

ВНД_Память и обуч 2012.ppt
- Количество слайдов: 67
 Виды памяти и их локализация в мозге
Виды памяти и их локализация в мозге
 Образная зрительная П. : данные эл-ии; феномены ‘flash-back’ и ‘déjà vu’
Образная зрительная П. : данные эл-ии; феномены ‘flash-back’ и ‘déjà vu’
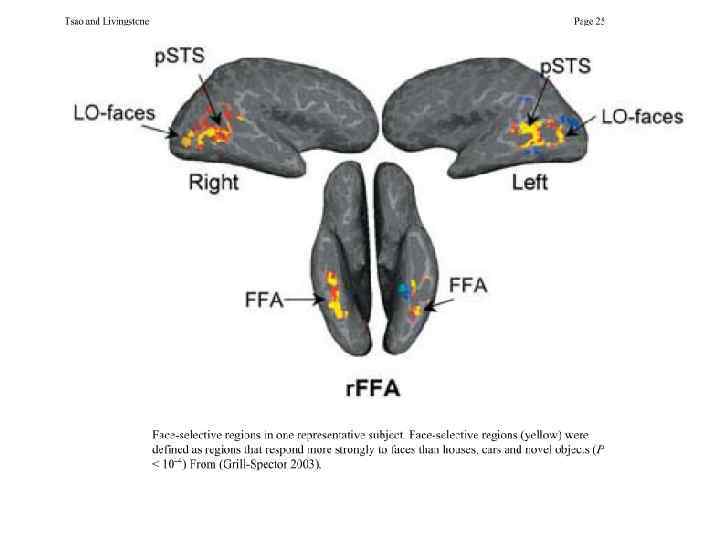
 Нейроны зрительной памяти
Нейроны зрительной памяти
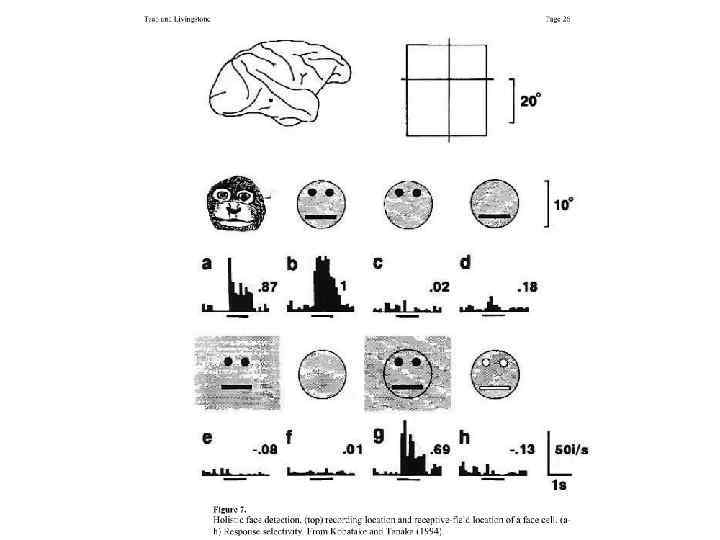
 Доказательство существования нейронов зрительной памяти у человека Эксперимент состоит из двух частей. Вначале испытуемому показывают разные объекты и смотрят, на что активируется тот или иной конкретный нейрон (височная область). А затем после окончания эксперимента его просят вспомнить (перечислить), что он видел, и при этом опять отслеживают активность того же нейрона. Обнаружили, что нейрон селективно разряжается как при запоминании, так и при воспоминании строго одного объекта. Далее, оказалось, что когда человек вспоминал про тот или иной объект, у него вначале активировался соответствующий нейрон памяти, а только потом с небольшой задержкой следовала речевая реакция.
Доказательство существования нейронов зрительной памяти у человека Эксперимент состоит из двух частей. Вначале испытуемому показывают разные объекты и смотрят, на что активируется тот или иной конкретный нейрон (височная область). А затем после окончания эксперимента его просят вспомнить (перечислить), что он видел, и при этом опять отслеживают активность того же нейрона. Обнаружили, что нейрон селективно разряжается как при запоминании, так и при воспоминании строго одного объекта. Далее, оказалось, что когда человек вспоминал про тот или иной объект, у него вначале активировался соответствующий нейрон памяти, а только потом с небольшой задержкой следовала речевая реакция.
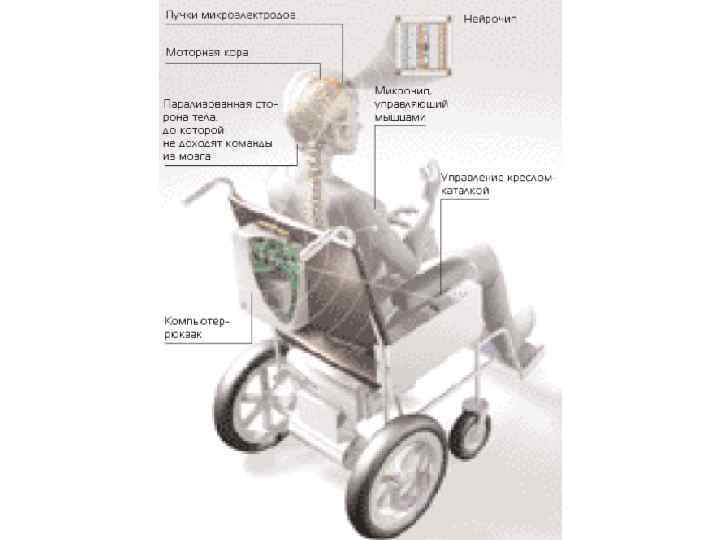
 От нейрона памяти – к интерфейсу «мозг-ЭВМ» Это открывает возможность чтения мыслей мозга (reading brain), т. е перейти от данных по нейронам памяти – к созданию интерфейса «мозг-компьютер» . Представьте, что вы возьмете активность такого нейрона у крысы, который активируется к моменту нажатия на педаль, и регистрируете сигнал с этого нейрона, когда крыса нажимает на этот рычаг, и получает пододвигающуюся к ней кормушку с подкреплением. А затем сигнал с этого нейрона запустите на кормушку еще до того, как животное нажмет на рычаг. У нас получится парадоксальная ситуация, когда животное собирается нажать на рычаг, чтобы получить подкрепление в кормушке, и, еще не нажав, получает подкрепление. То есть, то, что она хотела, опережает через внешний контур управления, созданный экспериментатором, то, что она должна сделать для этого. Крысы начинают вести себя в этой ситуации удивительным образом. Они некоторое время механически продолжают нажимать на рычаг, хотя кормушка уже поступила. Но через некоторое время они соображают, в чем дело, и начинают запускать передвижение кормушки с подкреплением просто активностью своего мозга, уже перестав подбегать к педали. Животные начинают мысленно управлять внешним устройством через электронный интерфейс. Это принцип был положен в основу так называемых мозг-машинных или мозг-компьютерных интерфейсов.
