ВОПРОСЫ К ГЭКУ ПО ФЗР 2013.ppt
- Количество слайдов: 50



ВОПРОСЫ К ГЭКУ ПО ФЗР 1. Митохондрии: общая морфология, ультраструктура, функции. Основные этапы дыхания: гликолиз, цикл трикарбоновых кислот, электронтранспортная цепь, синтез АТФ. 2. Хлоропласты: общая морфология, ультраструктура, функции. Характеристика основных этапов фотосинтеза. Значение фотосинтеза.
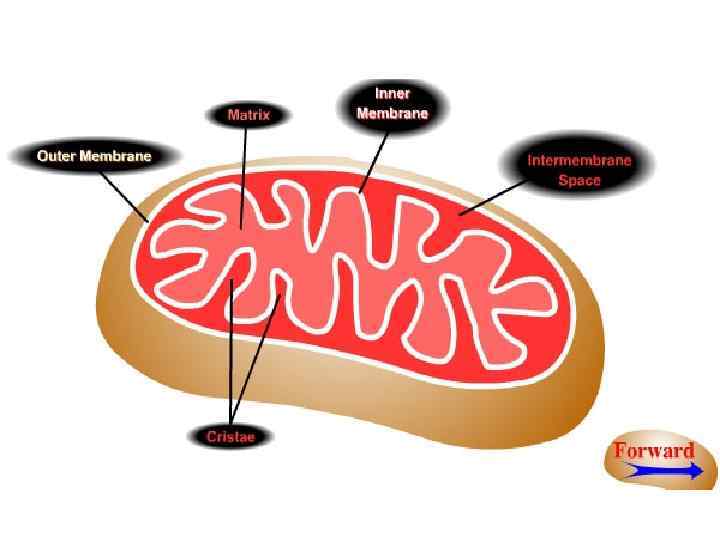

Наружная мембрана — главная функция ее - отделение внутренней части митохондрии от цитоплазмы. Мембрана проницаема для неорганических ионов и относительно крупных молекул, в том числе аминокислот, некоторых белков, АТФ, сахарозы, промежуточных продуктов дыхания. • В перимитохондриальном пространстве содержится большая часть митохондриальной воды (остальная в матриксе), ферменты (аденилаткиназа, АДФ-АТФ-ферменты фосфорилирования, не зависящие от дыхательной цепи). • Внутренняя мембрана по сравнению с наружной непроницаема для всех веществ в ионизированном состоянии, незначительное количество метаболитов пересекает ее в недиссоциированной форме, большая часть - при помощи разных систем переноса.
 (2 АТР 2 NADPH ) ГЛИКОЛИЗ ПФЦ (NADPH — 17 АТФ")
ГЛЮ (С 6) (2 АТР 2 NADPH ) ГЛИКОЛИЗ ПФЦ (NADPH — 17 АТФ из ЭТЦ) 2 ПВК (С 3) 2 NADPH АЦЕТИЛ Ко. А 2 АТР 2 СО 2 ЦТК 2 СО 2 8 NADPH АТР ФМН 2 FАD Ко. Q ЦИТ в, с1 АТР ЦИТ с ЦИТ а, а 3 2ê +2 Н+ АТР ½ О 2+2ê+ 2 Н+ Н 2 О
. •")
Гликолиз – последовательность биохимических реакций, приводящая к превращению гексозы в ПВК (пировиноградную кислоту). • Происходит в цитоплазме, ферменты гликолиза не связаны с мембранными структурами.
 С 6 С 3 Субстратное фосфорилирование")
Гликолиз (путь Эмбдена-Мейергофа-Парнаса) С 6 С 3 Субстратное фосфорилирование

