incompat.ppt
- Количество слайдов: 76



ВЕГЕТАТИВНАЯ НЕСОВМЕСТИМОСТЬ ГРИБОВ – ПРОСТЕЙШИЙ МЕХАНИЗМ ИММУННОГО ОТВЕТА

Важнейшие процессы, сопровождающие рост и развитие колоний мицелиальных грибов, удлинение кончиков гиф, ветвление и слияние гиф (анастомозы)
 клетками. Апикальные клетки сумчатых грибов варьируют по длине")
Все мицелиальные грибы растут верхушечными (апикальными) клетками. Апикальные клетки сумчатых грибов варьируют по длине и числу ядер, в то время как остальные клетки имеют одинаковые размеры и 3 -4 ядра. Ядра в апикальной клетке находятся в состоянии активного деления с синхронизированным митотическим циклом, при котором во всех ядрах клетки отдельные этапы деления протекают одновременно. В остальных клетках митозы не происходят (интерфазные ядра) до тех пор, пока не произойдет ветвление, и клетка станет апикальной. И тогда в ней начинается активная деятельность.

Из внутренних частей клетки к периферии направляются пузырьки, содержащие углеводы – заготовки для строительства новой клеточной стенки. Их содержимое выплескивается на поверхность клетки, где встроенные в оболочку ферменты собирают из них полисахариды – хитин и глюканы, из которых построена клеточная стенка. Скорость удлинения кончика гифы Neurospora crassa составляет 16 мкм в минуту, что требует потока микровезикул со строительным материалом, интенсивностью 37000 везикул в минуту и их слияния с мембраной

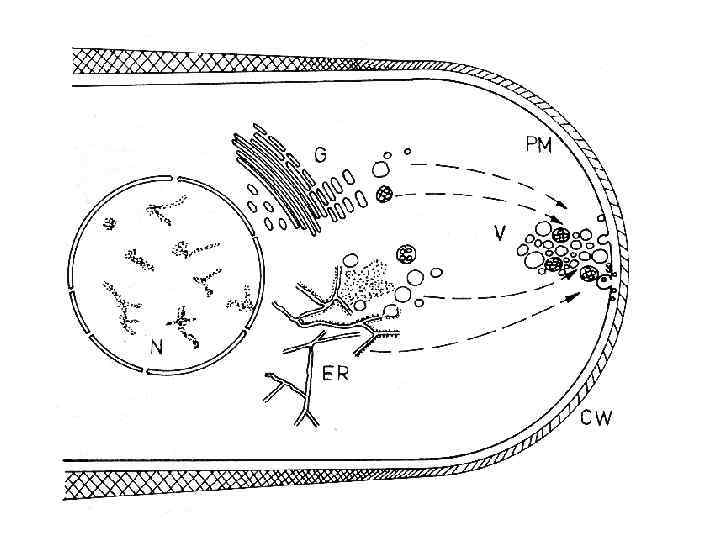

Движение пузырьков со строительными материалами осуществляется по внутриклеточным железнодорожным путям, называемым микротрубочками и микрофиламентами. Вблизи верхушки клетки рельсы заканчиваются, и пузырьки поступают на железнодорожный узел называемый spitzenkorper или верхушечное тело. Здесь происходит распределение отдельных пузырьков по необходимым участкам растущего кончика гифы. Достигнув мембраны клетки, пузырек сливается с ней и выливает содержимое на поверхность. А встроенные в клеточную стенку ферменты соединяют прибывшие строительные материалы по правилам, записанным в генетическом коде клетки
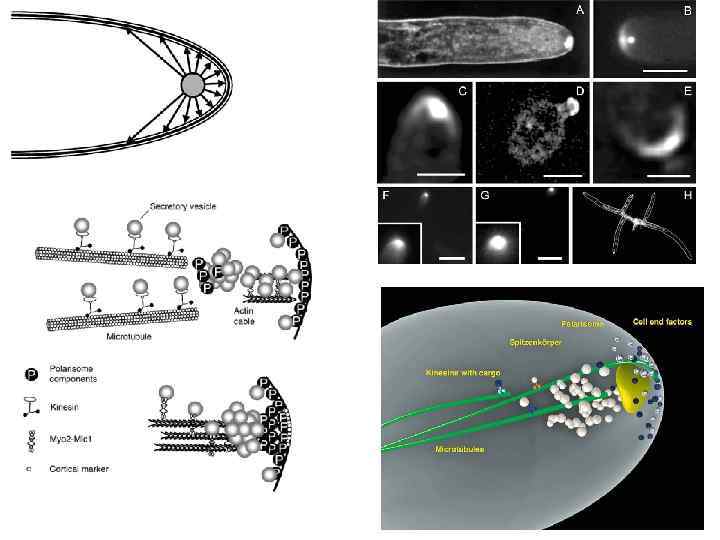

