взаимодействие вируса с клеткой.ppt
- Количество слайдов: 29



ВЗАИМОДЕЙСТВИЕ ВИРУСА С КЛЕТКОЙ ГБОУ ВПО РНИМУ им. Н. И. Пирогова Кафедра микробиологии и вирусологии Заседание СНК, февраль 2012 г. Руководитель: Лордкипанидзе А. Е. Доклад студ. II курса 208 гр. Татьяны Кручининой

• Процесс размножения вируса – происходит только внутриклеточно. Поэтому определяют жизненный цикл вирусов, который сводится к взаимодействию их с клеткой-мишенью – от момента адсорбции на мембране клетки до выхода вновь синтезированных вирионов (внеклеточных вирусов) из клетки.

Адсорбция на поверхности клетки • Для каждого вируса на мембране клеток существуют специфические рецепторы, с которыми вирусы связываются с помощью своих рецепторов (органотропность). Поскольку и у клеток, и у вирусов рецепторы разнообразны, то на одних и тех же клетках могут адсорбироваться различные вирусы.

• Вирион гриппа. Гемагглютинин – это тримерный гликопротеин вирусной оболочки. Каждый мономер – состоит из двух сегментов. Гемагглютинин является рецептором адгезии, связывающийся с рецептором клеток -мишеней (мукопептидом).

• Надо сказать, что поверхностные антигены вируса гриппа, в частности - гемагглютинин, меняются в связи с мутациями в геноме вирусных белков. Обычно антигенные структуры вирусов служат мишенями для антител и Т-клеток. Но при изменении структуры гемагглютинина, ранее выработанные антитела оказываются неэффективными. Антигенная вариабельность характерна также для некоторых других вирусов (ВИЧ и ящура).

Проникновение вируса в клетку Суперкапсид – биологическая мембрана (билипидный слой и выступающие над поверхностью вириона белки). [Для вируса гепатита В - характерен второй путь]. Вирус гепатита В • 1. Посредством слияния суперкапсида (второй оболочки) вируса с мембраной клетки. При этом происходит высвобождение нуклеокапсида (нуклеиновой кислоты и белковой оболочки) в цитоплазму.

Посредством слияния суперкапсида с мембраной клетки – проникают вирусы, обладающие F-белками или белками, выполняющими их функции (гемагглютинин вируса гриппа).
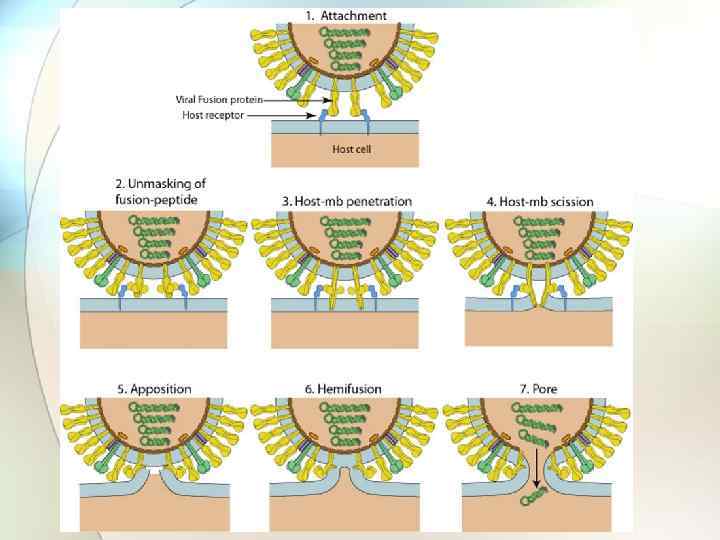
. В этом случае вирус связывается со специфическими рецепторами")
2. С помощью рецепторопосредованного эндоцитоза (пиноцитоза). В этом случае вирус связывается со специфическими рецепторами в области «окаймленной ямки» (область мембраны окаймлена щетинками - белком Проникновение клатрином). Она вируса гепатита С впячивается внутрь и превращается в окаймленный пузырек, который быстро сливается с эндосомой и далее – с лизосомой. Подкисленное содержимое активирует вирусные белки и суперкапсид сливается с мембраной лизосомы. Это приводит к выходу нуклеокапсида в цитозоль клетки, где далее высвобождается геномная нуклеиновая кислота.

