

IRES.ppt
- Количество слайдов: 26
 Внутрішня ініціація трансляції – різноманіття механізмів та можлива роль за умов стресу та апоптозу
Внутрішня ініціація трансляції – різноманіття механізмів та можлива роль за умов стресу та апоптозу
 Класичний кеп-залежний; 2) 2) Альтернативні: 3) А.") Механізми ініціації трансляції поділяються на : 1) Класичний кеп-залежний; 2) 2) Альтернативні: 3) А. Кеп-залежні; 4) Б. Кеп-незалежні (внутрішня ініціація трансляції).
Механізми ініціації трансляції поділяються на : 1) Класичний кеп-залежний; 2) 2) Альтернативні: 3) А. Кеп-залежні; 4) Б. Кеп-незалежні (внутрішня ініціація трансляції).
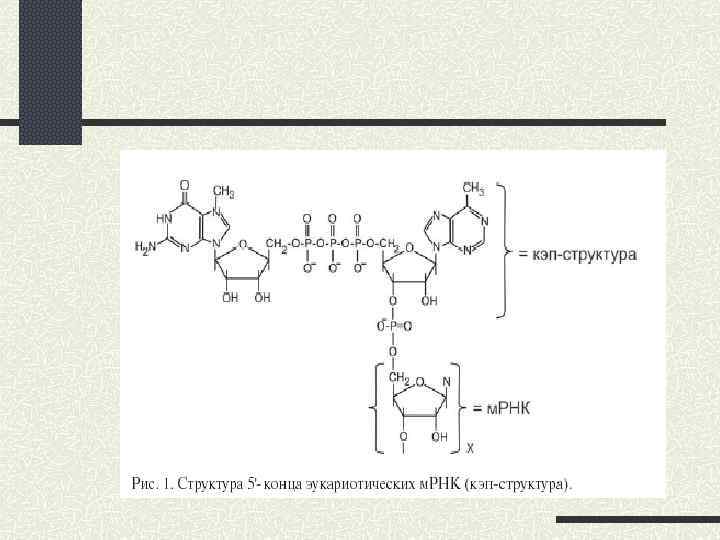
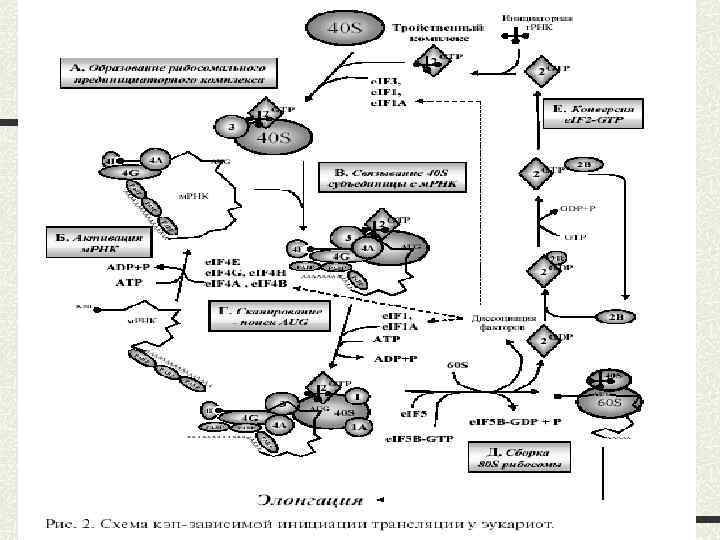
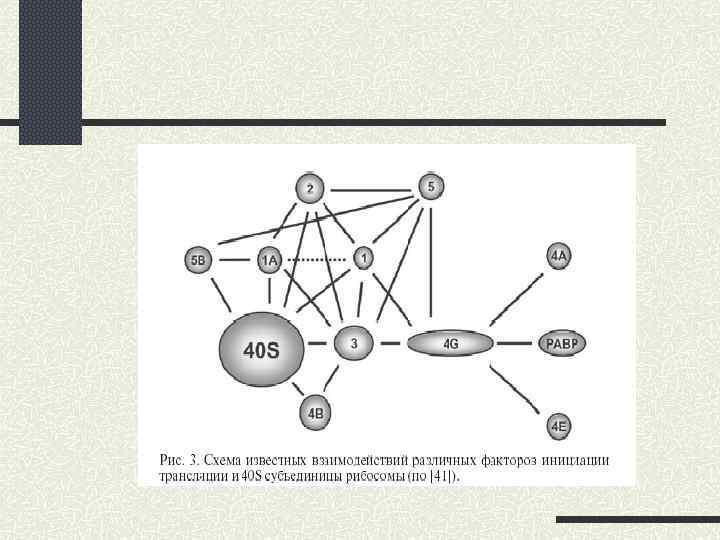
 leaky scаning (протікаюче сканування); 2) реініціація; 3) рибосомне шунтування.") Альтернативні шляхи кеп-залежної ініціації 1) leaky scаning (протікаюче сканування); 2) реініціація; 3) рибосомне шунтування.
