Virusi_1_Vchiti_same_z_tsogo_33__33__33.pptx
- Количество слайдов: 23



ВІРУСИ Історія. Методи вивчення. Будова. Взаємодія з клітиною. Вакцини

Основні поняття • Віруси – доклітинні інфекційні агенти, що в міру простоти їх організації не здатні до повноцінного метаболізму і не здатні проявляти ознаки життєдіяльності поза клітиною господаря. • Вірусологія – розділ мікробіології, що вивчає віруси, їх будову, взаємодію з клітиною, їх класифікацію й етіологію захворювань викликаних ними. • Віруси здатні вражати абсолютно всі живі об’єкти (при тому не тільки клітинні, а також й інших вірусів) • Віруси є космополітами – вони розповсюджені по всій планеті і навіть за її межами (вірусні частки знайдені у газових хвостах комет, з чим пов’язують можливість занесення невідомих раніше штамів на планету) • Також до доклітинних інфекційних агентів відносять віроїди і пріони

• Віроїди – це патогени рослин, які складаються з короткого фрагмента (кілька сотень нуклеотидів) висококомпактизованої, кільцевої, одноланцюгової РНК, не покритої білковою оболонкою, характерною для вірусів. • Пріони (від англ proteinaceous infectious particles - Білкові заразні частинки) - особливий клас інфекційних агентів, виключно білкових, (не містять нуклеїнових кислот), що викликають важкі захворювання центральної нервової системи у людини і ряду вищих тварин (т. зв «повільні інфекції» . ).

Історія вивчення вірусів • Першим досягненням вірусології є розробка вакцини проти натуральної віспи Едвардом Дженнером у 1798 році. • До винаходу Дженнера вже були спроби винотовити вакцину від віспи – для цього у хворих брали кірочки від струпів на місці пустул, розтирали їх у порошок і втирали у шкіру чи вдихали через ніс. Недоліком було те, що вірус міг бути або занадто ослабленим і не викликав необхідної дії, або ж викликав повноцінне захворювання. • Дженнер використав метод втирання вмісту пустул коров’ячої віспи, яка не є смертельною для людини , в подряпину на шкірі. Як результат перший його паціет – 8 річний хлопчик не захворів ні від привики ні від привитої йому повноцінної сильної натуральної віспи. • Натуральна віспа (лат. Variola, Variola vera) або, як її ще називали раніше, чорна віспа високозаразна вірусна інфекція, якою страждають тільки люди. Її викликають два види вірусів: Variola major (смертність 20 -40%, за деякими даними - до 90%) і Variola minor (смертність 1 -3%), які відносяться сімейства Poxviridae. Люди, які виживають після віспи, можуть частково або повністю втрачати зір, і практично завжди на шкірі залишаються численні рубці в місцях колишніх виразок.

• У 1881 році Луї Пастер став першим, хто став застосовувати практику використання лабораторних тварин для дослідження вірусу сказу. • Зрештою, раз за разом культивуючи вірус у кроликів він виявив, що сказ локалізується у нервовій тканині. Зрештою він, так і не змігши виділити збудника, таки створив вакцину від сказу (антирабічна вакцина), що являла собою шматочок нервової тканини, що був висушений у середовищі з їдкими хімікатами. Як результат вірус вбивався, але його поверхневі антигени лишались цілісними й доступними імунній системі. • Зрештою було організовано масову вакцинацію від сказу на основі цієї вакцини. • 28 вересня – всесвітній день боротьби зі сказом

• Першим хто виділи вірус був російський вчений Д. І. Івановський у 1892 році • Будучи студентом Петербурзького університету він був направлений в Україну на Бесарабію виявити причину захворювання тютюну – тютюнову мозаїку. • Виділивши сік із вражених листків він фільтрував його крізь свічку Шамберлана (керамічний бактерійний фільтр, винайдений у 1884 році). Завдяки цьому він виявив, що сік, що не містив клітин здатен викликати захворювання рослин. Зрештою він охарактеризував збудника тютюнової мозаїки фільтрівним збудником, або отрутою (вірус).

• У 1997 Лефлер і Фрош використовуючи спосіб Івановського довели схожу природу збудника ящуру. Після цього так само виділили збудників жовтої лихоманки (Рідд і Керол 1901), поліоміеліту (Ландштейнер і Поппер 1909) і багато інших. • Зрештою Туорт (1915) і Д’Еррель (1917) незалежно один від одного відкрили бактеріофагів • Хоч методи культивування тваринних клітин поза їх організмом були відомі у 1912 (Каррель), і в них вдалось культивувати віруси (Шейнгард, 1913) для вакцин віруси культивувались в основному в курячих ембріонах аж до 1952, коли Дульбеко вдосконалив метод культивації вірусів до культивації у одношаровій культурі клітин.

