водный режим-2.ppt
- Количество слайдов: 30



ВОДНЫЙ ОБМЕН-2

Движущие силы водного потока от почвы через растение в атмосферу Водный баланс растения Движущие силы водного потока от почвы через растение в атмосферу включают три типа градиентов: гидростатического давления ( р), водного потенциала ( w) и концентрации водяных паров ( сwv). Градиенты гидростатического давления ( р), влияющие на транспорт воды в растении, имеются на границе почвенного воздуха и клеток корня, а также между сосудами ксилемы и окружающими тканями. Из окончаний сосудов ксилемы вода поступает в клеточную стенку клеток мезофилла, где она испаряется в воздушной полости листа. Так концентрация водяных паров внутри листа обычно выше, чем в атмосфере, вода по градиенту Сwv диффузионным путем выходит из листа через устьица.

Поступлению воды в клетку корневого волоска способствует гидростатическое давление почвенного воздуха. Модель, демонстрирующая поступление воды (большие стрелки в клетку корневого волоска, как результат гидростатического давления (маленькие стрелки) почвенного воздуха.

Схема строения корня Почти вся поглощаемая растением вода поступает в него через корни. Лишь незначительное количество воды поглощают надземные части растения. Вода поглощается главным образом за счет осмотических сил, перемещаясь от участков с высоким водным потенциалом почвы к участкам с более низким водным потенциалом корня. В транспорте воды может также участвовать и микориза, особенно в старых частях корня. При недостатке влаги вода становится основным фактором, определяющим скорость и направление роста корня. При недостатке воды рост корня ориентирован по градиенту влажности, а не силы тяжести. При этом происходит резкое возрастание поглощающей поверхности за счет увеличения количества корневых волосков. Все особенности анатомии и морфологии корня связаны с его основной функцией поглощением воды и растворенных в ней минеральных веществ из почвы, а также их транспортом в надземную часть растения. В первичном строении корня различают более десяти типов тканей.

Радиальный путь воды в корне Радиальное передвижение воды в корне возможно двумя путями: через цитоплазму по плазмодесмам (симпластный поток) и по клеточным стенкам (апопластный поток). Апопластный транспорт воды быстрее, однако он обрывается на уровне эндодермы из-за непроницаемых для него поясков Каспари. Клетки эндодермы не всегда непроницаемы для воды. Во-первых, в молодых растущих клетках пояски Каспари еще Поэтому на уровне не сформированы. Во-вторых, эндодерма прерывается в эндодермы вода и ионы участках корня, где происходит заложение боковых корней. должны войти в клетку Тем не менее, массовый поток воды по апопласту через мембрану. клетки эндодермы резко ограничен.

Корневое давление В сосуды ксилемы вода поступает благодаря градиенту водного потенциала, содаваемому осмотически активными веществами в сосудах ксилемы. Эти вещества создают осмотический потенциал ( s), способствующий осмотическому транспорту воды в сосуды ксилемы. В результате этого процесса развивается гидростатическое (корневое) давление, обеспечивающее поднятие раствора по сосудам ксилемы в надземную часть растения. Механизм, обеспечивающий поднятие воды наверх за счет корневого давления, называют нижним концевым двигателем. Плач растений выделение ксилемного эксудата у деревьев весной на раневых поверхностях (березовый сок). Явление “плача” свидетельствует о значительном корневом давлении, которое в этот период у основания ствола деревьев достигает 1, 013 МПа. Если у вегетирующего растения удалить стебель, то на срезе начинает выделяться ксилемный сок, или пасока. Если на срез поместить манометр, можно измерить величину корневого давления. Гуттация процесс выделения капель жидкости на кончиках листьев при высокой влажности воздуха, когда транспирация затруднена. В таких условиях подъем воды осуществляется главным образом за счет корневого давления. Явление гуттации особенно характерно для тропиков, где можно попасть под дождь гуттирующих растений.

zeaxanthin Guttation in a plant overexpressing Le. NCED 1 Nt-aba 2 At-aba 1 ZEP violaxanthin 9’-cis-neoxanthin Zm-VP 14 NCED Le-not xanthoxin At-aba 2 ABA- aldehyde Le-flc At-aba 3 Le-sit ABA Thompson et al. , Plant Journal 23: 363 -374, 2000

