02 Возбудимые ткани.ppt
- Количество слайдов: 57



Возбудимые ткани Доцент Зверев М. Д. 2011 г.
. Мебранно-ионная теория возникновения МП 2. Локальный ответ (ЛО).")
Цели лекции: 1. Мембранный потенциал (МП). Мебранно-ионная теория возникновения МП 2. Локальный ответ (ЛО). Фазы ЛО. 3. Свойства ЛО 4. Потенциал действия (ПД). Фазы ПД. механизм генерации потенциала действия 5. Свойства ПД 6. Методы регистрации МП и ПД
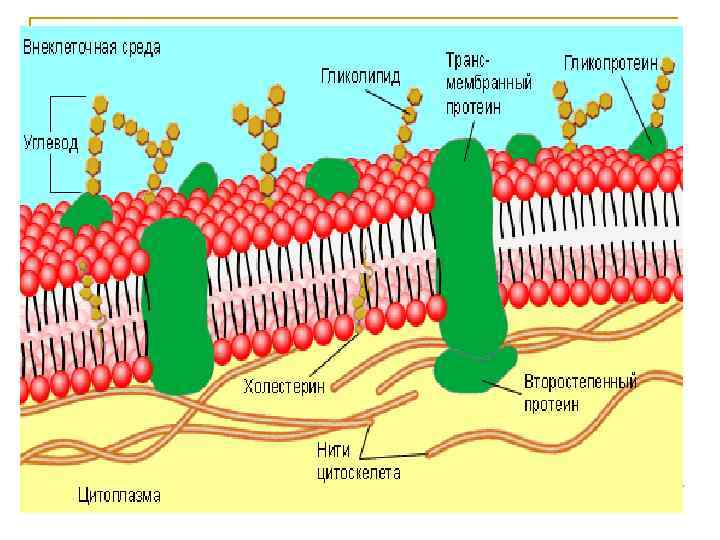
 Толщина около 7,")
Плазмалемма и другие мембраны клетки Жидко-мозаичная модель (1971 г. , Зингер-Николсон) Толщина около 7, 5 нм. Основа – билипидный слой и белки. На плазмалемме углеводный компонент образует гликокаликс (по массе 2 -10%, связан либо с липидами – гликолипиды, либо с белками - гликопротеиды). Снаружи внутренних мембран гликокаликса нет. В составе билипидного слоя преобладают фосфолипиды; внутренние мембраны мягче, т. к. в них больше ненасыщенных ж. к. и меньше холестерина. Незаякоренные молекулы фосфолипидов быстро перемещаются (до 2 мкмс).

ПОЛОЖЕНИЕ ХОЛЕСТЕРОЛА В ЛИПИДНОМ БИСЛОЕ Фосфолипиды 3 «Полярные головки» нм 2 1 0 Область холестерола, придающего жесткость мембране Область гидрофобного хвоста, обладающего большей жидкостностью

Мембранные белки плазмалеммы обычно составляют по массе около 50%, но во внутренних мембранах их количество может существенно отличаться – в мембранах митохондрий белки составляют 75% по массе, т. к. многие из них выполняют каталитические и транспортные функции, обеспечивающие дыхание. По расположению в мембране белки распределяются на несколько групп. Мембранные белки Периферические Погруженные Интегральные 1 Пронизывающие (трансмембранные) 2 1 - вязкий, рыхлый гликокаликс 2 - собственно плазмалемма 3 - кортикальный слой 3

Ионные каналы • Канальные белки формируют трансмембранные поры, через которые могут пассивно проходить маленькие водорастворимые молекулы, как в клетку, так и из клетки 1 • Большинство канальных белков плазматической 3 4 мембраны формируют поры, проницаемые только для 5 неорганических ионов – ионные каналы 6 2 7 • Ионные каналы высокоселективны для специфических 8 +, K+, Ca 2+, Cl-) ионов ( Na 9 1 • Ионную специфичность канала определяют: 0 диаметр – через 1 узкую пору не могут пройти большие ионы 1 форма – только 1 одиночные ионы могут беспрепятственно 2 проходить через 1 канал 3 Заряд – отдельные участки канала имеют разный заряд
 Ионная селективность – через канал")
Виды ионных каналов Два принципиальных свойства ионных каналов: (1) Ионная селективность – через канал может проходить только один вид ионов (2) Состояния: закрыты, открыты, инактивированы потенциалмеханохемочувствительные внеклеточные внутриклеточные +++ out --- in закрыты открыты + + out - - in Мембранный потенциал Молекула связывается с каналом Растяжение