Надалі розрізняють 4 періоди досліджень вірусології: • 30 -40 -ві роки – організмовий рівень v. В цей період вивчався загальний вплив вірусів на організм. v. У 1937 році було відкрито вірус кліщового енцефаліту v. У 1941 році амер. Вчений Херст відкрив явище гемаглютинації • 50 роки – клітинний рівень v. Відкрито метод одношарової культури клітин Дж. Ендерсом і ін. (Нобелівська премія) v 1959 – винайшли вакцину від поліомієліту • 60 -ті роки - молекулярний рівень v. Змогли розділити віруси на молекулярні складові і вивчити структуру геному й капсиду вірусів • 70 -ті роки – субмолекулярний рівень v. Секвенували спадкову інформацію багатьох вірусів, вивчили властивості й амінокислотний склад їх білків, дослідили вірусні ферменти. v 1970 – Балтімор і Темін незалежно один від одного відкрили зворотну транскриптазу вірусів. v 1976 р – американський вчений Й. Гайдушенко отримав новелівську премію за дослідження пріонного захворювання куру v. В той же рік ще одна нобелівська премія була присуджена вченому Г. Бламбергу за відкриття антигену вірусу гепатиту, за яким можна виявити даний вірус у людини

Походження вірусів • В загальному є 3 теорії виникнення вірусів. ØПринцип регресивної еволюції. Згідно з цією теорією віруси походять від бактерій, що настільки спростились, що втратили здатність жити за межами інших клітин. Ця теорія не здатна пояснити відсутність всласних систем синтезу вірусів і велику різноманітність геномів вірусів загалом, тому ця теорія не підтримується більшістю вірусологів. ØТеорія походження від древніх доклітинних організмів – протобіонтів. Ця теорія так само себе не виправдовує через те що не пояснює ті ж аспекти, що і перша ØТеорія автономізації мобільних генетичних елементів або теорія «скажених генів» . Згідно із цією теорією деякі гени набули здатності автономно розмножуватись і переміщатись між клітинами. Ця теорія підтримується найбільшою частиною сучасних вірусологів. Щоправда ця теорія не пояснює появу у вірусів РНК, в якості носія генетичної інформації.

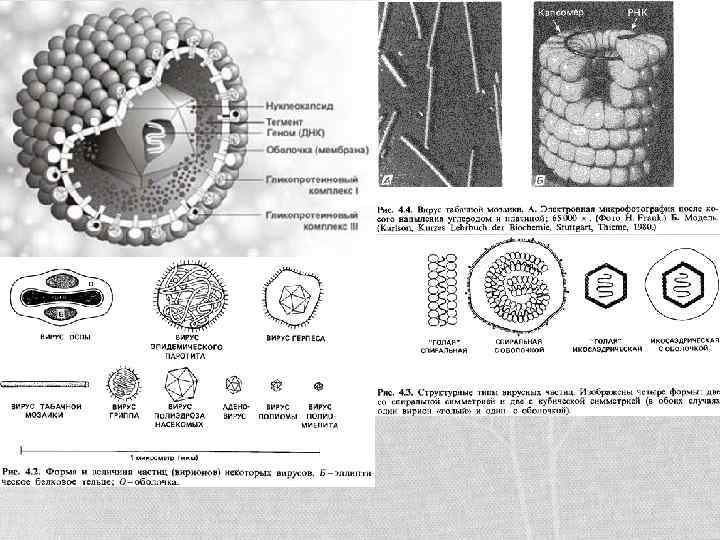
Склад вірусів • Вірусні частки завжди складаються із білкової оболонки і нуклеїнової кислоти. В якості додаткових елементів їх структури може бути ліпопротеїнова оболонка і додаткові ферменти, що в уже синтезованому вигляді присутні у вірусній частці. • За складом віруси поділяють на прості і складні: • Прості віруси це віруси що складаються виключно із нуклеїнової кислоти і білку. • Складні віруси містять додаткові оболонки і включають в своїй структурі елементи полісахаридних ланцюгів і ліпідів. Також вони здатні нести цілі органоїди клітин власників (рибосоми у аренавірусів)

