Лекция 12. Вторичные метаболиты.ppt
- Количество слайдов: 70



ВТОРИЧНЫЕ МЕТАБОЛИТЫ ГРИБОВ: ФУНКЦИИ В ПРИРОДЕ

Вторичные метаболиты и вторичный метаболизм Термины введены биохимиком А. Косселем в 1891 г. Первичные метаболиты присутствуют в каждой растительной клетке, способной к делению. Вторичные метаболиты присутствуют в клетке случайно и не являются необходимыми для ее жизни. Вторичные – второстепенные, случайные.
 и развиты К.")
Современные представления о вторичном метаболизме впервые сформулированы Ф. Чапеком (Czapek, 1921) и развиты К. Пэхом (Päch, 1950). Первичный метаболизм универсален. Он практически не различается у разных организмов и не видоспецифичен. Вторичный метаболизм не универсален. Он является производным от первичного метаболизма.

Вторичные метаболиты: 1. Индивидуальны и разнообразны, часто образуются у конкретных таксономических групп грибов; Среди грибов β-лактамные антибиотики образуют только эвроциомицеты (Penicillium spp. И др. ) и гипокрейные (Enericellopsis spp. , Acremonium spp, ) Эта группа соединений известна также у прокариот

2. Синтезируются из небольшого набора исходных соединений и в ограниченном числе биогенетических путей;

Несколько путей биогенеза вторичных метаболитов: Ацетил-Ко. А 1. Синтез поликетидов и жирных кислот. 2. Синтез терпеноидов. Не связанные с ацетатом 3. Нерибосомальный синтез пептидов. 4. Через шикимовую кислоту. 5. Непосредственно из глюкозы
;")
3. Имеют относительно низкую молекулярную массу(обычно не более 2. 0 КДа);

4. Образуются обычно в идиофазе, после периода активного роста гриба

4. Образуются обычно в идиофазе, после периода активного роста гриба

5. Гены для синтеза многих вторичных метаболитов образуют кластеры. Экспрессия кластеров индуцируется одним или немногими глобальными регуляторами. Некоторые из таких глобальных регуляторов связаны также с процессами споруляции и элонгации гиф. Экспрессия вторичных метаболитов наблюдается в определенных точках жизненных циклов.

Кластеры генов биосинтеза некоторых биологически активных веществ

6. Часто обладают разнообразной биологической активностью; Биологически активные вещества грибов: 1. Антибиотики (антибактериальные, антифунгальные, противоопухолевые, противовирусные и др. ). 2. Биологически активные вещества, действующие на различные процессы у макроорганизма (иммуномодуляторы, ингибиторы синтеза стеринов и т. п. ). 3. Микотоксины 4. Регуляторы процессов роста и развития продуцента 5. Сигнальные соединения

7. Выполняют экологические и регуляторные функции

Функции вторичных метаболитов в природе Существовала точка зрения , что вторичные метаболиты представляют лабораторные продукты, но сейчас уже не вызывает сомнений, что это соединения, играющие огромную роль в жизни их продуцентов. A. L. DEMAIN S. A. WAKSMAN

A. L. Demain, A. Fang, 2000 1. Биологически активные вторичные метаболиты найдены более чем у 40% мицелиальных грибов. Они известны практически у всех групп организмов – прокариот и эукариот. Альтернативный защитный механизм у организмов, не имеющих хорошо развитой и ммунной системы 2. Образуются продуцентами в природе как в наземных, так и водных экосистемах и участвуют в конкуренции между микроорганизмами, растениями и животными. 3. Возникновение и сохранение мультигенных биосинтетических путей вторичных метаболитов в процессе эволюции, отсутствие в кластерах нефункционирующих генов свидетельствуют о их роли в выживании продуцентов. 4. Кластеризация биосинтетических генов, которая могла сложиться только если продукт представлял селективное преимущество для продуцента. В кластерах отсутствуют не фунционирующие гены. Присутствие в кластерах регуляторных генов 5. Связь биосинтеза вторичных метаболитов с процессами дифференциации и морфогенеза