Для обеспечения процессов синтеза необходимых высокомолекулярных соединений и транспорта их к периферии клетки необходимо иметь много различных химических веществ и запас энергии. Однако электрофизиологические эксперименты (погружение микроэлектродов в мембраны клеток нейроспоры) показали, что в верхушечных клетках ее мицелия практически отсутствует мембранный потенциал, то есть верхушечная клетка, в отличие от ее расположенных ближе к центру соседок, не способна всасывать химические соединения из окружающей среды.

Изолированная от мицелия верхушечная клетка не может расти и делиться. Рост начинается только при наличии фрагмента из трех-четырех клеток. Клетки, примыкающие к верхушечной, имеют активный мембранный потенциал, более высокий, чем у клеток других исследованных организмов, который обеспечивает создание протонного насоса, необходимого для активного поступления метаболитов из окружающей среды в клетку, а также для мощного тока веществ в направлении верхушечной клетки. Вот для чего нужны отверстия в перегородках.

Итак. Вещества для построения клеточной стенки и других элементов клетки притекают в верхушечную клетку из соседних через поры в перегородках, а откуда берется необходимая энергия? Оказывается, грибы нашли простой выход. Плазматическая мембрана внутренних клеток мицелия для создания протонного насоса буквально нашпигована ферментами, освобождающими энергию, запасенную в аденозинтрифосфате (АТФ). На площади мембраны, равной 1 квадратному сантиметру, расположено несколько сотен миллиардов молекул этих ферментов, и каждая молекула ежесекундно выбрасывает 200 -300 протонов, создавая направленный наружу электрический ток.

Протонный насос нейроспоры потребляет около трети всей энергии, производимой клеткой. Это – плата за возможность активного всасывания веществ из окружающий среды и транспорта их по мицелию. Так вот, как сказано выше, верхушечная клетка лишено протонного насоса (зачем тратится на активное всасывании веществ из среды роста, если они поступают из внутренних клеток), поэтому вся энергия клетки идет на вытягивание и деление материнской клетки на два дочерние.

Получается разделение функций: одни клетки питаются¸ другие – растут и делятся. При разделении верхушечной клетки на две, только нижняя приобретает мембранный потенциал, необходимый для создания протонного насоса, причем, чем дальше от периферии к центру расположена клетка, тем ее потенциал выше. Так что мицелий – это единый организм, участки которого имеют различные функции.

Ветвление гиф происходит следующим образом: в отдельных клетках активизируется синтез гидролитических ферментов – хитиназ и глюканаз, локально разрушаются основные полисахариды стенки. Поврежденный участок клеточной стенки размягчается и под действием внутриклеточного тургорного давления выпячивается. Материнская клетка превращается в апикальную, в ней начинаются митозы и синтез полуфабрикатов – заготовок для дальнейшего вытягивания и укрепления клеточной стенки выроста. После перехода в него разделившихся ядер, вырост отделяется оболочкой от материнской клетки, которая успокаивается, а отросток от нее приобретает свойства апикальной клетки.
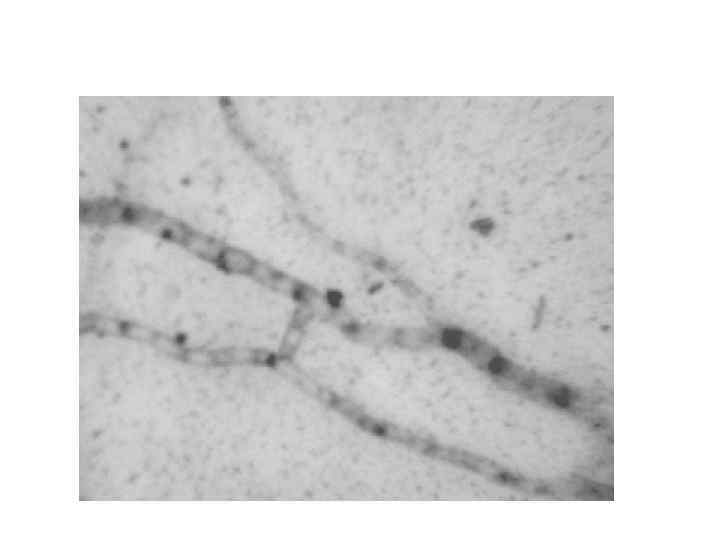