Білки вірусів • В супереч поширеному уявленню білки капсиду не є інертними. Це уявлення виникло внаслідок вивчення структури ВТМ, що складається лише з одного типу білка, що утворює чохол для РНК. Насправді ж функції капсиду виходять далеко за межі функціонування лише як футляр. Завдяки тому що білки капсиду по суті мають субодиничну структуру (можуть розділятись на окремі білки) в одному капсиді може бути до кількох різних за функціями ферментів, що проявляють свою роль в реалізації геному вірусу. Так, наприклад деякі білки капсиду після можифікації у клітині чи просто зміні просторової організації здатні модифікувати кінці нуклеїнових кислот віруса, проводити транскрипцію і реплікацію геному віруса, забезпечувати його вбудовування у геном клітини хазяїна. У випадку простих вірусів білки капсиду також відіграють роль проникнення у клітини хазяїна. У складних білків за це відповідають білки суперкапсиду.

Самозбирання папілома-вірусу

Геном вірусів • Віруси можуть містити як ДНК, так і РНК, в якості носія генетичної інформації. При цьому вони бувають, цілісні чи фрагментовані, а також одноланцюгові і дволанцюгові. • Розрізняють 2 типи структури генетичного матеріалу в структурі вірусів. Це так звані «плюс» , так і «мінус» нуклеїнові кислоти. ØПлюс ланцюги – це ланцюги ДНК з яких безпосередньо зразу можна проводити транскрипцію, чи РНК, які можуть одразу відігравати роль м. РНК. ØМінус ланцюги – це ланцюги ДНК чи РНК, до яких треба досинтезувати комплементарний ланцюг, щоб реалізувати генетичну інформацію віруса. • Також окремо варто виділити, що вірусні нуклеїнові кислоти можуть мати повтори на кінцях ланцюгів, для вбудовування в геном хазяїна, і також самі нуклеїнові кислоти можуть бути модифікованими на кінцях ланцюгів для того, що геном віруса обійшов систему захисту клітини – систему рестрикції

Класифікація структур вірусів • Вирізняють п’ять рівнів: ØСубодиниця – специфічним чином укладений білок. ØСтруктурний елемент - кілька субодиничних білків, що формують хімічно довершений комплекс, що володіє якимись функціями. ØКапсид – структурно завершений футляр для генетичної інформації віруса. ØНуклеокапсид – комплекс капсиду із нуклеїновою кислотою, що являє собою остаточно упаковану форму вірусного генетичного матеріалу. Може бути підструктурою у випадку складних вірусів ØВіріон – остаточно сформована вірусна частка.

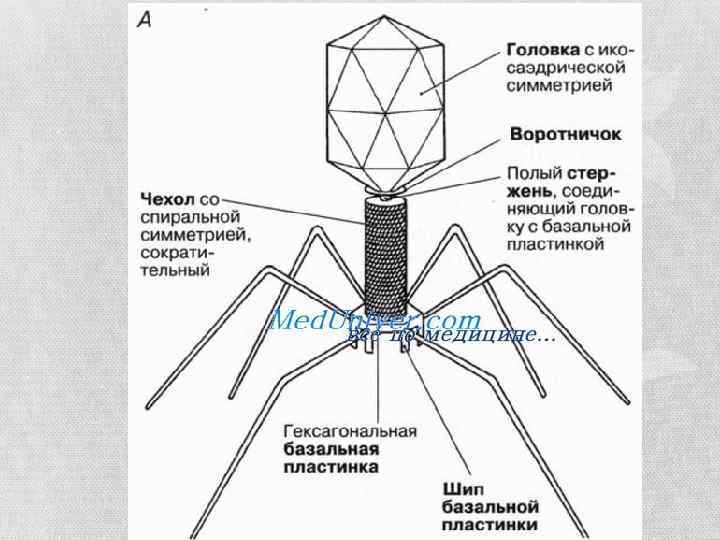
Побудова капсидів • Є 3 типи побудови вірусних часток: ØПаличкоподібний чи нитчастий – коли вірусна частинка має форму палички. В ньому нуклеїнова кислота згорнута у спіраль і з’єднана з білками капсиду. Прикладом є ВТМ і філовіруси (Ебола, Марбург) ØІзометричний, сферичний, кулястий – тип побудови, коли вірусна частка має форму правильного багатогранника. В даному випадку форма упакування вірусу не має значення для побудови білкового капсиду. В деяких випадках ДНК віруса може бути з’єднана із гістонами. Тобто нуклеокапсид може не утворюватись, а загалом зв’язок між нуклеїновою кислотою і капсидом є набагато гнучкішим і слабшим. ØКомплексний – даний тип побудови включає в собі елементи попередніх двох типів побудови і характерний для бактеріофагів.

