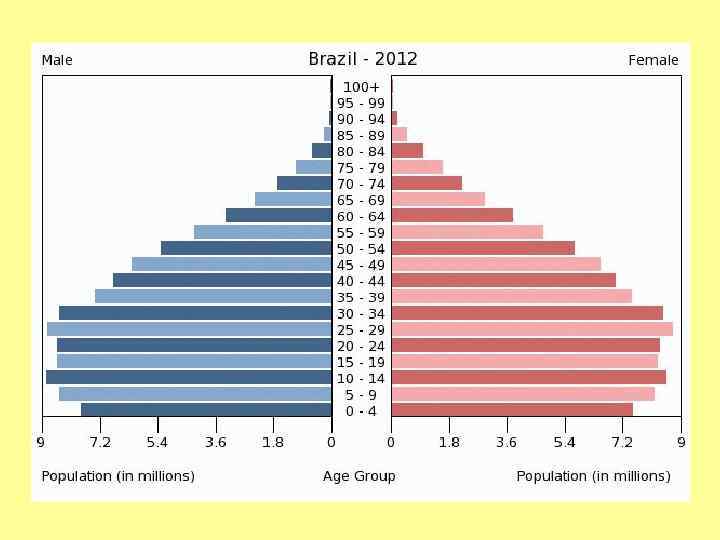
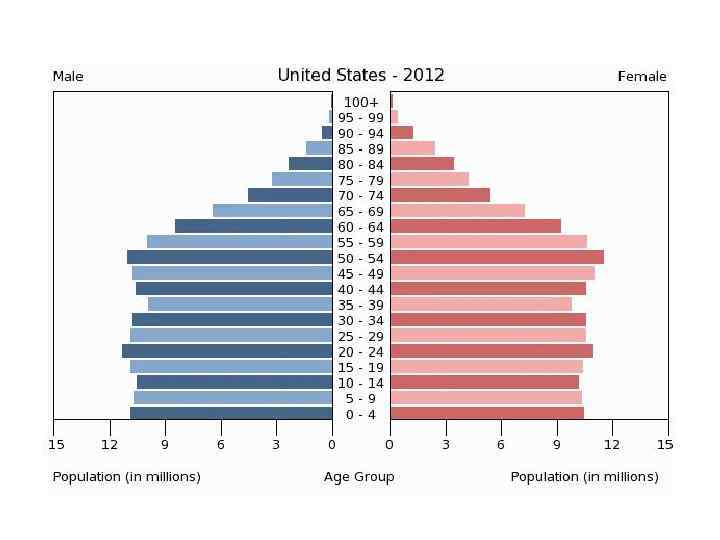
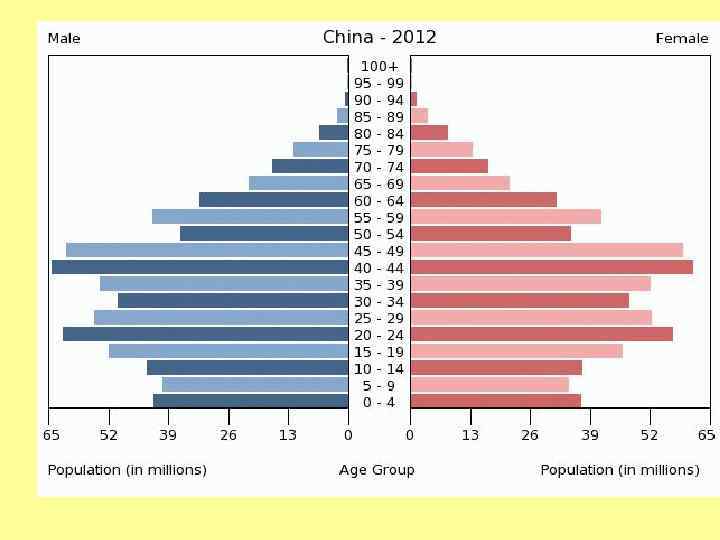
 Возрастное распределение Homo sapiens, 1970 г. а – Мексика; б – США; в - Швеция
Возрастное распределение Homo sapiens, 1970 