Анастомозы, происходящие между соседними гифами внутри колонии, армируют ее делают более прочной, сохраняют ее единство при распространении в пространстве

При слиянии гиф двух или более контактирующих колоний происходит 1/ Объединение в одной клетке нескольких генетически различных ядер, что обеспечивает состояние гетерокариоза, который у гаплоидных грибов заменяет диплоидность и является начальной стадией парасексуальной рекомбинации.

«Гетерокариоз – гибкий механизм физиологической адаптации, суть которого заключается в количественных изменениях качественно фиксированного множественного генома» R. Stanier, 1956


2/. Гетерокариоз – основа парасексуального процесса, т. е. рекомбинации без полового процесса.

3/ Анастомозы между гифами с последующий взаимной миграцией ядер приводят к исчезновению внутрипопуляционных единиц – индивидуумов, клонов и интеграции их в единую сеть мицелия (Buller, 1933).

Однако исследование многих изолированных из природных субстратов штаммов грибов, относящихся к различным таксонам, показало, что большинство изолятов при попарном посеве вегетативно несовместимы.

Поскольку вегетативная несовместимость препятствует формированию гетерокарионов со всеми их адаптивными преимуществами, наличие сильного естественного отбора в пользу несовместимости заставило обратить пристальное внимание, как на сам феномен, так и на его эволюционные последствия.

1. ЦИТОЛОГИЯ ВН Наиболее острое проявление ВН – барраж, при котором слившиеся и прилегающие к ним клетки погибают. Цитоплазма становится гранулированной, тонко-вакуолизированной, размеры вакуолей постепенно увеличиваются, межвакуолярное пространство уплотняется, плазмалемма отделяется от клеточной стенки. В зоне контакта двух колоний возникает более или менее четко заметная линия из мертвых клеток, иногда отличающаяся от прилегающих участков колонии пигментацией. Поры в септах закрываются пробками, по-видимому, для локализации погибающих участков гиф.
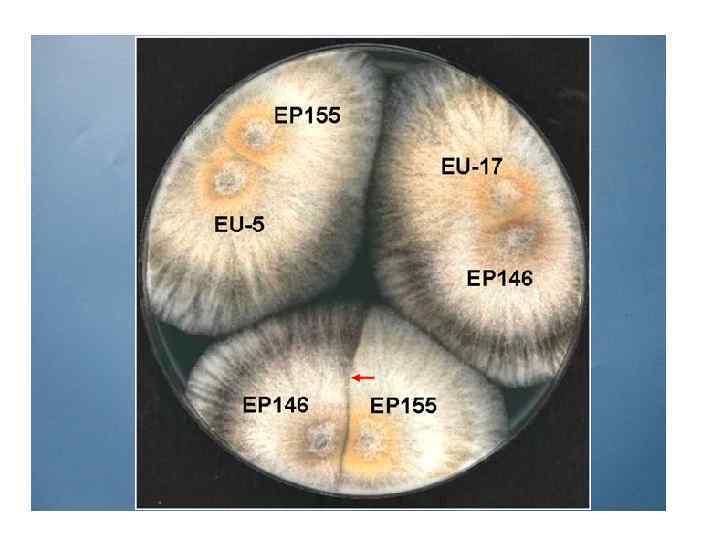
")
Взаимодействие колоний штаммов C. parasitica (фото Н. Н. Гринько)
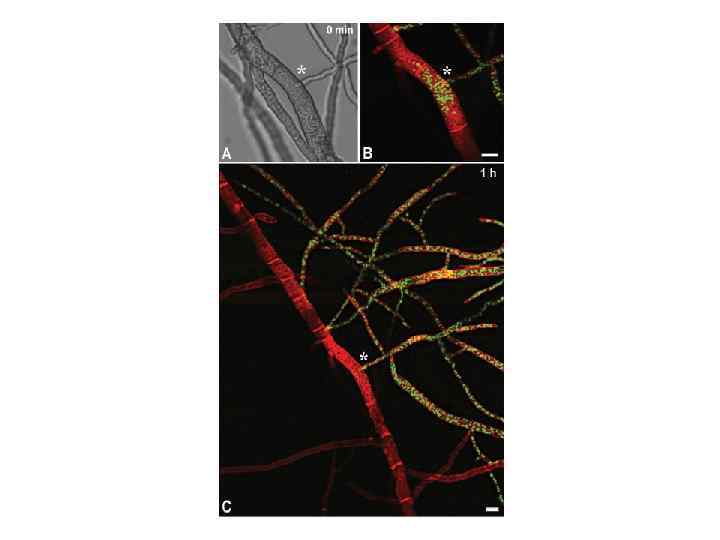