Альтернативні шляхи кеп-залежної ініціації 1) leaky scаning (протікаюче сканування); 2) реініціація; 3) рибосомне шунтування.
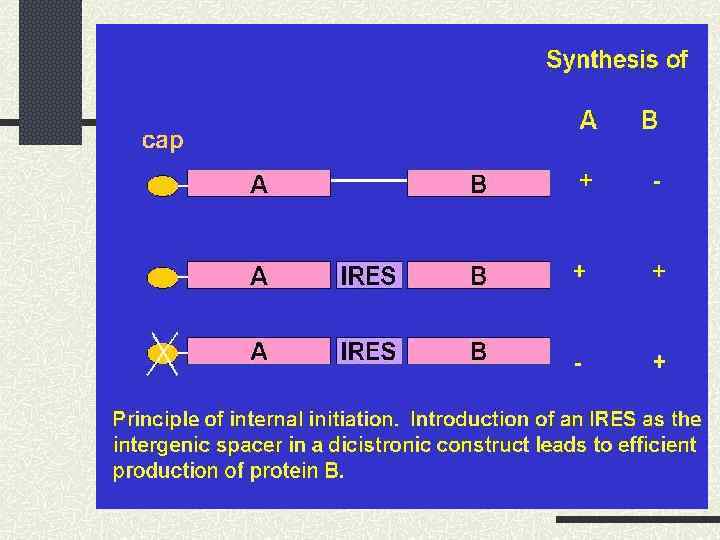
 здійснюється за участі IRES (Internal Ribosome Entry Site)") Кеп-незалежна ініціація трансляції (внутрішня ініціація трансляції) здійснюється за участі IRES (Internal Ribosome Entry Site) Це ділянка, яка входить до складу 5’-кінцевої нетрансльованої області та є місцем посадки малої субодиниці еукаріотної рибосоми. З цього місця трансляція може запускатися незалежно від кепу та пов’язаних з ним факторів трансляції
Кеп-незалежна ініціація трансляції (внутрішня ініціація трансляції) здійснюється за участі IRES (Internal Ribosome Entry Site) Це ділянка, яка входить до складу 5’-кінцевої нетрансльованої області та є місцем посадки малої субодиниці еукаріотної рибосоми. З цього місця трансляція може запускатися незалежно від кепу та пов’язаних з ним факторів трансляції
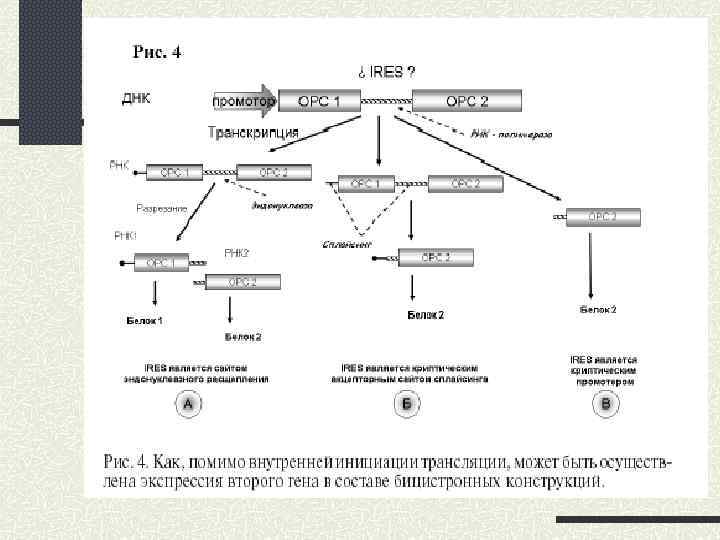
 Вторинна структура IRES
Вторинна структура IRES
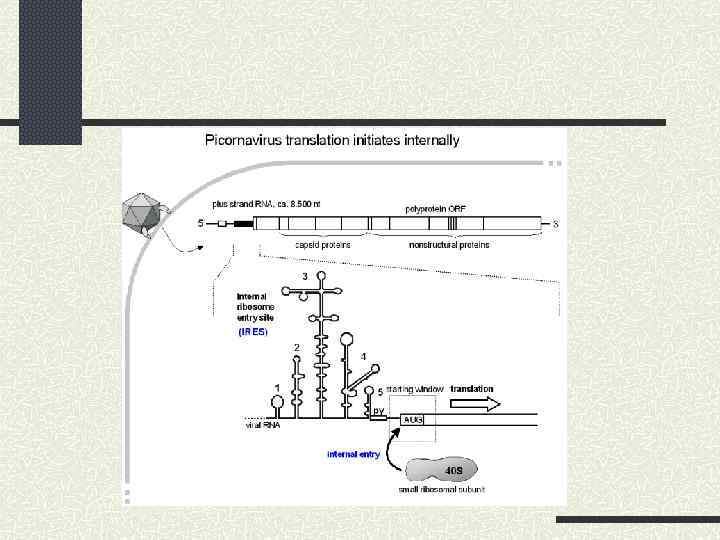


 IRES пікорнавірусів; 2) 2) IRES вірусу гепатиту С; 3) 3)") Основні типи IRES: 1) IRES пікорнавірусів; 2) 2) IRES вірусу гепатиту С; 3) 3) IRES пікорноподібних вірусів комах; 4) 4) Poly – (A) – IRES: А. IRES тобамовірусів; Б. Cdk p 110 ; 5) 5) Інші клітинні IRES.