• Если вирус простой, то его проникновение путем эндоцитоза заканчивается лизисом мембраны лизосомы или нарушением ее проницаемости (также – в слабокислой среде).
 двунитевую ДНК (расплетение")
ВНУТРИКЛЕТОЧНОЕ РАЗМНОЖЕНИЕ ВИРУСОВ • Механизм репродукции отличается у вирусов, имеющих: 1) двунитевую ДНК (расплетение нитей ДНК и комплементарное достраивание новых) происходит в ядре – у герпесвирусов, аденовирусов, папиллома- и полиомавирусов; и в цитоплазме – у поксвирусов. У гепандовирусов – одна нить ДНК – неполная (плюс-нить). Она достраивается при попадании вируса в клетку, когда созревающий геном – проникает в ядро.
 и РНКпрегеном")
Здесь ДНК-зависимая РНК -полимераза синтезирует разные и. РНК (для синтеза вирусных белков) и РНКпрегеном (матрица для репликации генома вируса). Далее и. РНК перемещаются в цитоплазму и транслируются с образованием белков вируса. Белки сердцевины вируса собираются вокруг прегенома, на матрице которого синтезируется минус-нить ДНК, на которой образуется плюснить ДНК. Оболочка вириона формируется на НВsсодержащих мембранах ЭПС и аппарата Гольджи. Выход – экзоцитозом.
 однонитевую ДНК (использование клеточной ДНК-полимеразы для создания двунитевого вирусного генома) – в ядре")
2) однонитевую ДНК (использование клеточной ДНК-полимеразы для создания двунитевого вирусного генома) – в ядре у парвовирусов. • На исходной вирусной ДНК (плюс-нить) комплементарно синтезируется минус-нить ДНК, служащая матрицей в синтезе плюснити ДНК для новых поколений вирусов. Параллельно синтезируется и. РНК, транслируются вирусные белки, которые возвращаются в ядро, где собираются вирионы.
 и клетке (В). • На фото")
Электронная микрофотография парвовируса В 19 в сыворотке (А) и клетке (В). • На фото В - изображен предшественник эритроцита, инфицированный парвовирусом in vitro.
 плюс-однонитевую РНК (после эндоцитоза освобождается геномная плюс. РНК, которая связывается с")
• 3) плюс-однонитевую РНК (после эндоцитоза освобождается геномная плюс. РНК, которая связывается с рибосомами для синтеза 4 -х структурных белков, один из которых - РНК-полимераза – транскрибирует минус-нить РНК, т. е. матрицу, по плюс-РНК) – у пикорнавирусов, флавивирусов, тогавирусов. • По матрице минус-нити – синтезируются: полная плюс-нить геномной РНК и неполная нить, кодирующая С-белок капсида и гликопротеины оболочки.

Пикорнавирус: плюс-нить РНК – выполняет функцию и. РНК. На матрице минус-нити – синтезируются плюс-нити генома и белки оболочки.
 минус-однонитевую РНК (синтез плюснитей с участием уже имеющейся в составе РНК-полимеразы) – у")
4) минус-однонитевую РНК (синтез плюснитей с участием уже имеющейся в составе РНК-полимеразы) – у рабдовирусов, парамиксовирусов, ортомиксовирусов. Минус-нить РНК – трансформируется РНКзависимой РНК-полимеразой в неполные и полные плюс-нити РНК. По неполным идет синтез вирусных белков. Полные – являются промежуточной матрицей для синтеза минус-нитей РНК потомства.

• Вирус гриппа, в отличие от других негативных однонитевых вирусов, нуждается в м. РНК, и поэтому синтез вирусной РНК идет в ядре.
 двунитевую РНК (процесс репродукции, происходящий в цитоплазме клеток, сходен с минусоднонитевыми")
• 5) двунитевую РНК (процесс репродукции, происходящий в цитоплазме клеток, сходен с минусоднонитевыми РНКвирусами: по минуснити синтезируется плюс-нить, являющаяся и. РНК, а также – матрицей для минуснитей) – у реовирусов и ротавирусов.
 идентичные плюс-нитевые РНК – обратнотранскрибирующиеся (ретровирусы), например, ВИЧ-вирионы через поверхностный антиген")
• 6) идентичные плюс-нитевые РНК – обратнотранскрибирующиеся (ретровирусы), например, ВИЧ-вирионы через поверхностный антиген gp 120 связываются с CD 4+ и рецепторами для хемокинов. Проникнув внутрь (слияние оболочки ВИЧ с плазмалеммой клетки), - встраивается в геном клетки-хозяина (латентное состояние). Под действием стимула (эффект цитокинов) начинается вирусная транскрипция и активация.