От нейрона памяти – к интерфейсу «мозг-ЭВМ» Это открывает возможность чтения мыслей мозга (reading brain), т. е перейти от данных по нейронам памяти – к созданию интерфейса «мозг-компьютер» . Представьте, что вы возьмете активность такого нейрона у крысы, который активируется к моменту нажатия на педаль, и регистрируете сигнал с этого нейрона, когда крыса нажимает на этот рычаг, и получает пододвигающуюся к ней кормушку с подкреплением. А затем сигнал с этого нейрона запустите на кормушку еще до того, как животное нажмет на рычаг. У нас получится парадоксальная ситуация, когда животное собирается нажать на рычаг, чтобы получить подкрепление в кормушке, и, еще не нажав, получает подкрепление. То есть, то, что она хотела, опережает через внешний контур управления, созданный экспериментатором, то, что она должна сделать для этого. Крысы начинают вести себя в этой ситуации удивительным образом. Они некоторое время механически продолжают нажимать на рычаг, хотя кормушка уже поступила. Но через некоторое время они соображают, в чем дело, и начинают запускать передвижение кормушки с подкреплением просто активностью своего мозга, уже перестав подбегать к педали. Животные начинают мысленно управлять внешним устройством через электронный интерфейс. Это принцип был положен в основу так называемых мозг-машинных или мозг-компьютерных интерфейсов.
 Researchers at the University of Pittsburgh have demonstrated a monkey controlling an advanced robotic arm by using its thoughts. The experiments were led by Dr. Andrew Schwartz, a professor of neurobiology and involved a high degree of complexity in the robotic arm, the level of control, and the intricacy of the manipulations.
Researchers at the University of Pittsburgh have demonstrated a monkey controlling an advanced robotic arm by using its thoughts. The experiments were led by Dr. Andrew Schwartz, a professor of neurobiology and involved a high degree of complexity in the robotic arm, the level of control, and the intricacy of the manipulations.

 КП и ДП – последовательно или параллельно?
КП и ДП – последовательно или параллельно?
 Роль гиппокампа и амигдалы в механизмах ДП: биохимические и нейрофизиологические механизмы
Роль гиппокампа и амигдалы в механизмах ДП: биохимические и нейрофизиологические механизмы
 Модулирующие влияния на память амигдалы
Модулирующие влияния на память амигдалы
 Connections between the hippocampus and possible declarative memory storage sites. The rhesus monkey brain is shown because these connections are much better documented in non-human primates than in humans. Projections from numerous cortical areas converge on the hippocampus and the related structures known to be involved in human memory; most of these sites also send projections to the same cortical areas. Medial and lateral views are shown, the latter rotated 180° for clarity.
Connections between the hippocampus and possible declarative memory storage sites. The rhesus monkey brain is shown because these connections are much better documented in non-human primates than in humans. Projections from numerous cortical areas converge on the hippocampus and the related structures known to be involved in human memory; most of these sites also send projections to the same cortical areas. Medial and lateral views are shown, the latter rotated 180° for clarity.
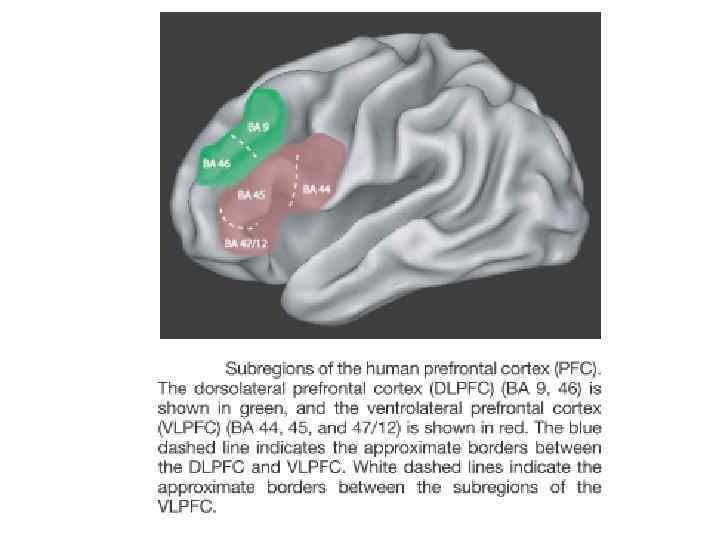
 Префронтальная кора и рабочая память. В процессе активизации устоявшихся, «консолидированных» воспоминаний ключевую роль играет медиальная префронтальная кора (МПФК). Когда мы вспоминаем события, случившиеся только что, гиппокамп активизируется сильно, а МПФК - слабо. Но если нам нужно вспомнить что-то более давнее, наблюдается обратная картина: нейроны МПФК работают очень активно, а нейроны гиппокампа - гораздо слабее. Эта подтверждено и на крысах, и на людях. Известно, также что повреждения МПФК ведут к нарушениям механизма вспоминания давних событий. .
Префронтальная кора и рабочая память. В процессе активизации устоявшихся, «консолидированных» воспоминаний ключевую роль играет медиальная префронтальная кора (МПФК). Когда мы вспоминаем события, случившиеся только что, гиппокамп активизируется сильно, а МПФК - слабо. Но если нам нужно вспомнить что-то более давнее, наблюдается обратная картина: нейроны МПФК работают очень активно, а нейроны гиппокампа - гораздо слабее. Эта подтверждено и на крысах, и на людях. Известно, также что повреждения МПФК ведут к нарушениям механизма вспоминания давних событий. .
 Американские нейробиологи предположили, что если МПФК так важна для «воспроизведения» воспоминаний, то она, наверное, должна принимать участие и в их «записи» , то есть в процессе консолидации воспоминаний (наряду с гиппокампом и некоторыми другими отделами мозга). Чтобы проверить это предположение, двум крысам имплантировали в мозг миниатюрное устройство, позволяющее следить одновременно за активностью 120 нейронов МПФК. Крысы каждый день выполняли серию заданий на ориентацию и пространственную память. За это их «награждали» электрической стимуляцией центров удовольствия. Выполнение одного задания занимало примерно 20 секунд, а задания следовали одно за другим в течение 50
Американские нейробиологи предположили, что если МПФК так важна для «воспроизведения» воспоминаний, то она, наверное, должна принимать участие и в их «записи» , то есть в процессе консолидации воспоминаний (наряду с гиппокампом и некоторыми другими отделами мозга). Чтобы проверить это предположение, двум крысам имплантировали в мозг миниатюрное устройство, позволяющее следить одновременно за активностью 120 нейронов МПФК. Крысы каждый день выполняли серию заданий на ориентацию и пространственную память. За это их «награждали» электрической стимуляцией центров удовольствия. Выполнение одного задания занимало примерно 20 секунд, а задания следовали одно за другим в течение 50
 Было обнаружено, что во время сна в первые два часа после выполнения заданий в МПФК возникают серии скоррелированных нервных импульсов, похожие на те, которые наблюдались во время «работы» , но прокрученные в ускоренном режиме. Наибольшее сходство графиков нейронной активности наяву и во сне получается в том случае, если второй график «растянуть» во времени в семь раз. Во время сна перед выполнением заданий ничего подобного не наблюдается.
