Метаболизм углеводов.pptx
- Количество слайдов: 23



ВНУТРИКЛЕТОЧНАЯ ЛОКАЛИЗАЦИЯ УГЛЕВОДНОГО МЕТАБОЛИЗМА. ПОДГОТОВИЛА: СТУДЕНТКА 3 КУРСА, 5 ГРУППЫ, ПЫЖИВ ВЕРОНИКА

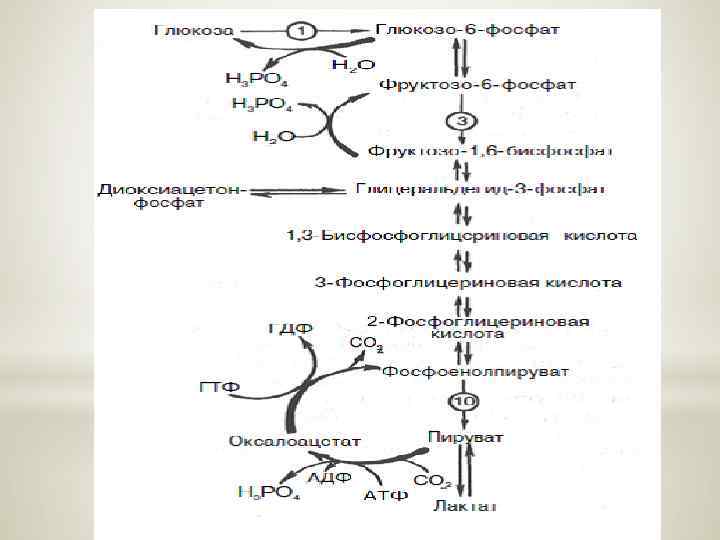
Гликолиз – это последовательность ферментативных реакций, приводящих к превращению глюкозы в пируват с одновременным образованием АТФ

Анаэробный гликолиз – сложный ферментативный процесс распада глюкозы, протекающий в тканях человека и животных без потребления кислорода. Конечным продуктом гликолиза является молочная кислота. В процессе гликолиза образуется АТФ. Суммарное уравнение гликолиза можно представить следующим образом:

Процесс гликолиза катализируется одиннадцатью ферментами, большинство из которых выделено в гомогенном, кристаллическом или высокоочищенном виде.

Биологическое значение процесса гликолиза заключается прежде всего в образовании богатых энергией фосфорных соединений. На первых стадиях гликолиза затрачиваются 2 молекулы АТФ (гексокиназная и фосфофруктокиназная реакции). На последующих образуются 4 молекулы АТФ (фосфоглицераткиназная и пируваткиназная реакции). Таким образом, энергетическая эффективность гликолиза в анаэробных условиях составляет 2 молекулы АТФ на одну молекулу глюкозы.

Глюконеогенез – синтез глюкозы из неуглеводных продуктов. Такими продуктами или метаболитами являются в первую очередь молочная и пировиноградная кислоты, так называемые гликогенные аминокислоты, глицерол и ряд других соединений. Иными словами, предшественниками глюкозы в глюконеогенезе может быть пируват или любое соединение, превращающееся в процессе катаболизма в пируват или один из промежуточных продуктов цикла трикарбоновых кислот.

Большинство стадий глюконеогенеза представляет собой обращение реакции гликолиза. Только 3 реакции гликолиза (гексокиназная, фосфофруктокиназная и пируваткиназная) необратимы, поэтому в процесс глюконеогенеза на 3 этапах используются другие ферменты. Рассмотрим путь синтеза глюкозы из пирувата. Образование фосфоенолпирувата из пирувата. Синтез фосфоенолпирувата осуществляется в несколько этапов. Первоначально пируват под влиянием пируваткарбоксилазы и при участии СО 2 и АТФ карбоксилируется с образованием оксалоацетата:

Затем оксалоацетат в результате декарбоксилирования и фосфорилирования под влиянием фермента фосфоенолпируваткарбоксилазы превращается в фосфоенолпируват. Донором фосфатного остатка в реакции служит гуанозинтрифосфат (ГТФ): Установлено, что в процессе образования фосфоенолпирувата участвуют ферменты цитозоля и митохондрий.