Основные функции вторичных метаболитов грибов: 1. Средства конкурентной борьбы в экосистемах 2. Орудия нападения при паразитизме и хищничестве 3. Агенты симбиотических ассоциаций (в широком смысле) 4. Защита субстрата от консументов 5. Защита от абиотических и биотических воздействий среды 6. Коммуникация –сигнальные метаболиты при межвидовых взаимодействиях 7. Эффекторы дифференциации 8. Половые феромоны 9. Регуляция популяций продуцента 10. Транспорт и резервы металлов

Биохимическая экология Г. Ф. Гаузе, 1935. Роль биологически активных веществ во взаимодействиях организмов М. Барбье, 1978. Взаимодействия организмов при посредстве химических веществ – химическая биохимия. Коммуникация – сигнальные метаболиты
")
СИГНАЛЫ ВНЕШНЕЙ СРЕДЫ СИГНАЛЫ ГРИБОВ (АВТОИНГИБИТОРЫ, ФЕРОМОНЫ И ДР, )
")
Взаимодействия грибов и бактерий: роль вторичных метаболитов (Wargo, Hogan, 2006)
 Характер действия")
Функции и адаптивная значимость метаболитов грибов Функциональные группы веществ (Whittaker, Feeny, 1971) Характер действия веществ Типы биотических взаимодействий Репелленты +/ Защита от нападения или инфекции. Защита субстрата Паразитизм, хищничество. Конкуренция Супрессанты +- Подавление конкурентов Конкуренция Яды (некротоксины) +Индуктанты +/ Умерщвление жертвы или хозяина Паразитизм, хищничество Модификация роста или физиологических функций другого организма Паразитизм, мутуализм, протокооперация Каунтерактанты +/ Нейтрализация ядов или других химических агентов Паразитизм, конкуренция АЛЛОМОНЫ
 Характер действия веществ Типы биотических взаимодействий")
Продолжение таблицы Функциональные группы веществ (Whittaker, Feeny, 1971) Характер действия веществ Типы биотических взаимодействий Аттрактанты +- Привлечение жертвы или хозяина +- ++ Привлечение опылителей, переносчиков спор и т. д. Паразитизм, хищничество Коменсализм, протокооперация, мутуализм КАЙРОМОНЫ Аттрактанты -+ Сигнализация локации хозяина или жертвы Паразитизм, хищничество Индуктанты +/ -+ Стимуляция развития грибов Мутуализм, паразитизм, хищничество ДЕПРЕССАНТЫ 0 -

Роль в экосистемах Конкуренция с другими организмами. Альтернативный защитный механизм у организмов, не имеющих хорошо развитой и ммунной системы. Они образуются в природе и участвуют в конкуренции между микроорганизмами, растениями и животными. На важную роль такого механизма защиты указывает и кластеризация биосинтетических генов, которая могла сложиться только если продукт представлял селективное преимущество для продуцента. В кластерах отсутствуют не фунционирующие гены. Молчащие гены. Активация скрытых кластеров генов и получение новых ВМ.

Cladonia cristatella образует метаболиты, игибирующие рост симбиотрофов Suillus luteus, Pisolithus tinctorius, Thelephora terrestris, но не влияющие на рост Cenococcum graniforme. В состав активного комплекса входят Dусниновая кислота, а как минорные компоненты дидимовая к-та и ее производные и другие лишайниковые кислоты

Биологический метод регуляции популяции фитопатогенных организмов и насекомых Многочисленные препараты Trichoderma spp. , Gliocladium virens против Pythium ultimum- мутанты с сверхпродукцией антибиотиков более эффективны. Даже в случаях прямого паразитизма агента биометода (как биотрофного, так и некротрофного) установлена роль антибиотиков в этом процессе: подавление Botrytis cinerea Trichoderma harzianum, роль хитиназы, β-1, 3 глюканазы и мембраноактивных (каналообразующих) антибиотиков из группы пептайболов (трихорцианинов)- синергизм действия. Паразитизм Monocillium nordinii на ржавчинных грибах Cronartium связан с образованием антибиотиков монордена и моноциллинов.