Было обнаружено, что во время сна в первые два часа после выполнения заданий в МПФК возникают серии скоррелированных нервных импульсов, похожие на те, которые наблюдались во время «работы» , но прокрученные в ускоренном режиме. Наибольшее сходство графиков нейронной активности наяву и во сне получается в том случае, если второй график «растянуть» во времени в семь раз. Во время сна перед выполнением заданий ничего подобного не наблюдается.
 Subjects were") Reactivation of visual cortex during vivid remembering of visual view images. (A) Subjects were instructed to view either images of objects (houses, faces, and chairs) (left) or imagine the objects in the absence of the stimulus (right). (B) (Left) Bilateral regions of ventral temporal cortex are specifically activated during perception of houses (yellow), faces (red), and chairs (blue). (Right) When subjects recall these objects, the same regions preferentially activated during the perception of each object class are reactivated.
Reactivation of visual cortex during vivid remembering of visual view images. (A) Subjects were instructed to view either images of objects (houses, faces, and chairs) (left) or imagine the objects in the absence of the stimulus (right). (B) (Left) Bilateral regions of ventral temporal cortex are specifically activated during perception of houses (yellow), faces (red), and chairs (blue). (Right) When subjects recall these objects, the same regions preferentially activated during the perception of each object class are reactivated.
") Выработка моторных навыков ( «некогнитивная память» : St R)
Выработка моторных навыков ( «некогнитивная память» : St R)
 Межполушарная специализация в механизмах памяти Имплицитная и эксплицитная память
Межполушарная специализация в механизмах памяти Имплицитная и эксплицитная память
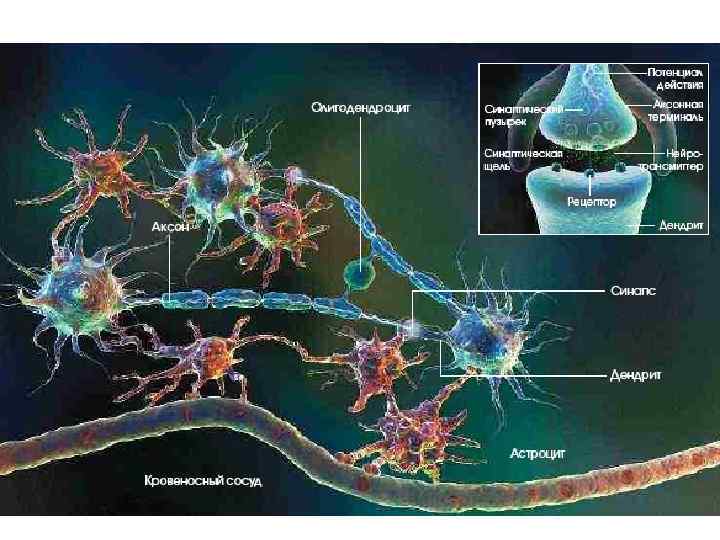
 КЛЕТОЧНЫЕ МЕХАНИЗМЫ ПАМЯТИ Синаптическая пластичность и роль глии
КЛЕТОЧНЫЕ МЕХАНИЗМЫ ПАМЯТИ Синаптическая пластичность и роль глии
 сформулировал теоретически возможный механизм хранения") Нейронные механизмы фиксации следов памяти Д. Хебб (Hebb, 1949) сформулировал теоретически возможный механизм хранения информации. С его точки зрения, основой обучения может быть совпадение во времени активности пресинаптической и постсинаптической клеток. В дальнейшем такого типа связь между клетками стала называться синапсом Хебба.
Нейронные механизмы фиксации следов памяти Д. Хебб (Hebb, 1949) сформулировал теоретически возможный механизм хранения информации. С его точки зрения, основой обучения может быть совпадение во времени активности пресинаптической и постсинаптической клеток. В дальнейшем такого типа связь между клетками стала называться синапсом Хебба.
: «Если аксон клетки А расположен настолько близко") A B Правило Хебба (1949 г. ): «Если аксон клетки А расположен настолько близко к клетке В, что может возбуждать ее, и если он многократно и непрерывно принимает участие в ее активации, то в одной или обеих клетках возникает какой-то процесс роста или метаболические изменения, и в результате эффективность клетки А как одного из активаторов клетки В возрастает» . Примечание: подразумевается, что клетка B может быть активирована каким-то другим способом, помимо синапса с клетки A – т. е. либо имеются и другие входы на клетку B, причем сильные, либо клетка B спонтанно активна.
A B Правило Хебба (1949 г. ): «Если аксон клетки А расположен настолько близко к клетке В, что может возбуждать ее, и если он многократно и непрерывно принимает участие в ее активации, то в одной или обеих клетках возникает какой-то процесс роста или метаболические изменения, и в результате эффективность клетки А как одного из активаторов клетки В возрастает» . Примечание: подразумевается, что клетка B может быть активирована каким-то другим способом, помимо синапса с клетки A – т. е. либо имеются и другие входы на клетку B, причем сильные, либо клетка B спонтанно активна.
 Больше постсинаптических рецепторов, сильнее реакция на нейромедиатор Дендритный шипик Выделение большего количества нейромедиатора Спраутинг: образование новой дополнительной терминали Утолщение и укорочение шейки шипика, что снижает его электрическое сопротивление Четыре возможных способа повышения эффективности синапса
Больше постсинаптических рецепторов, сильнее реакция на нейромедиатор Дендритный шипик Выделение большего количества нейромедиатора Спраутинг: образование новой дополнительной терминали Утолщение и укорочение шейки шипика, что снижает его электрическое сопротивление Четыре возможных способа повышения эффективности синапса
 • AMPA") Термины • NMDA = N-метил-D-аспартат – агонист кальциевых каналов, активируемых глутаматом (деполяризация) • AMPA - альфа-амино-3 -гидрокси-5 -метил-4 изоксазолпропионовая кислота • AMPA-рецепторы = ионотропные глутаматные рецепторы, активация которых приводит к деполяризации клетки и последующему срабатыванию NMDA-рецепторов • Глутаматные NMDA-рецепторы сконцентрированы в лобной коре, гиппокампе, лимбике и стриатуме • CREB-фактор = с. АМP response binding element (белок транскрипции, промотор)
Термины • NMDA = N-метил-D-аспартат – агонист кальциевых каналов, активируемых глутаматом (деполяризация) • AMPA - альфа-амино-3 -гидрокси-5 -метил-4 изоксазолпропионовая кислота • AMPA-рецепторы = ионотропные глутаматные рецепторы, активация которых приводит к деполяризации клетки и последующему срабатыванию NMDA-рецепторов • Глутаматные NMDA-рецепторы сконцентрированы в лобной коре, гиппокампе, лимбике и стриатуме • CREB-фактор = с. АМP response binding element (белок транскрипции, промотор)
 Консолидация следа памяти Один из первых вопросов, который подвергся объективным ПФ исследованиям, был вопрос о формировании следа памяти. Под влиянием работа Г. Эббингауза, два других немецких психолога Мюллер и Пильцекер (1900), задались вопросом, а что происходит на границе этого перехода от одной фазы памяти к другой? Активный ли это процесс? И они показали, что, если в момент запоминания и перехода от кратковременной в долговременную память человеку дается новая задача, которую он должен запомнить, то эта новая задача мешает запоминанию старой информации, интерферирует с ним. Они назвали это ретроградной интерференцией. Исходя из этого, они логично предположили, что в мозге во время запоминания идет активный процесс, и он требует ресурсов.