Животное электричество
")
Животное электричество Луиджи Гальвани (1737 -1798)
 мембране. Движение ионов через мембрану возможно")
Все электрические процессы клетки разворачиваются на плазматической (поверхностной) мембране. Движение ионов через мембрану возможно при наличии разности их концентраций снаружи и внутри клетки (концентрационный градиент) Электрические токи, возникающие в клетке, обеспечиваются движением ионов через мембрану
 МП – это разность зарядов между внутренней и наружной поверхностью мембраны.")
МЕМБРАННЫЙ ПОТЕНЦИАЛ (МП) МП – это разность зарядов между внутренней и наружной поверхностью мембраны. Внутренняя поверхность заряжена (-) а наружная (+) Мембранный потенциал покоя (МП) можно определить как Vm = Vin – Vout. , где Vin - потенциал внутри клетки, Vout - снаружи. Поскольку потенциал снаружи клетки можно принять за ноль, то МПП равен Vin. n
 Экспериментально доказали")
Причины поляризации клеточной мембраны Мембранно-ионная теория Теоретически разработал -Юлиус Бернштейн (1839 -1917) Экспериментально доказали – Ходжкин, Катц и. Хаксли n n n 1. Разность концентраций для ионов Na+ , K+ , Са +, A-. 2. Избирательная проницаемость мембраны в покое для ионов. Свободно проницаема для ионов калия и практически непроницаема для других ионов. Калий выходит из клетки- внутри становится (-), а с наружи (+). 3. Работа электрогенного Na+-K+ насоса

МЕМБРАННЫЙ ПОТЕНЦИАЛ Причины возникновения МП: 1. Разность концентраций ионов внутри и вне клетки. Высокий градиент концентрации K+, Na+, Сl, А- орган. анионов 12 раз ↑ 40 раз↑

Для того, чтобы ионы могли двигаться через мембрану необходимо иметь мембранные структуры, сообщающие вне- и внетриклеточную среду (ионные каналы) 1 -1000 каналов на квадратный микрометр мембраны

2. Избирательная проницаемость мембраны в покое для ионов K+ n n n Неуправляемые K+ (не имеющие ворот) ионные каналы открыты в состоянии покоя и создают возможность диффузии ионов K+ через мембрануионы K+ выходят наружу. Внутренняя поверхность мембраны становится (-) заряженной, а наружная (+) заряженной – возникает МП. Из клетки выходит небольшое количество ионов K+. Электростатические силы - (-) заряд внутр. мембраны удерживают K+ внутри клетки. В покое существует равновесие между концентрационным и электрохимическим градиентами K+ В покое мембранный потенциал и поток ионов через мембрану сбалансированы

В покое: n Калиевые каналы n 1. открыты в большом количестве 2. создают на внутренней поверхности мембраны отрицательный заряд n Натриевые каналы n n n 1. В основном закрыты 2. Ионы натрия перемещаются медленно

Генерация мембранного потенциала покоя - пассивный процесс, который не требует затраты энергии. Однако энергия необходима для поддержания первоначального концентрационного градиента, а также для его поддержания в процессе активности клетки.
 Разность концентраций ионов K+ и Na+ поддерживается работой")
3. Работа электрогенного Na+-K+ насоса 1) Разность концентраций ионов K+ и Na+ поддерживается работой Na+-K+ АТФ-азы 2) Увеличивает МПП на 11 м. В

Величина МП n. Ск. мышцы. : - 90 м. В; nнервн. кл-ки: - 60 -70 м. В. ; nгл. мышцы: - 40 -50 м. В

Изменения мембранного потенциала покоя n n n Деполяризация- уменьшение Реполяризация- возвращение к исходному уровню Гиперполяризация- увеличение 0 1 Деполяризация -30 -60 -90 Реполяризация 2 Гиперполяризация МПП Время