Субстратное фосфорилирование +Н 3 РО 4 Субстратное фосфорилирование

Значение гликолиза • С 6 Н 12 О 6 + 2 АТФ + 2 НАД+ + 4 АДФ + 2 ФН 2 С 3 Н 4 О 3 + 4 АТФ + 2 НАД. Н • Запасается 2 АТФ и 2 НАДН • Гликолиз важен для клетки прежде всего как поставщик не энергоносителя, а высокореакционного соединения — ПВК, способного включаться в процесс кислородного окисления
 • в матриксе - ЦТК, • в мембранах - ЭТЦ")
Аэробная фаза (ЦТК, ЭТЦ) • в матриксе - ЦТК, • в мембранах - ЭТЦ
 • • Идет в 3 стадии: а) активирование ПВК; б) окисление")
Аэробная фаза (митохондрии) • • Идет в 3 стадии: а) активирование ПВК; б) окисление ПВК в ЦТК (цикл Кребса); в)перенос Н+ и ē в ЭТЦ, при этом О 2 воздуха подключается к данному процессу только на последней стадии.
 Активирование ПВК • Активирование ПВК проходит с участием сложного полиферментного комплекса (3 фермента")
а) Активирование ПВК • Активирование ПВК проходит с участием сложного полиферментного комплекса (3 фермента + 5 коферментов - Ко. А, липоевая кислота, тиаминпирофосфат, ФАД, НАД). • Следует отметить, что ацетил - Ко. А – • 1)один из самых важных промежуточных продуктов метаболизма, узловой пункт перекрещивания многих путей, • 2)связующее звено между обменом жиров и углеводов, • 3)точка, с которой начинаются многие процессы биосинтеза.

Цикл трикарбоновых кислот 1939 г. А. Чибнелл

1 СТАДИЯ: ПВК + NAD + SH ~ K 0 A CH 3 CO ~ S · K 0 A + NADН + СО 2 (I) 2 СТАДИЯ: CH 3 CO ~ S – K 0 A + 3 H 2 O + 2 NAD+ + NADР+FAD + ADP + PJ 2 CO 2 + 2 NADH + NADРН +FADH 2 + ATP + HS – K 0 A ЭТЦ СУЩНОСТЬ ЦТК: 1. ДЕКАРБОКСИЛИРОВАНИЕ (ВЫДЕЛЕНИЕ 2 СО 2) 2. ПОГЛОЩЕНИЕ Н 2 О 3. ВЫДЕЛЕНИЕ Н+ Н+ акцептируется в форме NADPH+ и (2 Н+ + 2ē) ЭТЦ FADH 2 (2 Н+ + 2ē)
 • Приблизительно 10% площади внутренней мембраны заняты ЭТЦ, разделенной на 4 блока.")
ЭТЦ (дыхания) • Приблизительно 10% площади внутренней мембраны заняты ЭТЦ, разделенной на 4 блока. • Компоненты цепи занимают разное положение по толщине мембраны. У растений существуют 2 НАДНзависимые дегидрогеназы: одна локализована с внутренней стороны мембраны и катализирует окисление образованного в матриксе НАДН, • другая находится с наружной стороны мембраны и катализирует окисление НАДН, генерированного вне митохондрии. СДГ расположена с внутренней стороны кристы. Флавопротеины (НАДН-ДГ, СДГ) передают электроны через расположенные в центре цитохромы “в” к цитохромам “с”, которые находятся с наружной стороны кристы. • Компоненты цитохромоксидазы расположены поперек внутренней мембраны

Расположение блоков компонентов ЭТЦ и АТФ-синтетазы в кристах. Расположение блоков компонентов ЭТЦ и АТФсинтетазы в кристах. Цитохром соксидазный Убихинон-цитохром с оксидоредуктазой СДГ НАД(Р)Ндегидрогеназный
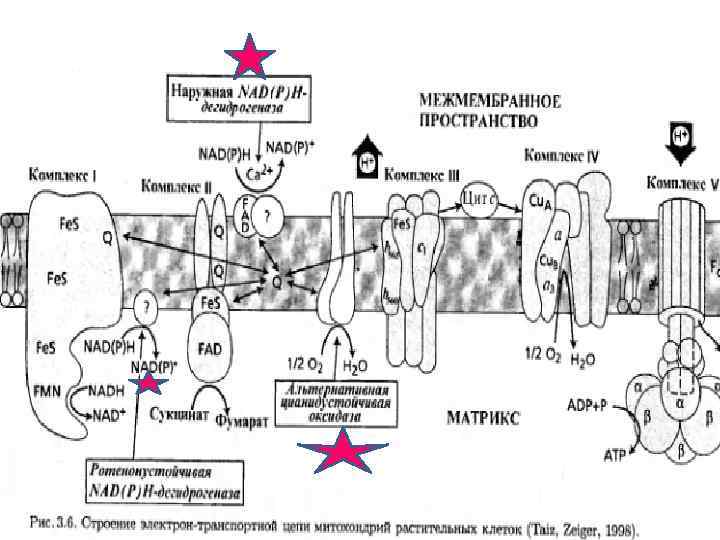