Консолидация следа памяти Один из первых вопросов, который подвергся объективным ПФ исследованиям, был вопрос о формировании следа памяти. Под влиянием работа Г. Эббингауза, два других немецких психолога Мюллер и Пильцекер (1900), задались вопросом, а что происходит на границе этого перехода от одной фазы памяти к другой? Активный ли это процесс? И они показали, что, если в момент запоминания и перехода от кратковременной в долговременную память человеку дается новая задача, которую он должен запомнить, то эта новая задача мешает запоминанию старой информации, интерферирует с ним. Они назвали это ретроградной интерференцией. Исходя из этого, они логично предположили, что в мозге во время запоминания идет активный процесс, и он требует ресурсов.
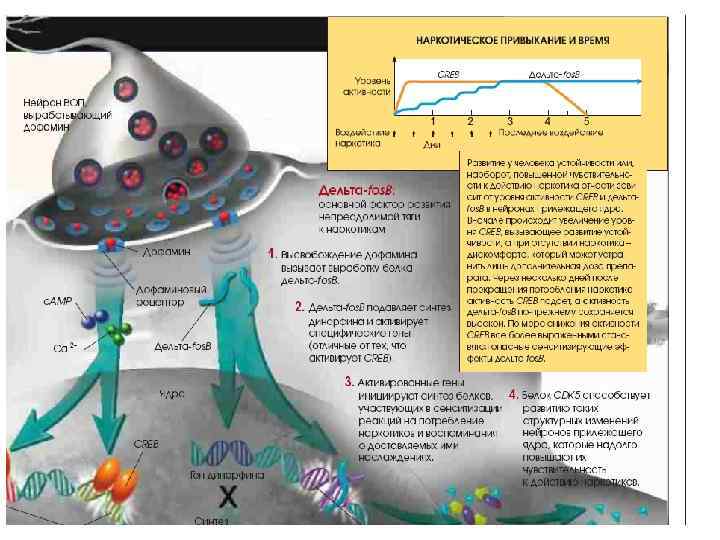

") (Fields, 2011)
(Fields, 2011)
 Сейчас считается, что эксплицитная память на нейрональном уровне представлена препостсинаптическим совпадением активности нейронов. Предполагается, что в приобретении и хранении новой информации участвует посттетаническая потенциация - длительное облегчение синаптической проводимости, обусловленное повторной стимуляцией возбуждающих синапсов. Большую роль в посттетанической потенциации играют глутаматные NMDA-рецепторы (N-метил-Dаспартат).
Сейчас считается, что эксплицитная память на нейрональном уровне представлена препостсинаптическим совпадением активности нейронов. Предполагается, что в приобретении и хранении новой информации участвует посттетаническая потенциация - длительное облегчение синаптической проводимости, обусловленное повторной стимуляцией возбуждающих синапсов. Большую роль в посттетанической потенциации играют глутаматные NMDA-рецепторы (N-метил-Dаспартат).

 Долговременная отенциация п сопровождается деполяризацией постсинаптической мембраны в результате активности HENMDA-рецепторных каналов. Деполяризация снимает блокаду NMDA-каналов магнием, что позволяет кальцию поступать в клетку через эти каналы. Кальций активирует зависимые от него киназы, и это приводит к индукции долговременной потенциации.
Долговременная отенциация п сопровождается деполяризацией постсинаптической мембраны в результате активности HENMDA-рецепторных каналов. Деполяризация снимает блокаду NMDA-каналов магнием, что позволяет кальцию поступать в клетку через эти каналы. Кальций активирует зависимые от него киназы, и это приводит к индукции долговременной потенциации.
 Роль генома в механизмах пластичности
Роль генома в механизмах пластичности
 Двухфазная регуляция транскрипции генов в клетке с помощью продуктов “ранних генов” Внеклеточные стимулы (гормоны, факторы роста, нейромедиаторы) могут вызвать активацию транскрипции “ранних генов", включая гены c-fos и c-jun. Воздействие экстраклеточных сигналов на промоторы этих генов осуществляется посредством вторичных мессенджеров. Fos, Jun и ряд других белков этого семейства синтезируются в цитоплазме и быстро транспортируются в ядро, где могут образовывать гетеромерные комплексы. Эти комплексы обладают специфической ДНК-связывающей активностью и способны изменять транскрипцию других генов-мишеней (“поздних” генов) (Curren & Morgan, 1991).
Двухфазная регуляция транскрипции генов в клетке с помощью продуктов “ранних генов” Внеклеточные стимулы (гормоны, факторы роста, нейромедиаторы) могут вызвать активацию транскрипции “ранних генов", включая гены c-fos и c-jun. Воздействие экстраклеточных сигналов на промоторы этих генов осуществляется посредством вторичных мессенджеров. Fos, Jun и ряд других белков этого семейства синтезируются в цитоплазме и быстро транспортируются в ядро, где могут образовывать гетеромерные комплексы. Эти комплексы обладают специфической ДНК-связывающей активностью и способны изменять транскрипцию других генов-мишеней (“поздних” генов) (Curren & Morgan, 1991).
 в мозге при обучении • В мозге взрослых") Особенности экспрессии непосредственных ранних генов (НРГ) в мозге при обучении • В мозге взрослых животных, находящихся в “спокойных” условиях транскрипция большинства НРГ находится на низком уровне. • В условиях, провоцирующих процесс обучения, – при потере результативности ранее выработанных действий животного, при новых и неожиданных воздействиях среды или исчезновении привычных и ожидаемых событий – происходит быстрая активация транскрипции НРГ в нервной системе. • Активация транскрипции НРГ начинается сразу после попадания животных в ситуацию обучения. • Экспрессия НРГ при обучении происходит в нервных, но не глиальных клетках. • Паттерны распределения клеток, экспрессирующих ранние гены, имеют генерализованный характер и охватывают обширные районы мозга. Конкретная топография экспрессии определяется характером воздействия и задачами обучения. • Экспрессия НРГ регулируется обучением в разные сроки постнатального развития, от рождения до взрослого возраста. • Активация НРГ затухает по мере потери новизны воздействия или после завершения выработки и автоматизации нового навыка. Обыденная поведенческая активность животных, выполнение ими приобретенных автоматизированных навыков, действие знакомых им сигналов и событий или нахождение животных в привычной им среде, не требующей обучения, не сопровождаются экспрессией НРГ в нервной системе.