г. а – Мексика; б – США; в - Швеция


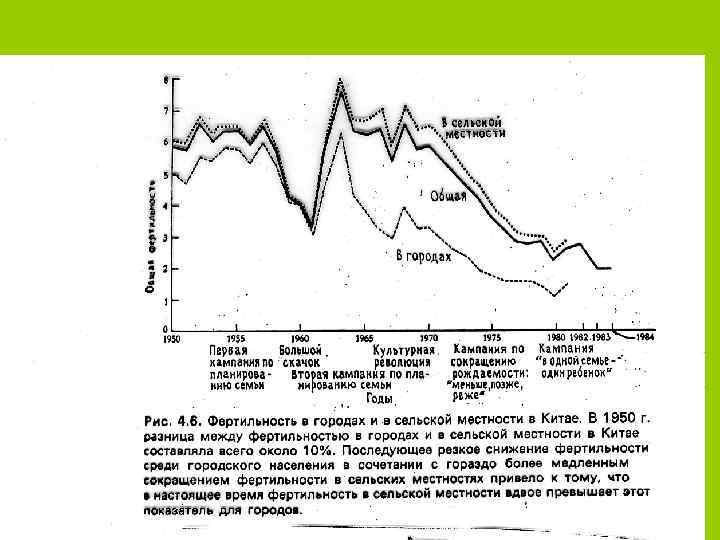

 Возрастное распределение народонаселения Франции на 1 января 1967 г.
Возрастное распределение народонаселения Франции на 1 января 1967 г.
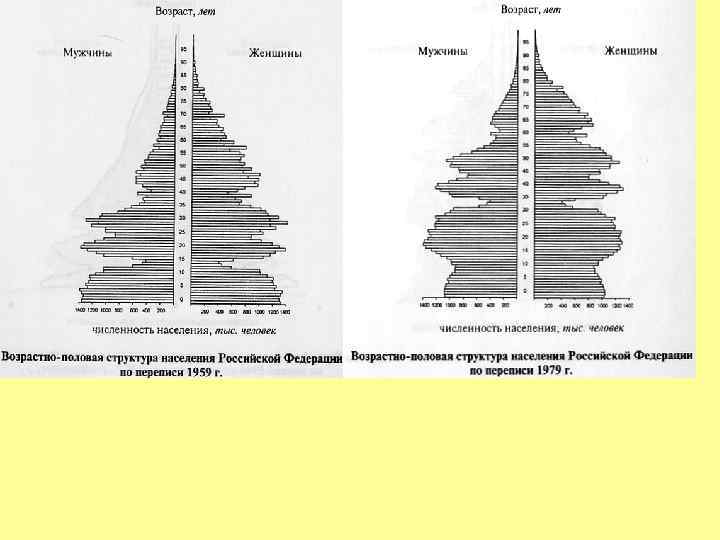