Взаємодія з клітиною • Включає в себе стадії адсобції, проникнення, експресії та реплікації геному, збирання вірусних часток і вихід із клітини. • Може завершуватись смертю клітини одразу в момент виходу з неї великої кількості вірусних часток чи перетворенням клітини на вірусну фабрику і поступовим відбруньковуванням вірусних часток з поверхні клітини. • Ще одним результатом взаємодії вірусу з клітиною є лізогенізація – перехід віруса у латентний стан у клітині до моменту набування клітиною необхідної кількості ресурсів для того, щоб вірус ефективно відтворився і покинув клітину. При цьому вірусний геном так само контролює процеси життєдіяльності клітини, як і при звичайному розвитку подій але вірусні частки не формуються. Даний варіант взаємодії розвивається у деяких бактеріофагів.

Адсорбція • Адсорбція – процес прикріплення вірусної частки до поверхні клітини для наступного проникнення вірусу вглиб клітини. • Спочатку відбувається завдяки взаємодії заряду на поверхні клітини і вірусної частки. • У деяких вірусів також відбувається реакція із специфічними рецепторами на поверхні клітини. При цьому рецептор опосередкована взаємодія не є дуже міцною, тому для ефективного закріплення віруса необхідно утворити багато зв’язків із рецепторами на поверхні клітини. В деяких випадках кількість таких зв’язків доходить до 3 тисяч на одну вірусну частку. З цим також пов’язане явище спеціалізації віруса – зрештою вірус може настільки пристосуватись до рецепторів на клітинах одного виду організмів, що стане нездатним заражати інші види організмів. Білки, що відповідають за таку взаємодію називають білками закріплення.

Проникнення в клітину • В основному іде 2 шляхами – рецептор-опосередкованим ендоцитозом і злиттям з мембраною. • Рецептор опосередкований шлях реалізується через той принцип, що при контакті речовини із специфічним до нього рецептором відбувається ендоцитоз. Завдяки цьому віруси навчились проникати в клітину шляхом виклакання ендоцитозу, подразнюючи рецептор на поверхні клітини. Зрештою, опиняючись у фагосомі віріон під дією знаження р. Н прикріплюється до мембрани фагосоми і вивільняє генетичний матеріал всередину цитоплазми. • Злиття з мембраною відбувається у складних вірусів, коли ліпопротеїнова мембрана віруса зливається із плазматичною мембраною клітини. Як результат в клітину попадає нуклеопротеїд, яких згодом «роздягається» із звільненням вірусного генома. • Окремо варто виділити бактеріофагів, які «вколюють» клітині свої нуклеїнові кислоти.

Експресія і реплікація вірусного геному • Коли вірусний геном з деякими білками проник у цитоплазму у вірусного геному є 2 шляхи: ØВірусний генетичний матеріал починає проявляти свою активність у цитоплазмі ØВірусний геном відправляється в ядро клітини, де в свою чергу може вбудовуватись у ДНК хазяїна чи лишитись дискретним. • При цьому може відбуватись зворотна транскрипція(утворення ДНК із РНК) та синтез комплементарних ланцюгів. • Далі починається синтез ранніх білків. Ці білки вбудовуючись у органоїди клітини чи модифікуючи власні ферменти клітини передають контроль над всіма процесами в клітині геному віруса. Після цього починається фаза синтезу пізніх білків – елементів капсиду. Тоді ж починається активна реплікація геному віруса.

Фаза збирання вірусних частинок і виходу із клітини • В результаті синтезу багатьох молекул капсомерів і геном віруса починається процес самозбирання віруса. При цьому характерним є те, що помилково вбудовані і пошкоджені білки заміняються нормальними без видимих потужніх способів контролю цього процесу. Зрештою, коли процес самозбирання завершився вірусні частки покидають клітину. • Вихід із клітини відбувається 2 шляхами: ØВибухоподібно(розривається мембрана клітини) із негайною смертю ØВідбруньковуванням (плавне відбруньковування) із виживанням клітини хазяїна і перетворенням її на вірусну фабрику.
 § Інактивовані вакцини")
vІ покоління Види вакцин § «Живі» вакцини (містять живі ослаблені мікроорганізми) § Інактивовані вакцини vІІ покоління § Вакцини підтипові або спліт-вакцини (Вакцини з ізольованими антигенами ) vІІІ покоління § Рекомбінантні вакцини v. IV покоління § ДНК-вакцини § Ідіотипові вакцини v. Комбіновані вакцини
Virusi_1_Vchiti_same_z_tsogo_33__33__33.pptx