Особенности экспрессии непосредственных ранних генов (НРГ) в мозге при обучении • В мозге взрослых животных, находящихся в “спокойных” условиях транскрипция большинства НРГ находится на низком уровне. • В условиях, провоцирующих процесс обучения, – при потере результативности ранее выработанных действий животного, при новых и неожиданных воздействиях среды или исчезновении привычных и ожидаемых событий – происходит быстрая активация транскрипции НРГ в нервной системе. • Активация транскрипции НРГ начинается сразу после попадания животных в ситуацию обучения. • Экспрессия НРГ при обучении происходит в нервных, но не глиальных клетках. • Паттерны распределения клеток, экспрессирующих ранние гены, имеют генерализованный характер и охватывают обширные районы мозга. Конкретная топография экспрессии определяется характером воздействия и задачами обучения. • Экспрессия НРГ регулируется обучением в разные сроки постнатального развития, от рождения до взрослого возраста. • Активация НРГ затухает по мере потери новизны воздействия или после завершения выработки и автоматизации нового навыка. Обыденная поведенческая активность животных, выполнение ими приобретенных автоматизированных навыков, действие знакомых им сигналов и событий или нахождение животных в привычной им среде, не требующей обучения, не сопровождаются экспрессией НРГ в нервной системе.
 Но как гены узнают, что им нужно включаться в нейронах того ансамбля, который участвовал в запоминании? Исследователи Уве Фрей и Ричард Моррис предложили гипотезу синаптических ярлыков: в ходе обучения в фазе кратковременного запоминания в синапсах обучающихся нейронов происходят небольшие метаболические изменения, в частности изменение некоторых мембранных белков. Эти измененные белки становятся своеобразными ярлыками активных синапсов, и именно к этим ярлыкам стремятся те «белки долговременного хранения» , которые были синтезированы с помощью поздних генов.
Но как гены узнают, что им нужно включаться в нейронах того ансамбля, который участвовал в запоминании? Исследователи Уве Фрей и Ричард Моррис предложили гипотезу синаптических ярлыков: в ходе обучения в фазе кратковременного запоминания в синапсах обучающихся нейронов происходят небольшие метаболические изменения, в частности изменение некоторых мембранных белков. Эти измененные белки становятся своеобразными ярлыками активных синапсов, и именно к этим ярлыкам стремятся те «белки долговременного хранения» , которые были синтезированы с помощью поздних генов.
 Как память может храниться десятками лет, если важнейшие элементы ее механизма — белки — живут в среднем двое суток? Гипотеза американского ученого Тодда Сактора - главной молекулой, ответственной за долговременное хранение, является фермент протеинкиназа — PKMzeta. Эта молекула увеличивает количество рецепторов в синапсе, т. е. его эффективность, А сигнал идет именно туда, где меньше всего сопротивления и где больше эффективность. При восприятии образа должен возникнуть ансамбль нейронов, где в каждом заработают эти молекулы. В каждом нейроне находятся матричные РНК, несущие информацию для синтеза этой протеинкиназы. Они заблокированы. Блокировка будет снята в тот момент, когда сойдется ряд стимулов, которые «приказывают» клетке надолго запомнить этот образ.
Как память может храниться десятками лет, если важнейшие элементы ее механизма — белки — живут в среднем двое суток? Гипотеза американского ученого Тодда Сактора - главной молекулой, ответственной за долговременное хранение, является фермент протеинкиназа — PKMzeta. Эта молекула увеличивает количество рецепторов в синапсе, т. е. его эффективность, А сигнал идет именно туда, где меньше всего сопротивления и где больше эффективность. При восприятии образа должен возникнуть ансамбль нейронов, где в каждом заработают эти молекулы. В каждом нейроне находятся матричные РНК, несущие информацию для синтеза этой протеинкиназы. Они заблокированы. Блокировка будет снята в тот момент, когда сойдется ряд стимулов, которые «приказывают» клетке надолго запомнить этот образ.
 Длительность хранения информации обеспечивает удивительное свойство этой молекулы. Протеинкиназа PKMzeta, однажды заработав, поддерживает сама себя: на смену одной приходит другая, в этом ей помогают другие белки, используя принцип положительной обратной связи.
Длительность хранения информации обеспечивает удивительное свойство этой молекулы. Протеинкиназа PKMzeta, однажды заработав, поддерживает сама себя: на смену одной приходит другая, в этом ей помогают другие белки, используя принцип положительной обратной связи.

 Из этих исследований последовали интересные следствия Следствие 1. Оказалось, что дефекты в некоторых из этих элементов каскада не только вызывают нарушение обучения у взрослых животных, но и являются причиной заболеваний, связанных с нарушениями умственного развития у детей. Такие заболевания, как синдром Рубинштейна–Тэйби, считались в течение долгого времени врожденными заболеваниями. Теперь поняли, что в действительности это нарушения, которые ведут к недостаткам возможности раннего обучения, формирования памяти у ребенка в первые недели и месяцы их жизни.
Из этих исследований последовали интересные следствия Следствие 1. Оказалось, что дефекты в некоторых из этих элементов каскада не только вызывают нарушение обучения у взрослых животных, но и являются причиной заболеваний, связанных с нарушениями умственного развития у детей. Такие заболевания, как синдром Рубинштейна–Тэйби, считались в течение долгого времени врожденными заболеваниями. Теперь поняли, что в действительности это нарушения, которые ведут к недостаткам возможности раннего обучения, формирования памяти у ребенка в первые недели и месяцы их жизни.
 Следствие 2 При расшифровке каскадов обнаружилось, что они сильно напоминают те клеточные процессы, которые происходят во время дифференцировки нервных клеток в развивающемся мозге. Они используют часто одни и те же сигнальные молекулы, причем, некоторые из этих молекул вначале были открыты при развитии, а потом, оказалось, как, например, различные нейротрофины, что они являются сигнальными молекулами и в моменты обучения. Это означает, что каждый эпизод развития напоминает эпизод обучения, или, что во взрослом мозге процессы развития никогда не заканчиваются. Каждый акт познания – это маленький эпизод морфогенеза и следующего развития. Но какой? - под когнитивным контролем, в отличие от того, что происходит во время эмбрионального развития. Иначе говоря, наши знания, наша психика, наш разум, определяя процессы приобретения новых знаний, являются также триггерами и для дифференцировки клеток, хранящих эти знания.
Следствие 2 При расшифровке каскадов обнаружилось, что они сильно напоминают те клеточные процессы, которые происходят во время дифференцировки нервных клеток в развивающемся мозге. Они используют часто одни и те же сигнальные молекулы, причем, некоторые из этих молекул вначале были открыты при развитии, а потом, оказалось, как, например, различные нейротрофины, что они являются сигнальными молекулами и в моменты обучения. Это означает, что каждый эпизод развития напоминает эпизод обучения, или, что во взрослом мозге процессы развития никогда не заканчиваются. Каждый акт познания – это маленький эпизод морфогенеза и следующего развития. Но какой? - под когнитивным контролем, в отличие от того, что происходит во время эмбрионального развития. Иначе говоря, наши знания, наша психика, наш разум, определяя процессы приобретения новых знаний, являются также триггерами и для дифференцировки клеток, хранящих эти знания.