Основні типи IRES: 1) IRES пікорнавірусів; 2) 2) IRES вірусу гепатиту С; 3) 3) IRES пікорноподібних вірусів комах; 4) 4) Poly – (A) – IRES: А. IRES тобамовірусів; Б. Cdk p 110 ; 5) 5) Інші клітинні IRES.
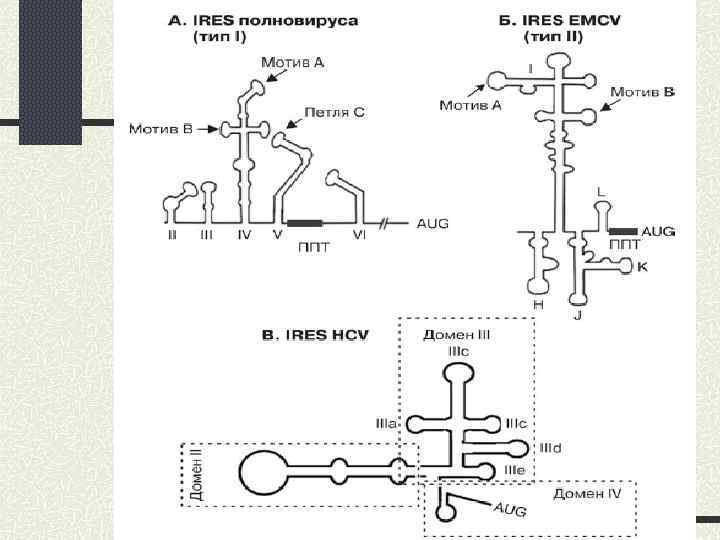
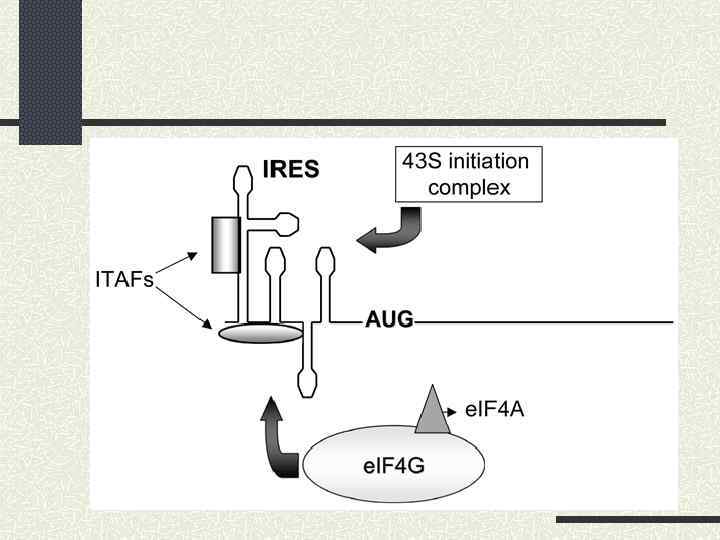
 Значна кількість поліпіримідинових ділянок 2) Використання стратегій внутрішньої посадки рибосоми") Особливості IRES пікорнавірусів: 1) Значна кількість поліпіримідинових ділянок 2) Використання стратегій внутрішньої посадки рибосоми та сканування 3) Використання типових факторів трансляції, крім e. IF 4 Е та РАВР 4) Ключовим моментом є зв’язування з IRES e. IF 4 G 5) У процесі беруть участь ITAFs (IRES – trans-acting factors)
Особливості IRES пікорнавірусів: 1) Значна кількість поліпіримідинових ділянок 2) Використання стратегій внутрішньої посадки рибосоми та сканування 3) Використання типових факторів трансляції, крім e. IF 4 Е та РАВР 4) Ключовим моментом є зв’язування з IRES e. IF 4 G 5) У процесі беруть участь ITAFs (IRES – trans-acting factors)
 4 домени у вторинній структурі 2) IRES зв’язується") Особливості IRES вірусу гепатиту С: 1) 4 домени у вторинній структурі 2) IRES зв’язується 40 S без участі факторів ініціації та потрійного комплексу, вони необхідні лише приєднанні 60 S 3) Розплітання вторинної структури РНК не відбувається 4) При зв’язуванні IRES 40 S-субчастинкою старт-кодон опиняється одразу в Р-сайті рибосоми, тому м. РНК не сканується. Механізм даного явища невідомий.