2. ГЕНЕТИКА ВН Система het- или vic-генов. Гетероаллельность по любому из этих генов вызывает острую реакцию гибели слившихся и прилегающих к ним клеток (барраж). Эта система генетического контроля несовместимости наиболее полно исследована у двух аскомицетов – N. crassa и Podospora anserina. У N. crassa несовместимость вызывает, в большинстве случаев, взаимодействие продуктов аллельных генов, а у P. anserina наряду с аллельной распространена и неалелльная несовместимость, при которой несовместимы штаммы, имеющие определенные аллели в двух несцепленных локусах. Последняя система усложнена наличием многих аллелей в нескольких mod-локусах, подавляющих проявление неаллельной несовместимости.

Взаимодействия между продуктами аллельных и неаллельных локусов Podospora anserina

ПОЛИМОРФИЗМ HET-ГЕНОВ У Neurospora crassa – 11 het-локусов; У Aspergillus nidulans – 18; У Cryphonectria parasitica – 6; У Podospora anserina – 9 Het и Pds гены аскомицетов из разных классов (дрожжей, эвроциевых и сордариевых) гомологичны.

3. ПРОДУКТЫ het-ГЕНОВ Локус het-s Podospora anserina включает аллели het-s het-S и нейтральный het-sc (совместимый с het-s и het. S). Они кодируют белки, имеющие высокую гомологию (различаются по 13 аминокислотам из 289). После анастомозов возможна конверсия белка нейтрального штамма het-sc (совместимого с het-s и het-S) в белок HET-s, т. е. этот белок имеет свойства прионного белка (Silar, Daboussi, 1999).
,")
Система неаллельных генов het-c/het-e и hetc/het-d. Ген het-c кодирует синтез небольшого белка (208 аминокислот), имеющего гомологию с ферментом гликолипид трансферазой. Делеция het-c не оказывает влияния на вегетативный рост, но приводит к почти полной редукции образования аскоспор. Получены мутации, вызывающие ненормальное ветвление апикальных гиф. Предполагают, что НЕТ-с белок влияет на биосинтез клеточной стенки (Saupe et al. , 1994).

Гены het-e и het-d контролируют крупные паралогичные белки, имеющие три функциональных домена: мотив связывания ГТФ (N-терминальный НЕТ-домен), центральный NACHT-домен и С-терминальные повторы последовательности WD 40, обеспечивающей взаимодействие белок-белок (Espagne et al. , 1997; Smith et al. , 2000).