Транспирация Одной из главных проблем, c которой растения столкнулись при выходе на сушу, была двойственность их требований к условиям существования в воздушной среде. С одной стороны, атмосфера является источником СО 2, необходимого для фотосинтеза. Поэтому клеткам, участвующим в фиксации углекислоты, необходим постоянный доступ к атмосферному воздуху. С другой стороны, сухость атмосферного воздуха является причиной обезвоживания растений. Так что растению одновременно приходится отвечать двум противоречащим другу требованиям: максимального поглощения СО 2 листовой поверхностью при минимальных потерях воды. В растительном организме должен существовать тонкий и динамичный механизм регулирования поглощения СО 2 и испарения воды, который бы обеспечивал максимальную продуктивность фотосинтеза при минимальных потерях воды. Процесс испарения воды надземными органами растения называют транспирацией. Различают устьичную и кутикулярную транспирацию.
 2) 3) 4) Транспирация через устьица слагается по крайней мере из")
Устьичная транспирация 1) 2) 3) 4) Транспирация через устьица слагается по крайней мере из четырех процессов: передвижение воды из сосудов в клеточные стенки клеток мезофилла; испарение с поверхности клеток мезофилла; диффузия водяных паров в полости листа; выход водяных паров в атмосферу через устьица. Площадь внутренней поверхности листа обычно на порядок выше площади его наружной поверхности. Вода в межклетниках испаряется со всех открытых участков клеток мезофилла, а затем через устьичную щель поступает в атмосферу. Лист представляет собой насыщенный влагой орган, пронизанный множеством пор, которые сообщаются друг с другом сетью разветвленных воздушных ходов. Не смотря на то, что площадь устьичных отверстий по отношению к поверхности листа составляет всего около 1%, испарение воды через них идет очень интенсивно. Это объясняет закон Стефана, который гласит, что скорость диффузии газов через малые отверстия пропорциональна не площади, а диаметру отверстия.

Устьица можно обнаружить в эпидерме всех надземных органов растения, но больше всего их на листьях. Количество устьиц варьирует в зависимости от возраста листа и условий среды и составляет от 50 до 500 на 1 мм 2. В листьях устьица могут располагаться или на обеих сторонах, или только на нижней. Устьица представляют собой отверстия (щели) в эпидерме, образованные двумя специализированными эпидермальными клетками, которые называют замыкающими. Замыкающие клетки, изменяя свою форму, вызывают открывание или закрывание устьичной щели. У многих видов растений вокруг замыкающих клеток расположены так называемые побочные клетки. В отличие от основных клеток эпидермы, они способны принимать непосредственное участие в движении устьиц.

Замыкающие клетки устьиц можно разделить на два типа: Клетки первого типа, характерные для травянистых злаков и некоторых других видов однодольных растений, например пальм, имеют гантелеобразную форму: узкие в средней части и расширенные на обоих концах. Электронные микрофотографии устьиц. а – устьице травянистых растений; б – сканирующие электронные фотографии эпидермиса лука: на верхней фотографии показано устьице на наружной поверхности листа, окруженное кутикулой, на нижней – замыкающие клетки, формирующие устьичную щель внутри полости листа. Замыкающие клетки второго типа, имеющие форму серпа с закругленными концами (почковидную форму), встречаются у всех двудольных растений, мхов, папоротников, голосеменных и многих однодольных растений.