ПОТЕНЦИАЛ ПОКОЯ

Регистрация мембранного потенциала покоя Б А 1. Внутриклеточная микроэлектродная регистрация Величина МП в возбудимых клетках – от -60 до -90 м. В Введение электрода 0 -30 -60 А Б Мембранный потенциал покоя Время

Регистрация мембранного потенциала покоя 2. Использование флуоресцентных красителей
, необходимая для возникновения потенциала действия.")
Порог раздражения- n n минимальная сила раздражителя (электрического тока), необходимая для возникновения потенциала действия. Подпороговый раздражитель - меньше по силе, чем пороговый Сверхпороговый раздражитель – больше по силе порогового
 раздражители вызывают местные изменения заряда")
Действие раздражителя на ткань Возбуждение n n Слабые (подпороговые) раздражители вызывают местные изменения заряда клеточной мембраны – локальный ответ Сильные (пороговые и надпороговые) раздражители вызывают появление инверсии заряда мембраны – потенциала действия
 n ЛО – это местный процесс возбуждения возникающий при действии раздражителя")
Локальный ответ (ЛО) n ЛО – это местный процесс возбуждения возникающий при действии раздражителя подпороговой величины

Локальный ответ Свойства локального ответа n n n Подчиняется закону силы Способен к суммации Распространяется на малые расстояния с декрементом КУД

+30 м. В ПД – универсальный ответ нервной клетки на стимуляцию +30 и более м. В (вершина, овершут: область положительных значений) 0 измерение и стимуляция -50 Подаем через микроэлектрод короткие электрич. импульсы нарастающей амплитуды КУД 1 -2 мс -70 5 м. В 10 м. В время, мс 20 м. В: пороговый стимул при ПП= -70 м. В 29

Локальные ответы и закон силы

+30 м. В Суммация ЛО 0 КУД -55 -70 5 м. В время, мс 15 м. В: пороговый стимул при ПП= -70 м. В 31


Нобелевская премия 1963 года в области физиологии и медицины Алан Ходжкин Эндрю Хаксли Сэр Джон Экклс за открытия ионных механизмов возбуждения и торможения нервных клеток
 ПД – это быстрое колебание мембранного потенциала с изменением заряда. Во")
Потенциал действия (ПД) ПД – это быстрое колебание мембранного потенциала с изменением заряда. Во время ПД заряд мембраны внутри клетки становится (+), а снаружи (-). ПД формируется когда мембрана частично деполяризуется до критического уровня. (NB!) Критический уровень деполяризации для мембраны нейрона — -55 м. В. Fig. 48. 8 c Campbell and Reece, 6 th Ed.

Потенциал действия нервного волокна
. Фазы ПД (ПД 3 2 - 55 м. В КУД 1")
Потенциал действия (ПД). Фазы ПД (ПД 3 2 - 55 м. В КУД 1 0 4 0 5 МП

Потенциал действия - Фазы ПД n Действие раздражителя → 1 - медленная деполяризация (локальный ответ) - активация потенциал зависимых Na каналов → вход Na+ в клетку → деполяризация до мембраны критического уровня деполяризации (КУД) → 2 - быстрая деполяризация - лавинообразный вход Na+ в клетку → инверсия заряда мембраны [внутри (+), снаружи (-) ] → инактивация Na каналов (закрытие) → 3 - реполяризация - усиление выхода К+ из клетки → следовые потенциалы 4 - следовая деполяризация, 5 - следовая гиперполяризация


Потенциал действия зависит от внеклеточного Na

Воротный механизм электровозбудимых Na+-каналов Динамика открытия ворот 1 - покой 2 -деполяризация 3 -Спайк (рефрактерность) За один ПД входит в клетку 1012 ионов Na+ (рост внутриклеточной концентрации 0, 7%) 1 2 3

Для закрытия Na+-каналов на пике ПД служит дополнительная внутриклеточная, инактивационная створка – h-ворота. n Вторая створка - активационная – m-ворота. Если заряд внутри нейрона вновь ниже -50 м. В – створка h- ворот закрывается, т. к. положительные заряды, расположенные на ней, притягиваются к отрицательно заряженным ионам цитоплазмы, m- ворот – закрывается. n

Блокирование потенциал-управляемых натриевых каналов нарушает генерацию потенциала действия Na+ Тетродотоксин – специфический блокатор натриевых каналов