Перенос Н+ и е- в ЭТЦ • Процесс проходит на мембранах митохондрий, где НАД. Н и ФАД. Н 2 через ряд этапов окисляются в окислительно-восстановительной цепи в и на внутренней мембране митохондрий. • Переносчики Н+ и е- - ферментные системы, расположенные по градиенту окислительно-восстановительных потенциалов в порядке их возрастания. • В живой растительной клетке роль системы с максимальной величиной положительного потенциала выполняет О 2(+0, 817 В). • Роль доноров электронов принадлежит молекулам различных органических веществ (ЦТК). Но отдаваемые ими электроны достигают О 2 не непосредственно, а через цепь о. в. систем, потенциалы которых расположены между потенциалами органических веществ и О 2. • Звенья цепи - ферментные системы.

В состав ЭТЦ дыхания входят: • 1. флавопротеины, содержащие в качестве простетической группы - ФАД или ФМН; • 2. цитохромы - с простетической группой гемом; • Fe-S-белки с простетической группой из негемового железа, комплексно связанного с неорганической серой или серой цистеина; • убихинон - липид, состоящий из бензохинона, соединенного с длинной гидрофобной изопреноидной боковой цепью.
 флавопротеины и убихинон переносят Н+ и ē, а цитохромы")
Следует отметить, что: • 1) флавопротеины и убихинон переносят Н+ и ē, а цитохромы и Fe-S-белки - только ē; • 2) ФП и убихинон переносят 2 ē, цитохромы и Fe -S-белки – 1 ē. • Поскольку НАДН 2 и сукцинат, которые окисляются в ЭТЦ, являются двухэлектронными окислительно-восстановительными системами, • то цитохромам и Fe-S -белкам необходимо работать парами.
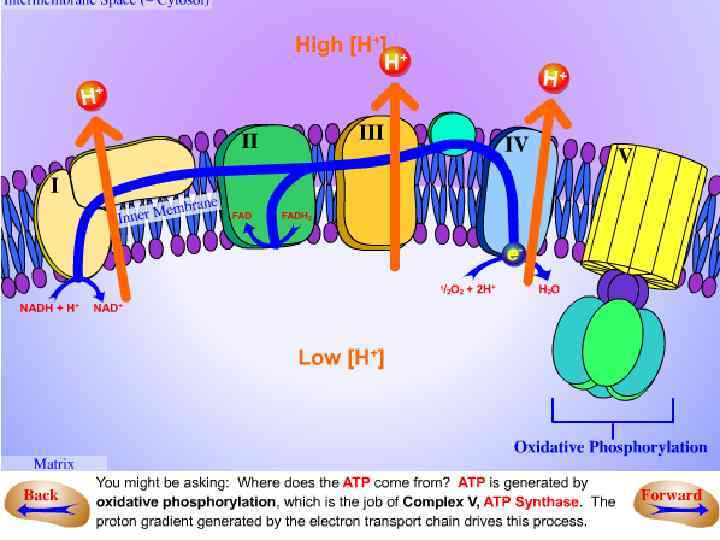

• Таким образом, • в ЭТЦ идет многоступенчатая передача электронов от восстановленного субстрата до О 2, сопровождающаяся трансмембранным переносом протонов Н+, • т. е. ЭТЦ митохондрий (как и хлоропластов ) выполняет функцию окислительновосстановительного Н+-насоса.