Защита от консументов. Грибы - источник пищи для разных групп организмов – животных, грибов, бактерий. Механизмы защиты от мицетофагов.


Грибы как источник пищи для разных групп – животные, грибы, бактерии. Механизмы защиты от мицетофагов Детерренты, или антифиданты в базидиомах базидиальных грибов. Chalciporus piperatus – хальципорин Токсины макромицетов

У некоторых грибов такие защитные соединения находятся в виде неактивных предшественников, но превращаются вс высоко токсические соединения или детерренты при нападении на них и повреждении. Терпеновый диальдегид веллерал – причина трудной доступности Lactarius vellereus для насекомых, млекопитающих и грибов – образуется при повреждении из стеароилвелютинала под действием эстеразы.

Антибиотики с антифунгальной активностьюстробилурины – защищают Strobilurus esculentus и другие виды этого рода от поражения микофильными грибами На основе стробилурина созданы фунгициды, широко применяемые в растениеводстве

Афлатоксины и стеригматоцистин – защита от выедания Сollembola Aspergillus flavus, A. parasiticus Aspergillus nidulans Мутанты Aspergillus nidulans с делецией Lae. A не образуют стеригматоцистин и активно выедаются коллемболами

H. luteovirens Hypomyces lateritius Микофильные грибы из рода Hypomyces образуют вторичные метаболиты, защищающие субстрат от заселения другими организмами (грибами, насекомыми, бактериями) Hypomyces lateritius

Эндофиты злаков из родов Neotyphodium, Epichloe, Balansia образуют 4 класса токсинов – эргоалкалоиды, лолитремы, лолины и перамин. Последние две группы не токсичны для теплокровных, но обладают высокой инсектицидной и инсектдетеррентной активностью. Защита растений от повреждения насекомыми.
")
ГРИБНЫЕ САДЫ МУРАВЬЕВ (ATTINAE)

…И ТЕРМИТОВ

Ophiostoma ulmi – аcсоциация с жукамикороедами

ГАСТЕРОИДНЫЕ АГАРИКОМИЦЕТЫ Melanogaster broomeanus Phallus impudicus Hymenogaster luteus Clathrus ruber

ASCOMYCOTA: подземные аскомы T. melanosporum Tuber magnatum Elaphomyces granulatus T. aestivum

ASCOMYCOTA: Claviceps purpurea «медвяная роса» Анаморфа Sphacelia

Вторичные метаболиты играют важную роль в инфекционном процессе у грибов-паразитов и симбиотрофов

Phylogeny of the kingdom Fungi using analysis of the сombined, six-gene data set. (Timothy et al. , 2006) 1 Школа по систематике грибоввозбудителей болезней 41

Роль в микосимбиотрофии. Антибиотики защищают растения от заражения фитопатогенными грибами. Симуляция развития. Pisolithus arhizus образует антифунгальные метаболиты Биосинтез антифунгальных метаболитов (оксалата и др. ) Paxillus involutus стимулируют эксудаты корней Pinus resinosa

Диффундирующий фактор из грибов арбускулярной микоризы индуцирует экспрессию симбиоз-специфичного nod-зависимого гена Mt. ENOD 11 в корнях Medicago truncata

Hypocreales

Фитотоксины фитопатогенных грибов играют ключевую роль в патогенезе и выживании многих представителей этой группы. Защитная реакция растений – конститутивные токсические метаболиты, механические защитные структуры, образование фитоалексинов ( изофлавоноиды, сесквитерпены, дитерпены, полиацетилены и многие другие). Грибы способны разрушать фитоалексины. .
 образуются A. mali – паразитом яблони;")
Токсины Alternaria – AM –токсины (пептидолактоны – альтернариолид) образуются A. mali – паразитом яблони; A. helianthi, A. chrysanthemi –пиранопироны дезоксирадицинин и радицинин; A. alternata – дикетопиперазин макулозин – избирательно действующий на Centaurea maculosa – опасный сорняк в Канаде и С. З. США.