Особливості IRES вірусу гепатиту С: 1) 4 домени у вторинній структурі 2) IRES зв’язується 40 S без участі факторів ініціації та потрійного комплексу, вони необхідні лише приєднанні 60 S 3) Розплітання вторинної структури РНК не відбувається 4) При зв’язуванні IRES 40 S-субчастинкою старт-кодон опиняється одразу в Р-сайті рибосоми, тому м. РНК не сканується. Механізм даного явища невідомий.
 м. РНК біцистронні, обидві рамки зчитування транслюються за") Особливості IRES пікорнаподібних вірусів комах: 1) м. РНК біцистронні, обидві рамки зчитування транслюються за участі IRES, розміщений між цистронами 2) Старт-кодони – CUU, CAA, GCU, а ключовий елемент IRES – псевдовузол розміщений безпосередньо перед старткодоном 3) Є індукторами псевдотранслокації
Особливості IRES пікорнаподібних вірусів комах: 1) м. РНК біцистронні, обидві рамки зчитування транслюються за участі IRES, розміщений між цистронами 2) Старт-кодони – CUU, CAA, GCU, а ключовий елемент IRES – псевдовузол розміщений безпосередньо перед старткодоном 3) Є індукторами псевдотранслокації
 Механізм ініціації трансляції м. РНК пікорнаподібних вірусів комах Просторово складний IRES взаємодіє з 40 S-субчастинкою так, що триплет CCU разом з шпилькою IRES опиняється в Р-сайті, а GCU, що йде за ним, знаходиться в А-сайті 40 Sсубчастки. Петля шпильки IRES комплементарна CCU та утворюється псевдовузол, імітуючи кодон-антикодонову взаємодію ініціаторної т. РНК. Далі на CCU-кодоні утворюється комплекс 80 S-рибосоми. До А-сайта GCU приєднується Глу-т. РНК, далі відбувається псевдотранслокація – витіснення з Р-сайту IRES, що каталізується самим IRES, та перехід у Р-сайт GCU –старткодону. Реакція від GTP не залежить та демонструє роль IRES як рибозиму.
Механізм ініціації трансляції м. РНК пікорнаподібних вірусів комах Просторово складний IRES взаємодіє з 40 S-субчастинкою так, що триплет CCU разом з шпилькою IRES опиняється в Р-сайті, а GCU, що йде за ним, знаходиться в А-сайті 40 Sсубчастки. Петля шпильки IRES комплементарна CCU та утворюється псевдовузол, імітуючи кодон-антикодонову взаємодію ініціаторної т. РНК. Далі на CCU-кодоні утворюється комплекс 80 S-рибосоми. До А-сайта GCU приєднується Глу-т. РНК, далі відбувається псевдотранслокація – витіснення з Р-сайту IRES, що каталізується самим IRES, та перехід у Р-сайт GCU –старткодону. Реакція від GTP не залежить та демонструє роль IRES як рибозиму.

 Інактивація 4 Е -дефосфорилювання і зв’язування") Механізми пригнічення кеп-залежної трансляції за умов стресу: 1) Інактивація 4 Е -дефосфорилювання і зв’язування з білком 4 ЕВР 1 2) Інактивація 4 G внаслідок розщеплення протеазами 3) Інактивація е. IF 2α внаслідок фосфорилювання по Ser 51 кіназами PKR, PERK, GCN 2 4) Фосфорилювання e. IF 4 B та S 6.