Генерация АТФ • Свободная энергия, высвобождающаяся при переносе пары электронов по ЭТЦ, используется для фосфорилирования АДФ и, следовательно, образования АТФ. Это впервые было показано в 1940 г. Белицер, а ранее Энгельгардтом (1929). • Превращение энергии, выделившейся в ЭТЦ, в энергию химических связей АТФ происходит в процессе фосфорилирования. окислительного • Различают два типа фосфорилирования - субстратное и фосфорилирование в дыхательной цепи (окислительное фосфорилирование). • Однако, основная часть энергии при окислительном распаде питательных веществ в ЦТК выделяется и аккумулируется в ЭТЦ.

Окислительное фосфорилирование — • процесс фосфорилирования АДФ с образованием АТФ, сопряженный с переносом электронов по ЭТЦ митохондрий.
 впервые установил количество молекул АТФ, образующихся при переносе")
В 1941 г. Очоа (Ochoa ) впервые установил количество молекул АТФ, образующихся при переносе пары электронов по ЭТЦ, определив коэффициент Р/O - количество молей ортофосфата, используемого для фосфорилирования АДФ, в расчете на 1 г/атом (половину моля )поглощенного О 2, который восстанавливается до Н 2 О. (Для НАДН Р/О=3, сукц. =2 ). • Определение Р/О легло в основу концепции пунктов фосфорилирования. Если при переносе е- по ЭТЦ образуется 3 АТФ, то должно быть 3 пункта, где они могут синтезироваться. • Согласно современным представлениям эти пункты находятся между: • • Между НАДФН и железосерным белком Fe. SN 2 (I комплекс), Убихиноном и цит. “с1”, в цитохромоксидазе. Именно здесь выделяется достаточное количество энергии (более 30 к. Дж/моль) для образования макроэргической связи АТФ.
;")
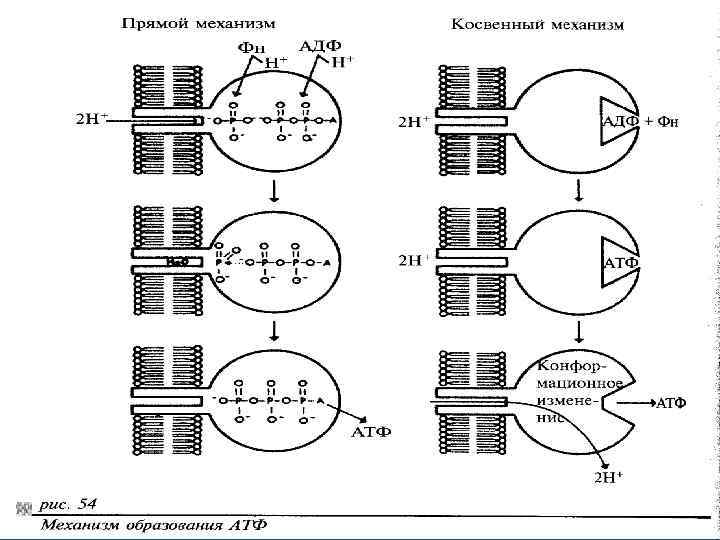
Основные гипотезы механизма окислительного фосфорилирования • 1. Гипотеза химического сопряжения (Слейтер и др. ); • 2. Гипотеза механохимического сопряжения (Бойер, Кинг); • 3. Хемиосмотическая теория (Митчелл, Скулачев)
 • Движение è по ЭТЦ сопровождается транспортом Н+ из матрикса")
Хемиосмотическая гипотеза (Mitchell, Скулачев) • Движение è по ЭТЦ сопровождается транспортом Н+ из матрикса в межмембранное пространство митохондрий. В результате на внутренней мембране митохондрий формируется Н+, который является непосредственным источником энергии для синтеза АТФ. • Для этого необходимы следующие условия: • 1) чередование переносчиков, которые переносят протоны и электроны и только электроны; • 2) наличие 3 петель для переноса 6 Н+; • 3) непроницаемость мембраны для Н+ (целостность, сопряженная мембрана); • 4) асимметрия организации ЭТЦ (в мембране). • Синтез АТФ осуществляется в процессе транспорта ионов Н+ по градиенту концентрации ч-з протонный канал фермента АТФ-синтетазы.