.") Общность молекулярных механизмов регуляции экспрессии генов при развитии нервной системы и обучении (Анохин, 1996).
Общность молекулярных механизмов регуляции экспрессии генов при развитии нервной системы и обучении (Анохин, 1996).
 Таким образом, исследования молекулярной биологии памяти привели к следующим решениям: формирование долговременной памяти основано на активации универсального каскада ранних и поздних генов, ведущей к перестройке обучающегося нейрона, его молекулярного, белкового фенотипа. Т. обр. , в основе как врожденной, так и индивидуальной П лежат одни и те же процессы нейрогенеза, регулируемые одним и тем же каскадом экспрессии генов.
Таким образом, исследования молекулярной биологии памяти привели к следующим решениям: формирование долговременной памяти основано на активации универсального каскада ранних и поздних генов, ведущей к перестройке обучающегося нейрона, его молекулярного, белкового фенотипа. Т. обр. , в основе как врожденной, так и индивидуальной П лежат одни и те же процессы нейрогенеза, регулируемые одним и тем же каскадом экспрессии генов.
 Как много генов участвует в обучении? В оп-ах по скрещиванию и искусственному отбору плодовых мушек дрозофил (от лат. “любящие росу”, т. е покидающие свой кокон рано утром) по критерию “скорости выработки усл. р-ции избегания” (пробирки с определ-ым запахом). Нетривиален сам факт наличия мушек с разными спос-и к обучению! Основной рез-т – рост кривой эффект-ти обуч-ия прекращался только к 24 поколению непрерывно скрещиваемых особей (искусственный отбор). Это свидет-т о наличии нескольких генов, так или иначе (прямо или косвенно) определ-их спос-ть к обучению. Отбор же по спос-ти к сенситизации потребовал всего одного скрещивания: кривая эффект-ти сенсит. выходит в этом случае на плато уже через один цикл отбора особей, спос-ых к обуч-ию, и их скрещивания. Такая разница может служить своеобразной количой мерой различий в сложности мех-ов простой пластичности (сенсит. или привыкания) и мех-ов условно-рефл. актом ассоц-го повед-ия.
Как много генов участвует в обучении? В оп-ах по скрещиванию и искусственному отбору плодовых мушек дрозофил (от лат. “любящие росу”, т. е покидающие свой кокон рано утром) по критерию “скорости выработки усл. р-ции избегания” (пробирки с определ-ым запахом). Нетривиален сам факт наличия мушек с разными спос-и к обучению! Основной рез-т – рост кривой эффект-ти обуч-ия прекращался только к 24 поколению непрерывно скрещиваемых особей (искусственный отбор). Это свидет-т о наличии нескольких генов, так или иначе (прямо или косвенно) определ-их спос-ть к обучению. Отбор же по спос-ти к сенситизации потребовал всего одного скрещивания: кривая эффект-ти сенсит. выходит в этом случае на плато уже через один цикл отбора особей, спос-ых к обуч-ию, и их скрещивания. Такая разница может служить своеобразной количой мерой различий в сложности мех-ов простой пластичности (сенсит. или привыкания) и мех-ов условно-рефл. актом ассоц-го повед-ия.
") Роль метилирования промотора ДНК в управлении процессом транскрипции (эпигенетика)
Роль метилирования промотора ДНК в управлении процессом транскрипции (эпигенетика)


 Воспроизведение следа памяти – просто повторение или активный процесс перезаписи?
Воспроизведение следа памяти – просто повторение или активный процесс перезаписи?
 Опыты с цыплятами, клюющими бусинку с хинином 1. Цыплята, которые получали только инъекцию ингибитора синтеза белка, но не извлекали след памяти, - это одна контрольная группа - все прекрасно помнят (…). 2. Цыплята, которые получали только бусинку, но не получали блокаторы синтеза белка, также все прекрасно помнят. 3. Но, вот цыпленок вспоминает то, что было связано с этой бусинкой на фоне невозможности заново синтезировать белки. Это приводит к тому, что животное теряет память, которая, казалось, давно сформировалась. Иначе говоря, если вы вспомните что-то старое и в этот момент будет интерференция с процессом запоминания, - то это шанс вам забыть это старое.
Опыты с цыплятами, клюющими бусинку с хинином 1. Цыплята, которые получали только инъекцию ингибитора синтеза белка, но не извлекали след памяти, - это одна контрольная группа - все прекрасно помнят (…). 2. Цыплята, которые получали только бусинку, но не получали блокаторы синтеза белка, также все прекрасно помнят. 3. Но, вот цыпленок вспоминает то, что было связано с этой бусинкой на фоне невозможности заново синтезировать белки. Это приводит к тому, что животное теряет память, которая, казалось, давно сформировалась. Иначе говоря, если вы вспомните что-то старое и в этот момент будет интерференция с процессом запоминания, - то это шанс вам забыть это старое.
 «…описание воспоминаний как фиксированных и безжизненных, есть всего лишь ошибочная фантазия; воспоминание не является повторным возбуждением неисчислимых фиксированных фрагментов следов, оно - всегда творческое воссоздание или конструирование, складывающееся из нашего отношения ко всей активной массе реакции и опыта прошлого» (Ф. Бартлетт)
«…описание воспоминаний как фиксированных и безжизненных, есть всего лишь ошибочная фантазия; воспоминание не является повторным возбуждением неисчислимых фиксированных фрагментов следов, оно - всегда творческое воссоздание или конструирование, складывающееся из нашего отношения ко всей активной массе реакции и опыта прошлого» (Ф. Бартлетт)
: «Как ты знаешь, я сейчас работаю над") З. Фрейд (из письма своему другу Флису): «Как ты знаешь, я сейчас работаю над новой теорией памяти, согласно которой, память – это ни что-то раз данное и навеки закрепленное, а меняющееся при каждом извлечении. Процесс, который я называю ретранскрипцией памяти» .
З. Фрейд (из письма своему другу Флису): «Как ты знаешь, я сейчас работаю над новой теорией памяти, согласно которой, память – это ни что-то раз данное и навеки закрепленное, а меняющееся при каждом извлечении. Процесс, который я называю ретранскрипцией памяти» .