Толщина клеточной стенки устьичных клеток неравномерна и на многих участках гораздо больше, чем у основных клеток эпидермы. У замыкающих клеток почкообразной формы дорсальная клеточная стенка тоньше и более эластичная, чем в вентральной части клетки, где клеточная стенка толще. Важная роль в регуляции формы замыкающих клеток принадлежит микрофибриллам целлюлозы, радиально ориентированным в клеточной стенке. Они выполняют роль своеобразных направляющих при движениях устьиц. Движение двух замыкающих клеток при изменении тургорного давления практически синхронно, поскольку их цитоплазма связана плазмодесмами. Радиальная ориентация микрофибрилл целлюлозы в клеточной стенке замыкающих клеток
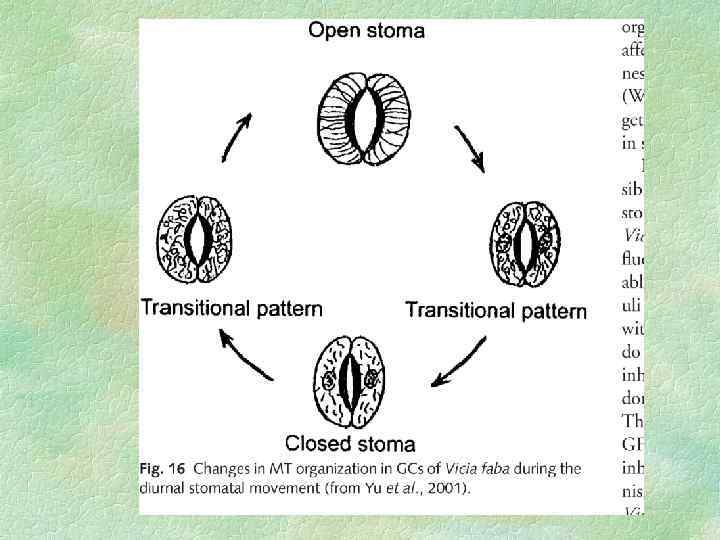

Процесс открывания устьиц начинается с поглощения замыкающими клетками осмотически активных соединений. Это индуцирует резкое падение осмотического потенциала ( s) и поток воды внутрь клеток. По мере поступления воды начинает возрастать тургорное давление и размеры клеток. За счет эластичных свойств клеточных стенок замыкающие клетки могут увеличиваться в объеме на 40 100%. Более интенсивное растяжение дорсальных участков клеточной стенки инициирует латеральное смещение ее вентральных участков и открывание щели. Закрывание устьиц индуцируется выходом осмотически активных веществ и воды из замыкающих клеток. Наиболее важное значение в механизме открывания устьиц имеет синий свет, а закрывания фитогормон абсцизовая кислота (АБК).

Открывание устьиц при освещении синим светом является очень специфической и обратимой реакцией.
. Закрывание устьиц индуцируется при экзогенной обработке")
Механизм закрывания устьиц под влиянием АБК (Schroeder, 2001). Закрывание устьиц индуцируется при экзогенной обработке листьев АБК. При обезвоживании и снижении водного потенциала растений в первую очередь регистрируется резкое повышение уровня АБК и уже после этого закрывание устьиц.
 Открытые устьица контрольного варианта, красное окрашивание – флуоресценция хлорофилла. в) Устьица, обработанные NO,")
а) Открытые устьица контрольного варианта, красное окрашивание – флуоресценция хлорофилла. в) Устьица, обработанные NO, закрыты, зеленое окрашивание – NO-специфический флуоресцентный зонд 4, 5 -диамино-флуоресцеин диацетат (DAF-2 DA). с) Устьица обработаны фитогормоном АБК.

Кутикулярная транспирация Интенсивность кутикулярной транспирации зависит от толщины кутикулы: у молодых листьев с тонкой кутикулой она составляет 50%, у зрелых листьев с мощной кутикулой 10% от всей транспирации, Схематическое изображение структуры кутикулы, покрывающей эпидермальные клетки молодых листьев и побегов в стареющих листьях кутикулярное испарение воды вновь возрастает изза разрушения кутикулы. Кутикула представляет собой трехслойную структуру. 1. Поверхность ее покрыта воском. 2. Средний слой, называемый истинным кутином, состоит из кутина, погруженного в воск. 3. Нижний слой включает кутин, воска и углеводороды, смешанные с элементами клеточной стенки.

Воска не являются полимерами. Это смесь алканов, жирных кислот, спиртов и их эфиров: СН 3 (СН 2)27 СН 3, СН 3 (СН 2)29 СН 3, СН 3 (СН 2)22 СООН, СН 3 (СН 2)24 СН 2 ОН, О СН 3 (СН 2)22 С О (СН 2)25 СН 3. Они имеют длинную алкильную цепь (25 35 углеродных атомов) и обладают (как и парафины) высокогидрофобными свойствами. Воска синтезируются в эпидермальных клетках. На поверхности клеток они часто кристаллизуются.