• При проникновении в клетку – освобождается геномная РНК и обратная транскриптаза вируса, которая синтезирует (на матрице геномной РНК) комплементарную минус-нить ДНК (линейная к. ДНК). С последней копируется плюснить к. ДНК и образуется двойная кольцевая к. ДНК, интегрируемая с ДНК клетки. При активации – с ДНКпровируса синтезируются геномная РНК и и. РНК, обеспечивающие синтез компонентов и сборку вирионов, которые отпочковываются с использованием модифицированной мембраны клетки.

• Тип взаимодействия вируса с клеткой, при котором образуются новые вирионы, называется «продуктивный» . • Если инфекционный процесс прерывается в клетке, то такой тип называется «абортивный» . • Если внедрение вируса приводит к встраиванию его ДНК в хромосому клетки, то тип взаимодействия называется «интегративный» .

• При взаимодействии с клеткой, все вирусы стараются уклониться от действия иммунной системы. Так, большинство из них разработало стратигию подавления системы интерферонов (т. к. INF – индуцируют молекулы МНС и запускают другие механизмы противовирусного действия); также стратегию, нарушающую хемокиновую сеть. Большинство ДНК-содержащих вирусов – используют стратегию контроля за экспрессией молекул МНС. Молекулы главного комплекса гистосовместимости класса I (присутствующие на всех ядросодержащих клетках организма) – презентируют антигены цитотоксическим Тклеткам.

• Презентация антигена: процессинг – обработка антигена и превращение его в форму, необходимую для распознавания лимфоцитами. Протеасомы – органеллы, которые вызывают распад клеточных белков, намеченных для расщепления посредством присоединения убиквитина в качестве метки для деградации. ТАР – это представители АТФ-связывающего суперсемейства транспортных белков, локализованных на мембране эндоплазматического ретикулума. Молекулы МНС класса I синтезируются в эндоплазматическом ретикулуме и проходят через аппарат Гольджи в комплексе с антигенным пептидом эндогенного происхождения.

• Молекулы МНС класса II образуют комплексы с пептидами экзогенного происхождения. Синтезируются в эндоплазматическом ретикулуме в комплексе с полипептидом (инвариантной цепью – Ii), проходят через аппарат Гольджи и соединяются в эндосомальном компартменте с содержащимися там пептидами. • [Молекулы МНС класса II презентируют антигены Тхелперам. ] • В то же время ответ, направленный против экзогенного антигена, может проходить и с участием MHC класса I.

• Вирусы стараются нарушить экспрессию молекул МНС на разных стадиях (блокадой поступления пептидов в ЭПР, нарушением транспорта молекул МНС, их транскрипции и т. д. ). • Вирусы, способные инфицировать клетки иммунной системы (ВИЧ), непосредственно инфицируют лимфоциты и макрофаги. • Согласно последним данным, в организме существует врожденный механизм защиты от репликации / транскрипции вирусных РНК. В клетках эукариот образуется ds. РНК (двуспиральная РНК double-stranded) – как часть процесса, который контролирует активность м. РНК при помощи ингибирующих РНК (si. РНК), что приводит к распознаванию и деградации ds. РНК.
 с")
ds. РНК нарезаются РНКазой Dicer на фрагменты длиной 21 -25 нуклеотида (si. РНК) с выступающими на два нуклеотида 3`-концами, после чего одна из цепей входит в состав RISC и направляет разрезание гомологичных молекул РНК. В составе RISC присутствуют si. РНК, соответствующие как плюс-, так и минусцепям ds. РНК.

СПАСИБО ЗА ВНИМАНИЕ!

Литература: • А. И. Коротяев, С. А. Бабич «Медицинская микробиология, иммунология и вирусология» , Санкт-Петербург, Спец. Лит, 2008 г. • А. С. Быков, А. А. Воробьев, В. В. Зверев «Атлас по медицинской микробиологии, вирусологии и иммунологии» , Медицинское Информационное Агентство, Москва, 2008 г. • Д. Мейл, Дж. Бростофф, Д. Б. Рот, А. Ройтт «Иммунология» , Москва, Издательство «Логосфера» , 2007 г. • Ю. А. Макарова, Д. А. Крамеров «Некодирующие РНК» , Биохимия, 2007 г. , том 72 вып. 11
взаимодействие вируса с клеткой.ppt