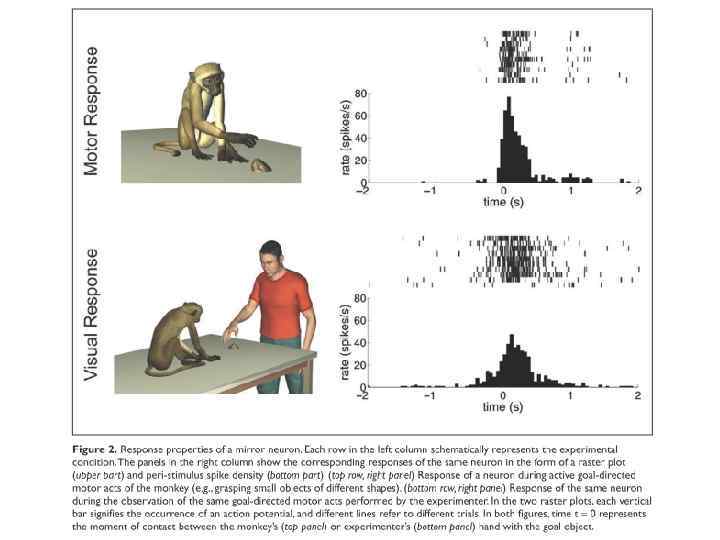
, профессор Пармского университета, Италия") Зеркальные нейроны Джакомо Риззолатти (Rizzolatti), профессор Пармского университета, Италия
Зеркальные нейроны Джакомо Риззолатти (Rizzolatti), профессор Пармского университета, Италия
") ЗН в мозге человека (п. п. 44 -45)
ЗН в мозге человека (п. п. 44 -45)
 Свойства ЗН: связь реакций с пространственной локализацией действия
Свойства ЗН: связь реакций с пространственной локализацией действия
") Свойства ЗН: связь с целью действий (намерениями)
Свойства ЗН: связь с целью действий (намерениями)
 Дефекты «зеркал» : аутизм Но что будет, если зеркальные нейроны перестанут работать или будут работать неправильно? В 2000 году известный нейрофизиолог из Калифорнийского ун-та в Сан-Диего Вилаянур Рамачандран (Vilayanur S. Ramachandran; дир-р Center for Brain and Cognition Калифорнийского университета в Сан-Диего; Рамачандран занимает пост директора центра, автор многочисленных работ, посвященных феномену фантомных конечностей и синестезии) предположил, что именно нарушение работы зеркальных нейронов является причиной детского аутизма — нарушения, от которого страдают, в среднем, два ребёнка на тысячу детей. Аутичные дети стараются отгородиться от внешнего мира и не устанавливать никаких отношений с другими людьми. Хотя их речь сохранна, они практически всегда молчат, но, в отличие от немых, они не пытаются объясняться при помощи мимики и жестов. Также аутичные дети не могут воспроизвести действия других людей. Им не чужды чувства страха, гнева и удовольствия, но они глухи к переживаниям других людей и не замечают тонких оттенков их поведения, понятных большинству их сверстников.
Дефекты «зеркал» : аутизм Но что будет, если зеркальные нейроны перестанут работать или будут работать неправильно? В 2000 году известный нейрофизиолог из Калифорнийского ун-та в Сан-Диего Вилаянур Рамачандран (Vilayanur S. Ramachandran; дир-р Center for Brain and Cognition Калифорнийского университета в Сан-Диего; Рамачандран занимает пост директора центра, автор многочисленных работ, посвященных феномену фантомных конечностей и синестезии) предположил, что именно нарушение работы зеркальных нейронов является причиной детского аутизма — нарушения, от которого страдают, в среднем, два ребёнка на тысячу детей. Аутичные дети стараются отгородиться от внешнего мира и не устанавливать никаких отношений с другими людьми. Хотя их речь сохранна, они практически всегда молчат, но, в отличие от немых, они не пытаются объясняться при помощи мимики и жестов. Также аутичные дети не могут воспроизвести действия других людей. Им не чужды чувства страха, гнева и удовольствия, но они глухи к переживаниям других людей и не замечают тонких оттенков их поведения, понятных большинству их сверстников.
 Для того, чтобы продемонстрировать нарушение функций зеркальных нейронов у детей с аутизмом, Рамачандран регистрировал мю-ритм (8– 13 Гц; максимум в сенсомоторной коре). Этот ритм исчезает, когда человек совершает любое произвольное действие, а также тогда, когда испытуемый наблюдает, как то же самое действие совершает другой человек. Рамачандран предположил, что реакцию подавления мю-волн можно использовать в качестве простого, надежного и безопасного инструмента для изучения активности зеркальных нейронов. Оказалось, что у аутичных детей, как и у здоровых, мю-ритм блокируется, когда они совершают произвольные движения. Однако в том случае, когда аутичный ребёнок наблюдал за выполнением того же движения другим человеком, подавления мю-волн не происходило. На основании этого Рамачандран заключил, что система моторных командных нейронов у аутичного ребенка оставалась целой и невредимой, но функции его системы зеркальных нейронов были нарушены.
Для того, чтобы продемонстрировать нарушение функций зеркальных нейронов у детей с аутизмом, Рамачандран регистрировал мю-ритм (8– 13 Гц; максимум в сенсомоторной коре). Этот ритм исчезает, когда человек совершает любое произвольное действие, а также тогда, когда испытуемый наблюдает, как то же самое действие совершает другой человек. Рамачандран предположил, что реакцию подавления мю-волн можно использовать в качестве простого, надежного и безопасного инструмента для изучения активности зеркальных нейронов. Оказалось, что у аутичных детей, как и у здоровых, мю-ритм блокируется, когда они совершают произвольные движения. Однако в том случае, когда аутичный ребёнок наблюдал за выполнением того же движения другим человеком, подавления мю-волн не происходило. На основании этого Рамачандран заключил, что система моторных командных нейронов у аутичного ребенка оставалась целой и невредимой, но функции его системы зеркальных нейронов были нарушены.
 Эксперименты Рамачандрана: депрессия мюритма при совершении реального движения
Эксперименты Рамачандрана: депрессия мюритма при совершении реального движения
 Эксперименты Рамачандрана: депрессия мю-ритма в норме и отсутствие таковой у аутистов при наблюдении за выполнением движения
Эксперименты Рамачандрана: депрессия мю-ритма в норме и отсутствие таковой у аутистов при наблюдении за выполнением движения
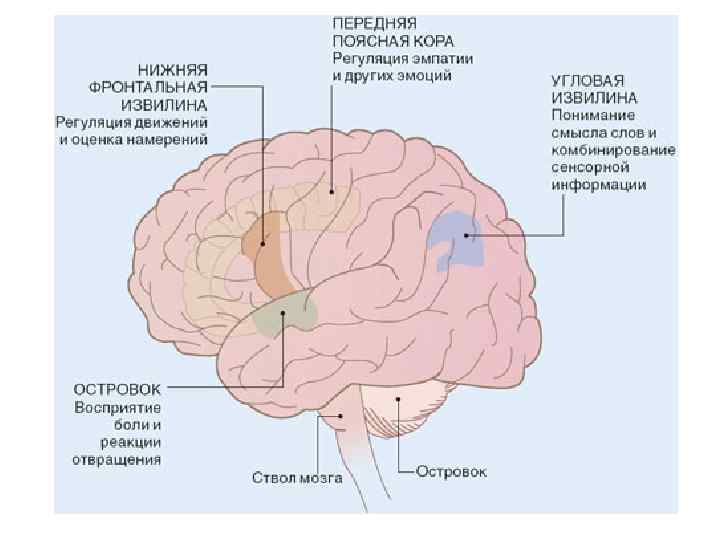
 Анатомия аутизма У пациентов с аутизмом отмечается снижение активности зеркальных нейронов в нижней фронтальной извилине — одном из отделов премоторной коры мозга. Данным обстоятельством можно объяснить их неумение распознавать намерения других людей. Дисфункции зеркальных нейронов островковой и передней поясной коры могут обусловливать их неспособность к сопереживанию, а нарушения зеркальной системы угловой извилины — дефекты речи. У людей с аутизмом выявлены и структурные изменения в мозжечке и стволе мозга.