Кутин полимер, состоящий из гидрокси-жирных кислот, связанных между собой эфирными связями в прочную трехмерную сеть. Эти жирные кислоты могут содержать от 16 до 18 углеродных атомов, а также гидроксильные остатки и одну двойную связь.

Кутикула является внешним защитным покрытием клеток эпидермиса листьев и молодых стеблей. Она защищает растения не только от потери воды, но также представляет собой барьер для бактерий и грибов. Еще одним вторичным соединением, имеющим важное значение в водном обмене, является суберин. Это компонент клеточных стенок многих клеток растения. Он содержится в поясках Каспари клеток эндодермы корня и входит в состав оболочек клеток феллемы (пробки). Суберин состоит из жирных кислот, в составе которых имеются гидроксильные и эпокси- группы: OH CH 2 (CH 2)14 COOH Эти жирные кислоты (так же, как и в кутине) полимеризуются за счет образования эфирных связей. В отличие от кутина, в состав суберина могут входить дикарбоновые жирные кислоты (HOOC (CH 2)14 COOH ) и фенольные соединения. Кутин, суберин и ассоциированные с ними воска формируют барьер между растением и средой, защищая его от потери воды, от патогенных грибов и от бактерий. Воска отталкивают воду, что не дает спорам грибов прорастать. Некоторые грибы выделяют кутиназу, которая гидролизует кутин и облегчает проникновение гифов грибов в ткани растения.

Некоторое количество воды выделяется в результате транспирации почек и репродуктивных органов. Например, корзинки подсолнечника, коробочки мака и плоды перца транспирируют интенсивнее, чем их листья. Помимо этого вода может испаряться с поверхности ветвей и стволов древесных растений через чечевички и окружающие их слои пробки. Из-за транспирации ветвей в зимний период часто возникает обезвоживание, приводящее к гибели растений.

Передвижение воды по сосудистой системе растения Главным путем, по которому в растении транспортируются вода и минеральные вещества являются сосуды ксилемы. Эта ткань включает несколько типов мертвых вытянутых проводящих клеток, контактирующих с живыми паренхимными клетками. Транспорт воды по сосудистой системе идет по градиенту водного потенциала. Основной водопроводящей тканью сосудистых растений является ксилема. Пространственно она объединена с флоэмой, вместе с которой они образуют непрерывную проводящую систему, проходящую через все органы и ткани растения. В зависимости от происхождения и этапа развития различают первичные и вторичные проводящие ткани. Первичные проводящие ткани образуются из прокамбия, вторичные – из камбия. И первичная, и вторичная ксилема, несмотря на ряд гистологических различий, содержат водопроводящие элементы, паренхимные клетки, а также некоторые другие типы клеток, например опорные.

Специализированными клетками ксилемы, выполняющими функцию проведения воды и растворенных в ней веществ, являются трахеальные элементы, клетки которых в зрелом состоянии мертвые и имеют вытянутую форму. Для них характерна лигнифицированная, пронизанная порами клеточная стенка. Существует два типа трахеальных элементов трахеиды и членики сосудов. Они отличаются друг от друга тем, что у члеников сосудов имеются перфорации на каждом конце клетки. У трахеид же таких перфораций нет. В трахеидах передвижение воды из клетки в клетку осуществляется через поровые мембраны. Продольные ряды члеников, связанных перфорациями, чаще называют сосудами. Сосуды ксилемы похожи на полые трубки из клеточных стенок, которые расположены одна над другой. Средняя длина сосудов около 10 см, Средний диаметр сосудов от 0, 3 до 0, 5 мм. Перфорированную часть члеников сосудов называют перфорационной пластинкой. Сосуды ксилемы и трахеиды. а – схематическое изображение трахеид и члеников сосудов ксилемы: 1 – лестничная перфорационная пластинка, 2 – простая перфорационная пластинка, 3 – поры; б – сканирующая электронная фотография древесины дуба, на которой представлено два сосуда, каждый из которых состоит из двух сосудистых элементо.