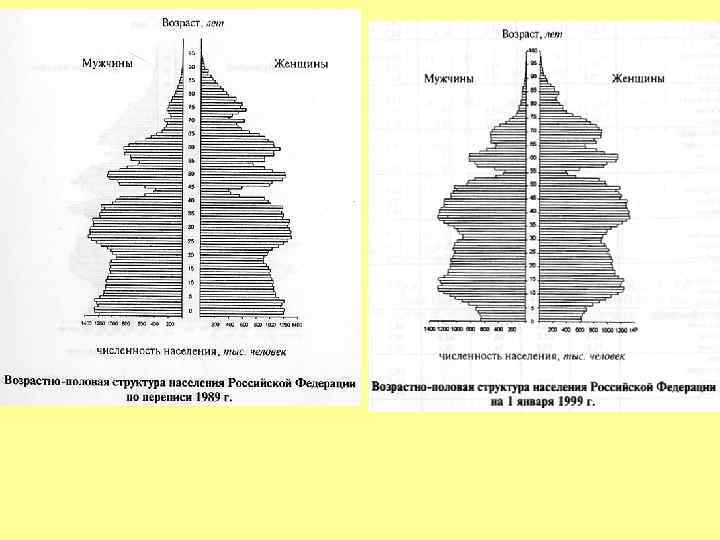
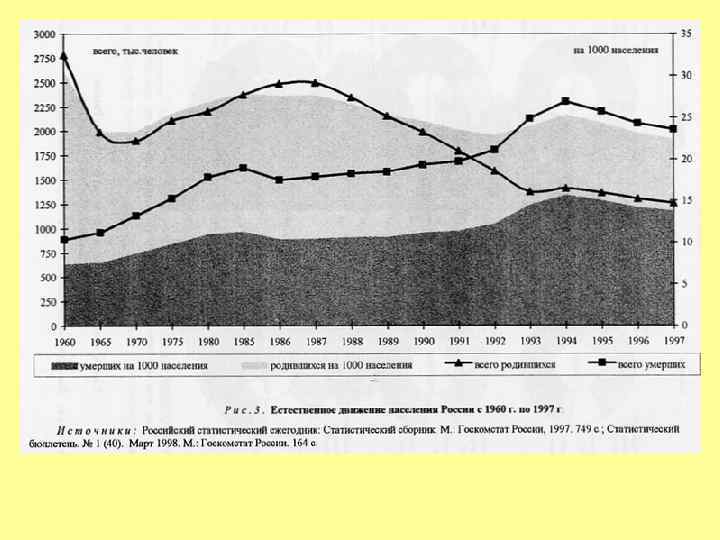
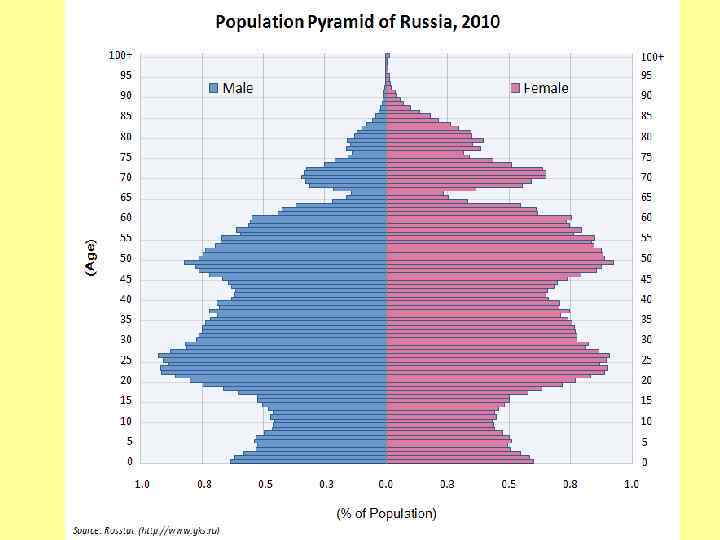
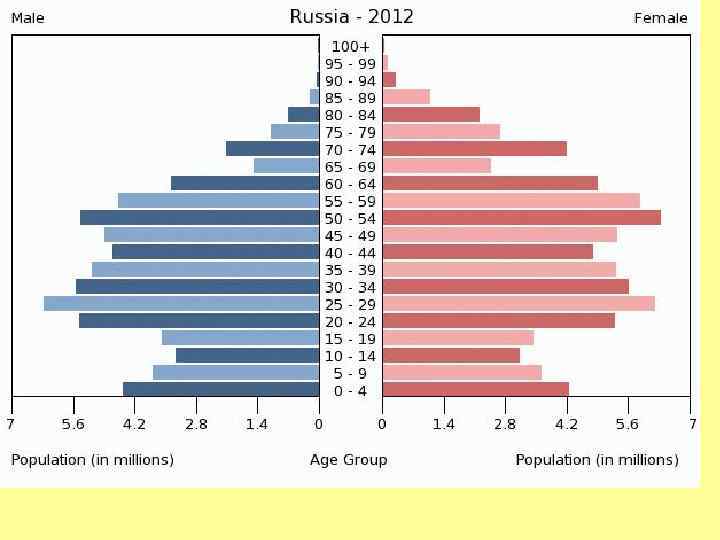
 Возрастное распределение народонаселения России на 1 января 1997 г.