Анатомия аутизма У пациентов с аутизмом отмечается снижение активности зеркальных нейронов в нижней фронтальной извилине — одном из отделов премоторной коры мозга. Данным обстоятельством можно объяснить их неумение распознавать намерения других людей. Дисфункции зеркальных нейронов островковой и передней поясной коры могут обусловливать их неспособность к сопереживанию, а нарушения зеркальной системы угловой извилины — дефекты речи. У людей с аутизмом выявлены и структурные изменения в мозжечке и стволе мозга.
 Исследования ЗН открывают новые возможности в лечении аутизма Если причина аутизма заключается в нарушении функции зеркальной системы мозга, можно попытаться разработать новые подходы к диагностике и лечению этого расстройства. 1) Так, для раннего выявления данного заболевания у детей врачи могут использовать в качестве показателя отсутствие подавления мю-ритма в определенных ситуациях. Чем раньше аутизм будет диагностирован, тем быстрее можно начать его поведенческую терапию. Временной фактор имеет здесь критическое значение: лечение аутизма подобными методами утрачивает эффективность после появления основных симптомов расстройства (обычно в возрасте от двух до четырех лет).
Исследования ЗН открывают новые возможности в лечении аутизма Если причина аутизма заключается в нарушении функции зеркальной системы мозга, можно попытаться разработать новые подходы к диагностике и лечению этого расстройства. 1) Так, для раннего выявления данного заболевания у детей врачи могут использовать в качестве показателя отсутствие подавления мю-ритма в определенных ситуациях. Чем раньше аутизм будет диагностирован, тем быстрее можно начать его поведенческую терапию. Временной фактор имеет здесь критическое значение: лечение аутизма подобными методами утрачивает эффективность после появления основных симптомов расстройства (обычно в возрасте от двух до четырех лет).
 Многообещающим представляется также использование биологической обратной связи. Врачи (психологи) могут регистрировать мю-волны у") 2) Многообещающим представляется также использование биологической обратной связи. Врачи (психологи) могут регистрировать мю-волны у ребенка с аутизмом и демонстрировать их на экране. Если функции зеркальных нейронов утрачены не окончательно, то дети могут научиться (методом проб и ошибок, используя зрительную обратную связь) подавлять мю-ритм, глядя на монитор. 3) Еще один возможный подход основан на коррекции биохимического равновесия нейромодуляторов, нарушение которого, возможно, приводит к нарушению функций ЗН у аутистов. В подтверждение тому факт влияния метилендиоксиметамфетамина (МДМА) (экстази), способствующего повышению эмоциональной близости и общительности людей. Не исключено, что удастся модифицировать данное соединение и создать на его основе безопасное и эффективное средство для облегчения хотя бы некоторых симптомов аутизма.
2) Многообещающим представляется также использование биологической обратной связи. Врачи (психологи) могут регистрировать мю-волны у ребенка с аутизмом и демонстрировать их на экране. Если функции зеркальных нейронов утрачены не окончательно, то дети могут научиться (методом проб и ошибок, используя зрительную обратную связь) подавлять мю-ритм, глядя на монитор. 3) Еще один возможный подход основан на коррекции биохимического равновесия нейромодуляторов, нарушение которого, возможно, приводит к нарушению функций ЗН у аутистов. В подтверждение тому факт влияния метилендиоксиметамфетамина (МДМА) (экстази), способствующего повышению эмоциональной близости и общительности людей. Не исключено, что удастся модифицировать данное соединение и создать на его основе безопасное и эффективное средство для облегчения хотя бы некоторых симптомов аутизма.
 Теория «эмоционального ландшафта» Гипотеза дисфункций зеркальных нейронов не в состоянии объяснить происхождение всех симптомов аутизма, таких, например, как повторяющиеся движения (раскачивание из стороны в сторону), избегание визуального контакта с собеседником, гиперчувствительность и отвращение к определенным звукам. Для выяснения их природы Рамачандран и др. разработали так называемую теорию эмоционального ландшафта (salience landscape theory). Суть теории. Миндалина формирует «эмоциональный ландшафт» внешней среды, своего рода карту значимости всех объектов и событий, окружающих человека. У детей с аутизмом эмоциональный ландшафт окружения искажен вследствие нарушения связей между корковыми областями, перерабатывающими сенсорную информацию, и миндалиной, или между лимбическими структурами мозга и фронтальными долями коры, которые регулируют возникающее в результате поведение. Нарушение связей приводит к тому, что любой пустяк может вызвать у ребенка экстремальную эмоциональную реакцию — настоящую «автономную бурю» . Данная гипотеза объясняет стремление детей с аутизмом избегать зрительных контактов и вообще всех непривычных ощущений, способных привести их в смятение. Искаженным восприятием эмоциональной значимости окружающих предметов и событий можно объяснить и тот факт, почему многие аутичные дети нередко поглощены пустяками и совершенно равнодушны к тому, что вызывает острый интерес у их здоровых сверстников.
Теория «эмоционального ландшафта» Гипотеза дисфункций зеркальных нейронов не в состоянии объяснить происхождение всех симптомов аутизма, таких, например, как повторяющиеся движения (раскачивание из стороны в сторону), избегание визуального контакта с собеседником, гиперчувствительность и отвращение к определенным звукам. Для выяснения их природы Рамачандран и др. разработали так называемую теорию эмоционального ландшафта (salience landscape theory). Суть теории. Миндалина формирует «эмоциональный ландшафт» внешней среды, своего рода карту значимости всех объектов и событий, окружающих человека. У детей с аутизмом эмоциональный ландшафт окружения искажен вследствие нарушения связей между корковыми областями, перерабатывающими сенсорную информацию, и миндалиной, или между лимбическими структурами мозга и фронтальными долями коры, которые регулируют возникающее в результате поведение. Нарушение связей приводит к тому, что любой пустяк может вызвать у ребенка экстремальную эмоциональную реакцию — настоящую «автономную бурю» . Данная гипотеза объясняет стремление детей с аутизмом избегать зрительных контактов и вообще всех непривычных ощущений, способных привести их в смятение. Искаженным восприятием эмоциональной значимости окружающих предметов и событий можно объяснить и тот факт, почему многие аутичные дети нередко поглощены пустяками и совершенно равнодушны к тому, что вызывает острый интерес у их здоровых сверстников.