У голосеменных растений функцию транспорта воды выполняют трахеиды остроконечные, вытянутые, мертвые клетки длиной 1 10 см. Трахеиды обладают сильно лигнифицированной клеточной стенкой, которая пронизана многочисленными порами. Трахеиды имеются во всех сосудистых растениях. У покрытосеменных растений они чаще встречаются в молодых тканях. В процессе эволюции специализация трахеальных элементов шла преимущественно в направлении разделения механической и проводящих функций. Примером наименее выраженной специализации являются трахеиды, совмещающие и проводящую, и механическую функции. По мере дальнейшего развития в древесине среди проводящих элементов появляются членики сосудов, более эффективные в проведении веществ, чем выполнении механических функций. В противоположность этому волокна эволюционировали, как специализированные опорные элементы. Следовательно, от примитивных трахеид идут два направления специализации, одно к сосудам, другое – к волокнам. Однако с появлением сосудов трахеиды не исчезли, а претерпели ряд изменений – они стали короче, но все же не такими короткими, как членики сосудов.

Строение окаймленной поры во время массового потока воды и его блокирования. 1 – окаймление, 2 – полость поры, 3 – замыкающая пленка поры, 4 – торус. Между соседними трахеальными элементами расположено множество пор, с помощью которых трахеиды и членики сосудов сообщаются между собой. Поры двух смежных клеток обычно располагаются друг против друга. Над порой вторичная клеточная стенка фактически прерывается. Каждая пора имеет поровую камеру. Обе камеры отделяются друг от друга тонким участком клеточной стенки замыкающей пленкой поры, или поровой мембраной. Замыкающая пленка состоит из первичной клеточной стенки и срединной пластинки. Если вторичная клеточная стенка обрывается резко у краев камеры, поры называются простыми. В том случае, когда вторичная клеточная стенка нависает над камерой поры, образуя ее окаймление, поры называют окаймленными. Поровая камера, ограниченная окаймлением, открывается в полость клетки через отверстие в окаймлении апертуру поры. В трахеидах голосеменных, в особенности у Pinaceae, замыкающая пленка, которая разделяет окаймленную пару пор, имеет утолщение, называемое торусом. Так как поровая мембрана эластична, то при определенном давлении торус смещается к одной или другой стороне окаймления, закрывая поровое отверстие. В таком положении пора не участвует в проведении веществ и называется закрытой. Т. о. окаймленные поры функционируют как клапаны, предотвращающие поступление воздуха в проводящие трахеиды, который мог бы нарушить непрерывность водяного столба.

Водный обмен у растений различных экологических групп По способности адаптироваться в различных условиях обеспеченности водой наземные растения обычно делят на 4 группы: Ксерофиты преобладают в местностях с жарким и сухим климатом и хорошо приспособлены к перенесению атмосферной и почвенной засухи. Имеют мощную кутикулу на поверхности листьев и хорошо развитую корневую систему, обеспечивающую поставку воды. Гигрофиты произрастают в условиях повышенной влажности, преимущественно атмосферной, и плохо переносят почвенную и воздушную засуху. Мезофиты по способности регулировать водный режим занимают промежуточное положение между гигрофитами и ксерофитами. Обладают тонкими механизмами регуляции устьичной и кутикулярной транспирации. К гидрофитам относят растения, для которых наилучшими условиями является полупогруженный (или полностью погруженный) в воду способ обитания.

Зависимость физиологических процессов от величиныводного потенциала у растений различных экологических групп. При уменьшении водного потенциала активизируется накопление АБК и солей растениями. При этом падает интенсивность фотосинтеза и роста клеток растяжением, снижается скорость синтеза белков и элементов клеточной стенки, уменьшается величина устьичной щели.

Значения водного потенциала и его компонентов на пути транспорта воды от почвы до атмосферы Движущей силой водного потока по сосудам вдоль растения является градиент водного потенциала, который включает как минимум 4 типа градиентов: осмотического потенциала, гидростатического давления, гравитационного потенциала и концентрации водяных паров. Несмотря на то, что водный потенциал в некоторых частях растения может быть одинаков, его компоненты ( s, p, g) могут сильно разниться. Например, водный потенциал клеточной стенки и вакуоли клеток мезофилла равен 0, 8 МПа. При этом осмотический потенциал клеточной стенки составляет 0, 2 МПа, а вакуоли 1, 1 МПа.
водный режим-2.ppt