Sonchus arvensis Аскосонхин, токсин Ascochyta sonchi Cirsium arvense Альтернетаноксины А и В Alternaria sonchi

Cochliobolus victoriae – на Drechslera victoriae ячмене и других злаках Cochliobolus carbonum – на кукурузе НС -токсин

Botrytis cinerea не способен поражать живые клетки растений, необходимо наличие хотя бы небольших участков отмершей ткани. После развития на мёртвом субстрате происходит выделение токсинов, повреждающих соседнюю живую ткань и гриб таким способом распространяется по субстрату.

Токсины Botrytis cinerea

Metarhizium Beauveria bassiana Энтомопатогенные грибы образуют токсины, участвующие в патогенезе Hirsutella

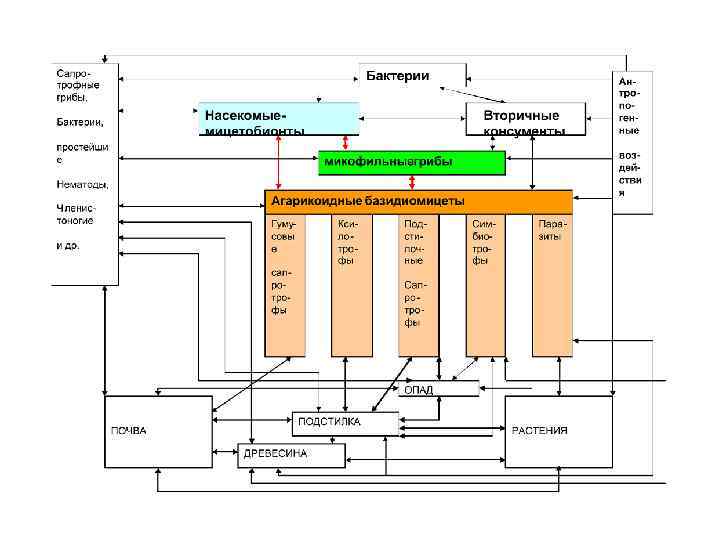
ХИЩНЫЕ ГРИБЫ a, b – Arthrobotrys oligospora c- Monacrosporium haptotalum d- M. gefiropagum e- A. brochopaga f- Drechmeria coniospora Нематоды привлекаются соединениями, выделяемыми мицелием, ловушками хищных грибов и спорами эндопаразитов. Хищные грибы улавливают животных, значительно превосходящих их по размерам. Улавливание таких крупных, подвижных и сильных жертв, как нематоды. стало возможным в результате приобретения грибами в процессе эволюции специализированных аппаратовловушек. Образование нематотоксинов многими хищными грибами
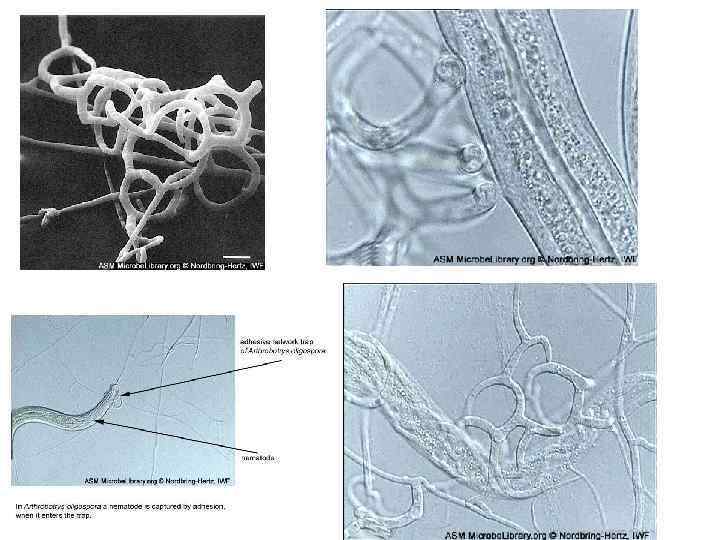