Первый этап синтеза протекает в митохондриях. Пируваткарбоксилаза, которая катализирует эту реакцию, является аллостерическим митохондриальным ферментом. В качестве аллостерического активатора данного фермента необходим ацетил-Ко. А. Мембрана митохондрий непроницаема для образовавшегося оксалоацетата. Последний здесь же, в митохондриях, восстанавливается в малат:

Реакция протекает при участии митохондриальной НАД-зависимой малатдегидрогеназы. В митохондриях отношение НАДН/НАД+ относительно велико, в связи с чем внутримитохондриальный оксалоацетат легко восстанавливается в малат, который легко выходит из митохондрии через митохондриальную мембрану. В цитозоле отношение НАДН/НАД+ очень мало, и малат вновь окисляется при участии цитоплазматической НАД-зависимой малатдегидрогеназы: Дальнейшее превращение оксалоацетата в фосфоенолпируват происходит в цитозоле клетки.

Превращение фруктозо-1, 6 -бисфосфата во фруктозо-6 -фосфат. Фосфоенолпируват, образовавшийся из пирувата, в результате ряда обратимых реакций гликолиза превращается во фруктозо-1, 6 бисфосфат. Далее следует фосфофруктокиназная реакция, которая необратима. Глюконеогенез идет в обход этой эндергонической реакции. Превращение фруктозо-1, 6 -бисфосфата во фруктозо-6 -фосфат катализируется специфической фосфатазой:

Образование глюкозы из глюкозо-6 -фосфата. В последующей обратимой стадии биосинтеза глюкозы фруктозо-6 -фосфат превращается в глюкозо-6 -фосфат. Последний может дефосфорилироваться (т. е. реакция идет в обход гексокиназной реакции) под влиянием фермента глюкозо-6 -фосфатазы:

АЭРОБНЫЙ МЕТАБОЛИЗМ ПИРУВАТА Клетки, недостаточно снабжаемые кислородом, могут частично или полностью существовать за счет энергии гликолиза. Однако большинство животных и растительных клеток в норме находится в аэробных условиях и свое органическое ≪топливо≫ окисляет полностью до СО 2 и Н 2 О. В этих условиях пируват, образовавшийся при расщеплении глюкозы, не восстанавливается до лактата, а постепенно окисляется до СО 2 и Н 2 О в аэробной стадии катаболизма, при этом первоначально происходит окислительное декарбоксилирование пирувата с образованием ацетил-Ко. А. Окисление пирувата до ацетил-Ко. А происходит при участии ряда ферментов и коферментов, объединенных структурно в мультиферментную систему, получившую название ≪пируватдегидрогеназный комплекс≫.
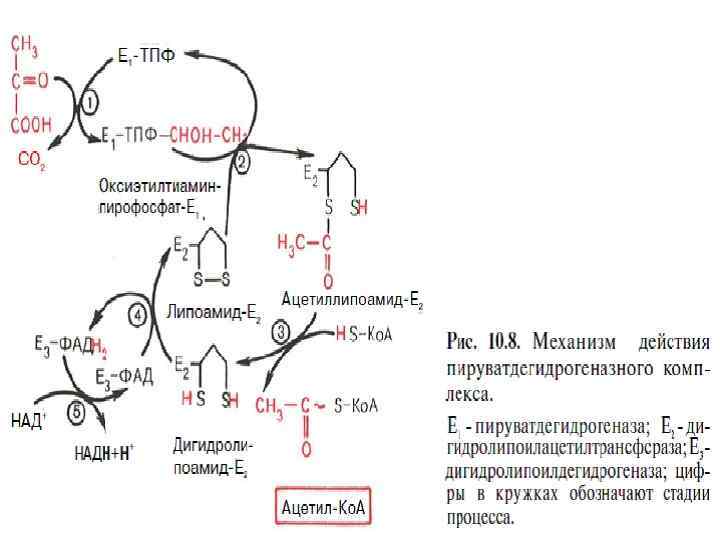