В результате действия токсина прекращается генерация и проведение ПД: сначала – по периферическим нервам ( «иллюзии» кожной чувствительности, параличи, нарушения зрения и слуха), позже – потеря сознания; смерть от остановки дыхания (сэр Джеймс Кук). электрочувствительный Na+-канал тетродотоксин – яд рыбы фугу (аминогруппа работает как «пробка» для Na+-канала) 43

Блокада калиевых каналов тетраэтиламмонием резко удлиняет процесс реполяризации

Блокирование потенциал-управляемых калиевых каналов резко затягивает потенциал действия K+ K+ Тетраэтиламмоний – специфический блокатор калиевых каналов

ТЭА – тетраэтиламмоний: работает как «пробка» по отношению к К+-каналу. В результате восходящая фаза ПД изменяется мало, нисходящая – затягивается до 50 и > мс (реполяризация происходит за счет постоянно открытых К+-каналов, которых примерно в 100 раз <, чем электрочувствительных); ТЭА вызывает глубокую потерю сознания. «затянутый» ПД на фоне ТЭА: 50 мс ПД в норме: 1 мс 46

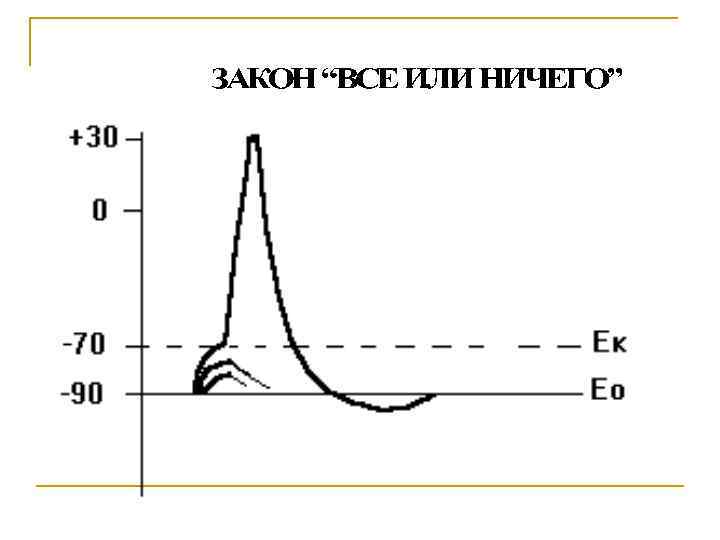
Свойства потенциала действия Вызывается сверхпороговым раздражением Подчиняется закону «Все или ничего» Распространяется по всей мембране не затухая Не суммируется
")
Обмен веществ и энергии при возбуждении. Теплообразование при возбуждении метаболический хвост кометы возбуждения (Ухтомский) 2 -10% начальное тепло 90% запаздывающее тепло

Рефрактерность снижение способности клетки отвечать на раздражение в результате временной инактивации натриевых каналов Абсолютная рефрактерность Генерация ПД невозможна Вызвана инактивацией большинства Na каналов Относительная рефрактерность Генерация ПД возможна при увеличении интенсивности раздражителя 0 Связана с тем, что некоторая часть Na каналов все еще инактивированы
: а. Супернормальной (повышенной) возбудимости; n б. Абсолютной рефрактерности;")
Изменения возбудимости во время возбуждения (ПД): а. Супернормальной (повышенной) возбудимости; n б. Абсолютной рефрактерности; n в. Относительной рефрактерности; n г. Супернормальной (повышенной) возбудимости n д. Субнормальной (пониженной) возбудимости.

Абсолютная рефрактерность n Генерация ПД невозможна n Вызвана инактивацией большинства Na каналов Относительная рефрактерность n Генерация ПД возможна при увеличении интенсивности раздражителя n Связана с тем, что некоторая часть Na каналов все еще инактивированы

Виды регистрации ПД Внутриклеточная монополярная Внеклеточная биполярная

Внеклеточная биполярная регистрация ПД Электромиография Произвольное сокращение : Слабое Среднее максимальное

Регистрация ЭКГ в начале 20 века

Аппарат, читающий мысли. Эти датчики на шее снимают слабые биотоки, которые компьютер переводит в слова

Успехов Вам в достижении заоблачных высот при
02 Возбудимые ткани.ppt