Возрастное распределение народонаселения России на 1 января 1997 г.
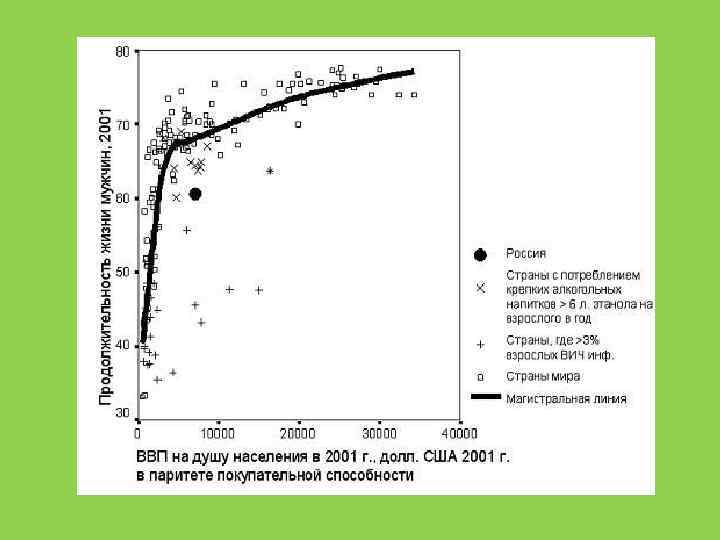
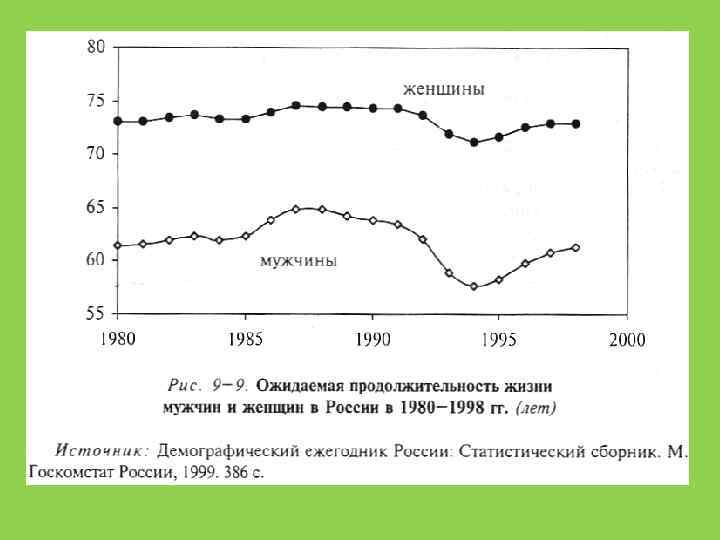
 5. 5 5 4 3. 5 3") Плодовитость человечества (кол-во детей за жизнь матери) 5. 5 5 4 3. 5 3 2. 5 2 1. 5 1 1940 1950 1960 1970 1980 1990 2000 2010 2020 2030 2040
Плодовитость человечества (кол-во детей за жизнь матери) 5. 5 5 4 3. 5 3 2. 5 2 1. 5 1 1940 1950 1960 1970 1980 1990 2000 2010 2020 2030 2040
") Скорость прироста населения Земли (% в год)
Скорость прироста населения Земли (% в год)