• Хлоропласты: общая морфология, ультраструктура, функции. Характеристика основных этапов фотосинтеза. Значение фотосинтеза.

Фотосинтез процесс поглощения солнечного света фотосинтетическими пигментами, превращение данной энергии в энергию химических связей (АТФ и НАДФ·Н) и использование ее для преобразования углерода из неорганической формы (СО 2) в органическую, при этом выделяется кислород.

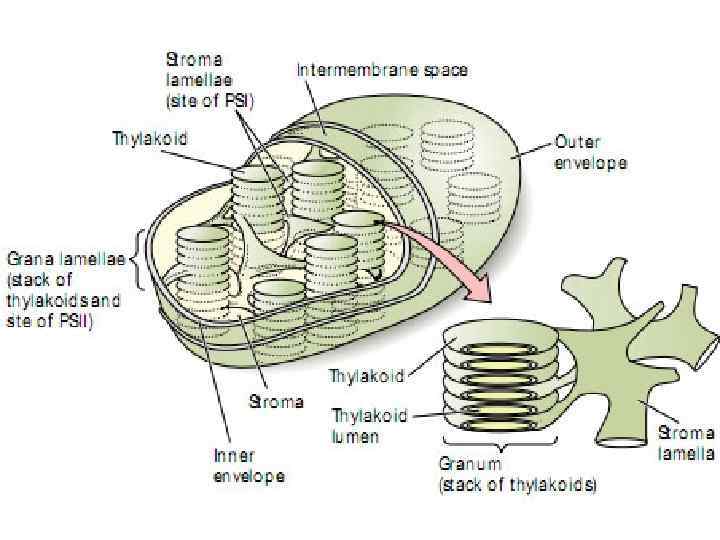
Современные представления о строении хлоропласта 1. 2. 3. В ранней концепции внутренней структурной организации хлоропластов высших растений предполагалось, что существует два типа тилакоидов, различающихся по размерам: большой тилакоид (стромы) и меньший (граны). Менке (1962) высказал предположение, что мембраны располагаются параллельно другу, имеют двухмерную структуру. Вейер предложил позднее гранулярно-решетчатую структуру (сетчатую). Размер перфораций существенно варьировал, часто они были такими большими, что мембраны, которые соединяли между собой граны, представляли не более чем узкие трубочки. Вермейер и Паолино назвали их фретами (1970) (рис. 2 Б). Хеслон - Харрисон высказали мысль о том, что граны соединяются между собой не только в горизонтальном, но и в вертикальном направлении. Каждый диск граны имеет множественные сообщения с другим диском через фреты (рис. 2 В - Д).

Значение фотосинтеза • 1. Синтез органических соединений, необходимых для восполнения убыли органических веществ. • 2. Запасание химической энергии в продуктах фотосинтеза. • 3. Поддержание определенного содержания О 2, образование озонового экрана. • 4. Поглощение СО 2 и предотвращение накопления СО 2 в атмосфере. Борьба с парниковым эффектом.

Фотосинтез — • Это сложный многоступенчатый окислительновосстановительный процесс, в котором при поглощении света и при участии хлорофилла происходит окисление воды до кислорода и восстановление углекислого газа до уровня углеводов. h • 6 CO 2 + 12 H 2 O _____ С 6 Н 12 О 6 + 6 O 2↑ + 6 H 2 O • Хлорофилл

«Ассими ляционная сила» Сущность фотосинтеза

Световые и темновые реакции Световые реакции: Темновые реакции: Зависят от света Не зависят от температуры Зависят от температуры Быстрые < 10 -5 (сек) Медленные ~ 10 -2 (сек) Протекают на мембранах тилакоидов гран и стромы Протекают в строме

Процесс фотосинтеза: Световая v. I - фотофизический стадия v. II- фотохимический v. III- темновой (ферментативный)