Молекулярный механизм узнавания чужого – прямое взаимодействие белков – продуктов het-генов (благодаря наличию домена WD-40), причем димеризуются только гетеромерные белки
")
4. Вегетативная несовместимость – форма програмированной смерти клетки (PCD)

Отмирание клеток дрожжей и мицелиальных грибов сопряжено с процессами, характерными для PСD высших эукариот (фрагментация ДНК, сохранение целостности мембран, аккумуляция активных форм кислорода, зависимость от белкового синтеза, наличие метакаспаз, подавление ингибиторами каспаз и др. )

Отмирание клеток при ВН протекает по типу II PCD, для которого характерна автофагия. При этом цитоплазматический материал разбивается в везикулы, покрытые двумя мембранами – автофагосомы. Их наружные мембраны сливаются с мембранами вакуолей и лизосом, превращаясь внутри них в автофаговые тельца, покрытые одной мембраной, и перевариваются. Признаки ВН, характерные для PCD: повышенная протеолитическая активность, вакуолизация клеток, положительная реакция ядер на TUNEL-тест и др.

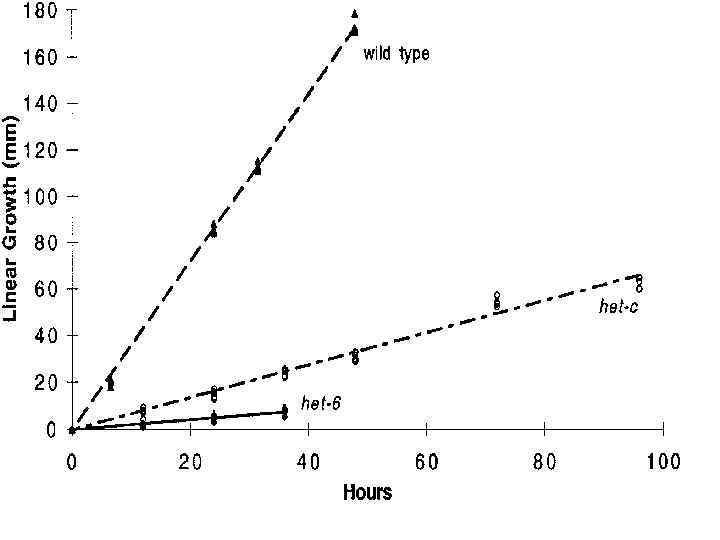
Штаммы Neurospora crassa, гетероаллельные по локусам het-c и het-6

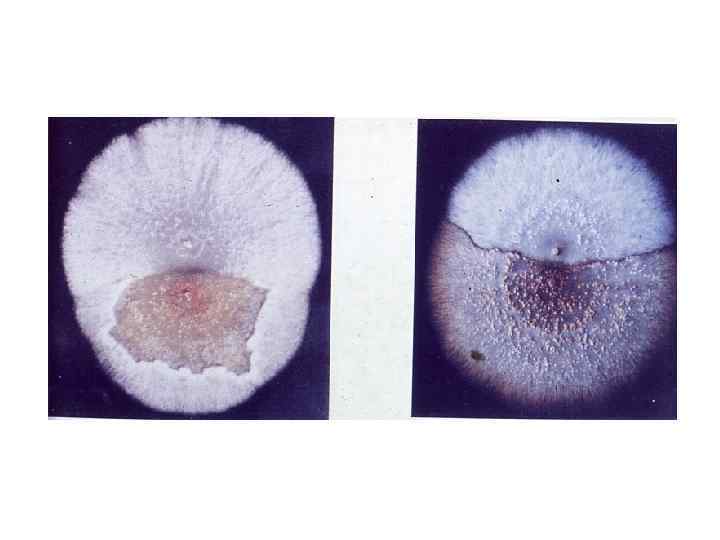
Гетерокарионы между мутантами несовместимых штаммов Phytophthora infestans

Неаллельная несовместимость P. anserina, обусловленная локусами R-V, температурочувствительна и не проявляется у штаммов, растущих при рестриктивных температурах. Сконструированы штаммы P. anserina, содержащие в геноме оба несовместимых локуса. Они способны нормально расти при повышенной температуре, но погибали при пониженной
 и участвующих")
Несовместимость R/V штаммов сопровождается экспрессией нескольких генов, названных idi (induced during incompatibility) и участвующих в регуляции процессов вакуолизации. Гены idi-1, idi-2, idi 3 кодируют низкомолекулярные белки, локализованные в клеточной стенке.