Регуляция образования ловушек происходит разными путями. В отсутствие нематод хищные грибы обычно не образуют ловушек. Если в культуру добавить нематод или воду, в которой находились нематоды, ловушки образуются через несколько часов. В культуре некоторых нематод обнаружен «немин» – вещество или комплекс веществ (низкомолекулярные белки). У А. dactyloides образованиеп ловушек происходит в отсутствие нематод в условиях голодания.
 – ксилотроф, но в условиях дефицита азота в среде этот гриб")
Pleurotus ostreatus (вешенка) – ксилотроф, но в условиях дефицита азота в среде этот гриб способен улавливать нематод и использовать их в пищу. Значительную роль в его «хищничестве» играет нематотоксин

Транспорт металлов Некоторые вторичные метаболиты действуют как агенты транспорта металлов. Важнейшая группа – сидерофоры или сидерамины – поглощение, транспорт и перевод в растворимое состояние железа. Сложные молекулы, переводящие в растворимое состояние ионы железа, растворимость которых составляет 10 -18 моль/л при р. Н 7. 4. Отличаются крайне высоким аффинитетом к железу. Антибиотики-ионофоры – транспорт К (мембраноактивные соединения). Сидерофоры связанные с клеткой – хранение железа в спорах – по аналогии с ферритинами животных и фитоферритинами растений. Neurospora crassa, необходим для прорастания спор. .
")
Защитная роль каротиноидов и меланинов (микробный. лизис, инсоляция, уф, синглетный кислород и др. )

Лектины грибов Известны у макромицетов и микромицетов. Роль в росте, морфогенезе, дифференциации, морфологической реакции на инфекцию и молекулярном узнавании хозяина на ранних стадиях микоризации. Специфичны к (α 2, 3 -)связанным сиалилированным гликанам, у некоторых –к (α 2 -6) связанной сиалевой кислоте, ацетилгалактозоамину, ацетилглюкозамину. . Молекулярная масса 18 -65 k. Da. Гликопротеины с различным содержанием углеводов, состоят из двух или четырех субъединиц (димеры или тетрамеры, гетеро- или гомо) ), термостабильны, устойчивы к экстремальным значениям р. Н. У некоторых грибов не гликозилированы. Volvariella volvacea. У разных групп роль разнообразная. Dictyostelium discoideum - дискоидин 1 – межклеточная адгезия в процессе морфогенеза. Хищные грибы – поверхностный лектин Arthrobotrys oligospora -улавливание нематод. Дрожжи Kluyveromyces bulgaricus – лактоза –специфичные лектины. Митогенное, иммуномодулирующее, противоопухолевое, сосудорасширяющее, гипотензивное действие. Эксперименты и практический интерес.

РЕГУЛЯЦИЯ РОСТА КОЛОНИЙ И КОНИДИОГЕНЕЗА

Биосинтез ВМ и спорообразование регулируется общими факторами. Регуляция прорастания, защита спор от поедания (амебы и др. ), конкуренция при прорастании. Глобальный регулятор ВМ транскрипции Lae. A- ядерный белок. Контролирует экспрессию многих ВМ, включая антибиотики и микотоксины. Делеция гена Lae. A приводит к активному росту при снижении или элиминации транскриптов генов ВМ.

Автоингибиторы прорастания спор. Cостояние покоя. Регуляция плотности популяции. Дискаденин – у миксомицетов Dictyostelium (образуется из 5 -АМР). Прорастание урединиоспор Puccinia coronata –метил-3, 4 -иметоксициннамат. Автоингибитор Colletotrichum graminicola Циклический АМР – у диктиостелиум , аттрактант для амеб и их аггрегации. Связь со стадиями развития. Penicillium patulum – образование 6 -метилсалициловой кислоты только после образования воздушного мицелия. У P. brevicompactum – микофеноловая кислота, асперфенамат и эргостерин образуются перед формированием конидий, а бревианамды – в процессе образования конидий.