Сущность световой стадии • Поглощение квантов света молекулой хлорофилла РЦ с участием дополнительных пигментов и трансформация Е квантов h в Е • АТФ и восстановленного надфосфата (NADP·H)

Световые реакции 1. Введение энергии в биологические системы через воспринимающие пигментные системы 2. Преобразование энергии света в «биологическую энергию»

Пути дезактивации электронно-возбужденных состояний
 =РЦ + ФЕ (ССК) + ЭТЦФС • Пигменты, которые улавливают")
• Фотосистема (ФС) =РЦ + ФЕ (ССК) + ЭТЦФС • Пигменты, которые улавливают свет, объединяются в фотосинтетическую единицу — ФЕ. • В растениях существуют 2 системы ФС I и ФС II. • Компоненты ФС I встроены в тилакоидные мембраны стромы, • а компоненты ФС II встроены в тилакоиды гран
 собирают")
• 200 -400 молекул хлорофилла и других пигментов (каротиноиды, фикобилины в водорослях) собирают h - это ССК (светособирающий комплекс, который собирает свет) и передают их в РЦ. • Сборщики более коротковолновые, чем хлорофилл из РЦ.

Функции РЦ: • Получение Е от вспомогательных пигментов и поглощение Е; • Трансформация Е квантов h Е возбужденного состояния и в химическую Е первичных фотовосстановленных и фотоокисленных соединений
")
Механизмы энергетического переноса на фотофизическом этапе Индуктивный резонанс Полупроводниковый механизм (гипотеза о миграции экситона)

Темновая фаза фотосинтеза — это совокупность биохимических реакций, в результате которых происходит усвоение растениями углекислого газа атмосферы (СО 2) и образование углеводов. Ферменты, катализирующие темновые реакции растворены в строме.
 умеренный климат С 4 -путь, кооперативный фотосинтез тропики САМ")
С 3 -путь (цикл Кальвина) умеренный климат С 4 -путь, кооперативный фотосинтез тропики САМ или МОКТ засушливые зоны

Отличие темновой стадии у С 3 от С 4 и САМ-растений — С 3 + доп. реакции превращения СО 2: Ø временем суток, когда эти реакции происходят, Ø в каких клетках находятся вещества, участвующие в этих реакциях.

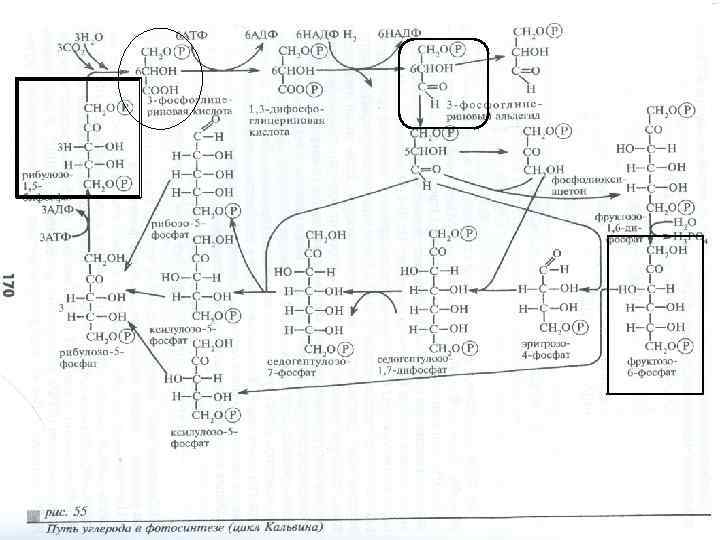
Сущность темновых реакций процесса фотосинтеза была раскрыта: Мельвином Кальвином (Нобелевская премия в 1961 г. ) с сотрудниками, Бэнсоном, Басхэмом.
, Ø восстановление (СО 2")
Цикл можно разделить на 3 фазы: Ø карбоксилирование (+СО 2), Ø восстановление (СО 2 до органических веществ), Ø регенерация (акцепторов).

Предварительная схема темновой стадии фотосинтеза
ВОПРОСЫ К ГЭКУ ПО ФЗР 2013.ppt