IDI-1 - септальный белок, а септирование мицелия усиливается при голодании, ВН и после обработки ингибитором TOR-киназы рапамицином. Поэтому индукция экспрессии idi-1 – idi-3 генов является защитной реакцией на ВН, направленной на локализацию повреждения и построение барьеров на пути его распространения вдоль мицелия. idi-4 – фактор регуляции транскрипции; idi-6 кодирует вакуолярную сериновую протеазу, участвующую в деградации автофаговых телец;
 – ортолог генов автофагии дрожжей (ATG 8) и человека")
idi-7 (или Pa. ATG 8) – ортолог генов автофагии дрожжей (ATG 8) и человека (LC-3). У мутантов, имеющих инактивированный ген Ра. ATG 8, не формировались автофагосомы, т. е был нарушен процесс автофагии, но реакция несовместимости наступала раньше, чем у немутантных штаммов, и протекала более интенсивно. Следовательно, автофагия сопровождает ВН, но не является ее причиной. Полагают, что автофагия защищает клетки мицелиев, прилегающие к анастомозированным, от распространения сигнала PCD.

5. Полиморфизм и эволюция het-локусов Многие het-локусы полиморфны У Podospora anserina: НЕТ-6 – 35; НЕТ-D – 94; PIN – 51; TOL – 48 гомологов het-локусы подвержены действию дарвиновского балансового отбора, характерными чертами которого являются:

1/. Межвидовой полиморфизм. Аллельная специфичность локуса het-c N. crassa обусловлена вариабельностью домена из 34 -48 аминокислот в белке НЕТ-С. ВН происходит при взаимодействии штаммов, гетероаллельных по любым двум из трех аллельных состояний локуса (het-c. OR , het-c. PA и hetc. GR). Полиморфные варианты всех трех аллелей обнаружены в природных штаммах грибов из семейства Sordariaces (Neurospora spp. , Sordaria spp. , Gelasinospora sрp. ), т. е. дивергенция полиморфизмов происходит по ортологическим группам, а не по таксонам.

2/. Присутствие большого числа аллельных вариантов в популяциях. У P. anserina неаллельная несовместимость обусловлена взаимодействиями продуктов определенных аллелей локуса het-C и локусов het-e и het-d. Эти продукты высоко вариабельны. В генах het-e и het-d наиболее полиморфен домен, контролирующий WD-повторы, которые обеспечивают белковые взаимодействия и необходимы для формирования гетеромерных белков. 4 кодона этого домена находятся под давлением сильного позитивного отбора. Вследствие механизма согласованной эволюции изменения в одном повторе затрагивают изменения в остальных. Поскольку анализируемые het-гены содержат не менее 10 повторов, можно предположить, сколь высок потенциал изменчивости.

3/. Примерно равное соотношение аллельных частоты в популяциях. Это правило соблюдается для большинства hetгенов в популяциях N. crassa и P. anserina, однако в по-пуляциях Cryphonectria parasitica наблюдалось отклонение в соотношениях аллельных частот от 1: 1

4/. Частота несинонимических замен на сайт в полиморфных районах локусов превышает частоту синонимических замен. При стабилизирующем отборе, направленном на сохранение фенотипа, отбор отметает несинонимические мутации, вследствие чего отношение их частоты к частоте синонимических мутаций бывает меньше единицы. В локусах, полиморфизм которых благотворен для популяции, наоборот, такое соотношение превышает единицу. Превышение частоты несинонимических замен над синонимическими установлено для вариабельных доменов локуса het-c в популяциях N. crassa и локусов het-s, het-c, het-e, het-d в популяциях P. anserina

6. ВН – ФОРМА ПРОСТЕЙШЕГО ИММУННОГО ОТВЕТА ВН грибов имеет все признаки, позволяющие рассматривать ее как иммунную реакцию.