АВТОРЕГУЛЯЦИЯ ПРОРАСТАНИЯ СПОР: ингибиторы прорастания
 Сиренин (сесквитерпен) – феромон + гаметангиев и гамет у Allomyces spp.")
Половые гормоны (феромоны) Сиренин (сесквитерпен) – феромон + гаметангиев и гамет у Allomyces spp. Хемотаксис – подвижные гаметы, очень высокая активность (0. 1 нг/мл).

Триспоровые кислоты у мукоровых Индукция образования зигофоров. На начальной стадии синтеза у + и – мицелиев. Триспоровые к-ты- только при контакте.
 – регулятор полового")
У аскомицетов – пептидные факторы агглютинации у Saccharomyces cerevisiae Зеараленон (токсин) – регулятор полового размножения у Gibberella zeae; пси-фактор у Aspergillus nidulans.
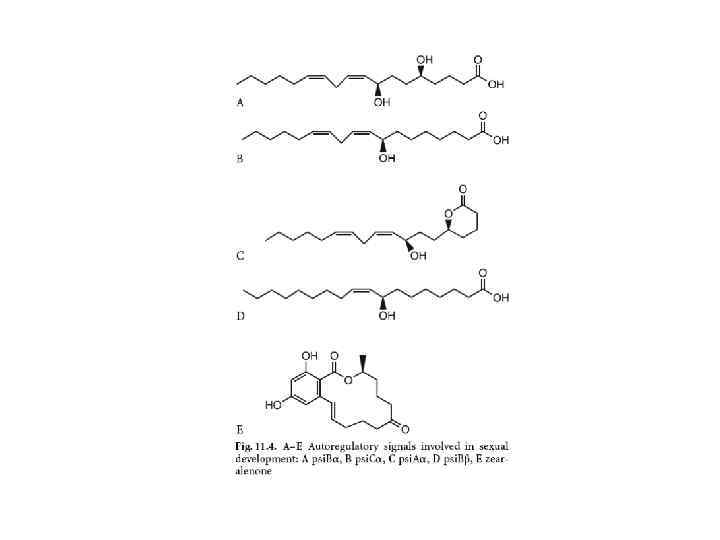

У базидиомицетов – пептид родоторуцин А у клеток А-типа у Rhodosporidium toruloides, вызывает образование трубок скрещивания у типа а. Tremella mesenterica (тремелломицеты) пептид тремероген А-10, индуцирующий ростковые трубки у типа а.

Антеридиол у грибоподобного оомицета Achlya – стероидный вторичный метаболит - образуется + мицелием и инициирует образование – гаметангиев (10 – 11 моль/л ). Последние также образуют вторичный метаболит (гормон В), стимулирующий образование на +мицелии оогониев


Микоспорины и микоспоринподобные аминокислоты Низкомолекулярные (менее 400 Da, у некоторых до 1800 Da, например, Nostoc) водорастворимые соединения с характерной абсорбцией 310 -363 нм. Состоят из кольца аминоциклогексенона или аминоциклогексимина, содержат азот- или иминозамещенные спирты. Известны гликозилированные производные микоспоринов. У грибов – микоспорин-глутамикол-глюкозид и микоспоринглутаминол- глюкозиды. Образуются широким кругом организмов, в том числе и грибами. УФ –абсорбирующие пигменты, открытые относительно недавно (в 60 -х годах прошлого века), у грибов –немного позже – 1976 -1979 (Favre-Bonvin et al. , Arpin et al. ). Сразу же была обнаружена их роль в защите клетки от УФ радиации, сейчас показана их многофункциональность: антиоксиданты, защита от кислородных радикалов, солевого стресса, защита от высушивания и температурного (термического) стресса, внутриклеточный резервуар азота, роль в размножении грибов- споруляция, индуцированная светом. Могут индуцировать спорогенез в отсутстие света. Микроколонии грибов, растущие на камне.
Лекция 12. Вторичные метаболиты.ppt