Процесс окислительного декарбоксилирования пирувата происходит в матриксе митохондрий. В нем принимают участие (в составе сложного мультиферментного комплекса) 3 фермента (пируватдегидрогеназа, дигидролипоилацетилтрансфераза, дигидролипоилдегидрогеназа) и 5 коферментов (ТПФ, амид липоевой кислоты, коэнзим А, ФАД и НАД), из которых три относительно прочно связаны с ферментами (ТПФ-E 1, липоамид -Е 2 и ФАД-Е 3), а два – легко диссоциируют (HS-Ko. A и НАД). Суммарную реакцию, катализируемую пируватдегидрогеназным комплексом, можно представить следующим образом: Реакция сопровождается значительным уменьшением стандартной свободной энергии и практически необратима. Образовавшийся в процессе окислительного декарбоксилирования ацетил-Ко. А подвергается дальнейшему окислению с образованием СО 2 и Н 2 О.
 Цикл трикарбоновых кислот впервые был открыт английским биохимиком Г.")
ЦИКЛ ТРИКАРБОНОВЫХ КИСЛОТ (ЦИКЛ КРЕБСА) Цикл трикарбоновых кислот впервые был открыт английским биохимиком Г. Кребсом. Он первым постулировал значение данного цикла для полного сгорания пирувата, главным источником которого является гликолитическое превращение углеводов. В дальнейшем было показано, что цикл трикарбоновых кислот является тем центром, в котором сходятся практически все метаболические пути. Таким образом, цикл Кребса – общий конечный путь окисления ацетильных групп (в виде ацетил-Ко. А), в которые превращается в процессе катаболизма большая часть органических молекул, играющих роль ≪клеточного топлива≫: углеводов, жирных кислот и аминокислот.
 – цитрат-синтаза; 2) – аконитат-гидротаза; 3) –")
Ферменты, катализирующие реакции цикла, обозначены цифрами: 1) – цитрат-синтаза; 2) – аконитат-гидротаза; 3) – изоцитратдегидрогеназа; 4) αкетоглутаратдегидрогеназный комплекс; 5) – сукцинил-Ко. Асинтетаза; 6) сукцинатдегидрогеназа; 7) – фумараза; 8) - малатдегидрогеназа

ПЕНТОЗОФОСФАТНЫЙ ПУТЬ ОКИСЛЕНИЯ УГЛЕВОДОВ Открытие пути прямого окисления углеводов, или, как его называют, пентозофосфатного цикла, принадлежит О. Варбургу, Ф. Липману, Ф. Дикенсу и В. А. Энгельгарду. Расхождение путей окисления углеводов – классического (цикл трикарбоновых кислот, или цикл Кребса) и пентозофосфатного – начинается со стадии образования гексозомонофосфата. Если глюкозо-6 -фосфат изомеризуется во фруктозо-6 -фосфат, который фосфорилируется второй раз и превращается во фруктозо-1, 6 -бисфосфат, то в этом случае дальнейший распад углеводов происходит по обычному гликолитическому пути с образованием пировиноградной кислоты, которая, окисляясь до ацетил-Ко. А, затем ≪сгорает≫ в цикле Кребса.

Значение этого пути в обмене веществ велико. Он поставляет восстановленный НАДФН, необходимый для биосинтеза жирных кислот, холестерина и т. д. За счет пентозофосфатного цикла примерно на 50% покрывается потребность организма в НАДФН. Другая функция пентозофосфатного цикла заключается в том, что он поставляет пентозофосфаты для синтеза нуклеиновых кислот и многих коферментов. При ряде патологических состояний удельный вес пентозофосфатного пути окисления глюкозы возрастает.

ПЕНТОЗОФОСФАТНЫЙ ПУТЬ ОКИСЛЕНИЯ УГЛЕВОДОВ

Валовое уравнение окислительной и неокислительной стадий пентозофосфатного цикла можно представить в следующем виде: Образовавшийся НАДФН используется в цитозоле на восстановительные синтезы и, как правило, не участвует в окислительном фосфорилировании, протекающем в митохондриях.

СПАСИБО ЗА ВНИМАНИЕ!
Метаболизм углеводов.pptx