Механізми пригнічення кеп-залежної трансляції за умов стресу: 1) Інактивація 4 Е -дефосфорилювання і зв’язування з білком 4 ЕВР 1 2) Інактивація 4 G внаслідок розщеплення протеазами 3) Інактивація е. IF 2α внаслідок фосфорилювання по Ser 51 кіназами PKR, PERK, GCN 2 4) Фосфорилювання e. IF 4 B та S 6.
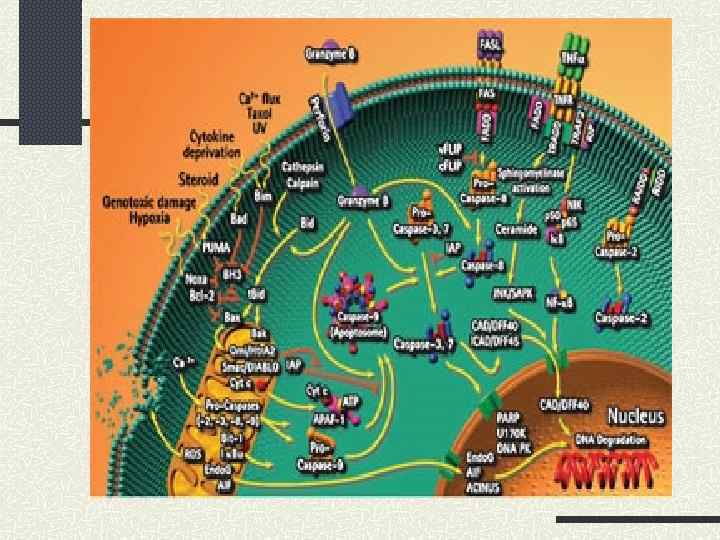
 Розщеплення") Інгібування біосинтезу білків на рівні ініціації трансляції під час апоптозу відбувається завдяки: 1) Розщеплення факторів ініціації каспазами, зокрема e. IF 4 G, e. IF 4 B, e. IF 3, e. IF 2α, 4 EBP 1 2) Експресія DAP 5/NAT 1/p 97 3) TRAIL та TNF стимулюють дефосфорилювання 4 EBP 1 та активують кіназу PKR, яка фосфорилює e. IF 2α 4) Активація JNK та p 38 -SAPK 5) Пригнічення активності кінази m. TOR, що веде до зниження рівня фосфорилювання 4 EBP 1 6) Розщеплення каспазами S 6 p 70 – кінази, яка фосфорилює S 6 – білок рибосоми
Інгібування біосинтезу білків на рівні ініціації трансляції під час апоптозу відбувається завдяки: 1) Розщеплення факторів ініціації каспазами, зокрема e. IF 4 G, e. IF 4 B, e. IF 3, e. IF 2α, 4 EBP 1 2) Експресія DAP 5/NAT 1/p 97 3) TRAIL та TNF стимулюють дефосфорилювання 4 EBP 1 та активують кіназу PKR, яка фосфорилює e. IF 2α 4) Активація JNK та p 38 -SAPK 5) Пригнічення активності кінази m. TOR, що веде до зниження рівня фосфорилювання 4 EBP 1 6) Розщеплення каспазами S 6 p 70 – кінази, яка фосфорилює S 6 – білок рибосоми
Фосфорилювання e. IF 2α кіназою PKR,") Орієнтовна послідовність процесів інгібування трансляції під час апоптозу: 1)Фосфорилювання e. IF 2α кіназою PKR, дефосфорилювання 4 EBP 1 2) Розщеплення e. IF 4 G 3) Розщеплення e. IF 4 B і e. IF 3, дефосфорилювання e. IF 4 Е, S 6 4) Розщеплення каспазами e. IF 2β, наслідком чого є пізня реактивація апарату трансляції
Орієнтовна послідовність процесів інгібування трансляції під час апоптозу: 1)Фосфорилювання e. IF 2α кіназою PKR, дефосфорилювання 4 EBP 1 2) Розщеплення e. IF 4 G 3) Розщеплення e. IF 4 B і e. IF 3, дефосфорилювання e. IF 4 Е, S 6 4) Розщеплення каспазами e. IF 2β, наслідком чого є пізня реактивація апарату трансляції
 Дякую за увагу!!!
Дякую за увагу!!!