1/. ВН есть форма узнавания и отторжения «чужого» , ибо чем больше у контактирующих штаммов гетероаллельных het-локусов, и, следовательно, чем больше различия их геномов, тем сильнее реакция отторжения. Отторжение чужого – одна из важнейших иммунных функций организмов (напр. реакции гистонесовместимости).

2/. ВН сопровождается PCD, которая является наиболее распространенной формой иммунного ответа у растений и животных.

3/. Внутрипопуляционная вариабельность частот аллелей het-генов подвержена позитивному дарвиновскому отбору, причем число несинонимических замен оснований ДНК в вариабельных участках генов превышает число синонимических замен, что характерно для большинства генов иммунного ответа у растений, комплекса генов гистосовместимости млекопитающих, поверхностных антигенов паразитов и вирусов.

7. ЭКОЛОГИЧЕСКАЯ РОЛЬ ВН
.")
1. Ограничение гетероплазмона (защита от вирусной инфекции).

В мицелии возбудителя рака каштанов Сryphonectris parasitica описано семейство вирусов, которые вызывают ряд негативных воздействий, в т. ч. гиповирулентность. Их передача через анастомоз приводит к превращению вирулентного штамма в гиповирулентный

Разные комбинации het-генов тормозят перенос в разной степени. Показано, что 1/ интенсивность переноса вируса из одного штамма в другой отрицательно коррелирует с интенсивностью PCD, 2/ при асимметричной гибели клеток анастомозирующих штаммов перенос вируса идет слабее в тот штамм, у которого погибает больше клеток, 3/ в одних случаях вирусная инфекция, по-видимому, может супрессировать развитие PCD, а в других – зараженные клетки, при слиянии, наоборот, интенсивнее погибают, чем не зараженные

Таким образом, у мицелиальных грибов PCD при ВН имеет такой же характер защиты от инфекции, как у растений и животных.

1. Защита занятой территории, обозначение границ индивидуума. При непрерывной сети Буллера, исчезает понятие о фенотипической или генетической пространственно-дискретной индивидуальности. Индивидуальность генетически разнородных споровых проростков кратковременна, ибо они рано или поздно включались бы в единую мицелиальную цепь, растворяя свою индивидуальность в общем гетерокариотическом котле. Поэтому непрерывный мицелий Буллера – эволюционный тупик, защита от которого – ВН.



8. РОЛЬ ВН В ЭВОЛЮЦИИ ПОПУЛЯЦИЙ И В ВИДООБРАЗОВАНИИ Видообразование грибов без географической изоляции может быть вызвано следующими причинами:

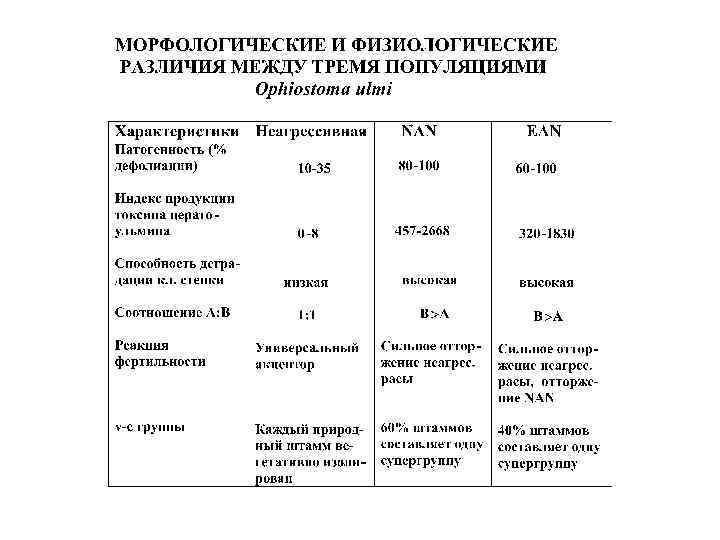
Расхождением по трофическим нишам v-с группы Fusarium oxysporum f. sp. vasinfectum, изолированные из корней и ризосферы хлопчатника в 3 х районах Израиля (Katan, 1988) Группы совместимости Число штаммов, выделенных из ткани корней ризосферы патогенные не патоген. v-c 1 v-c 2 236 0 0 47 138 0 0 210

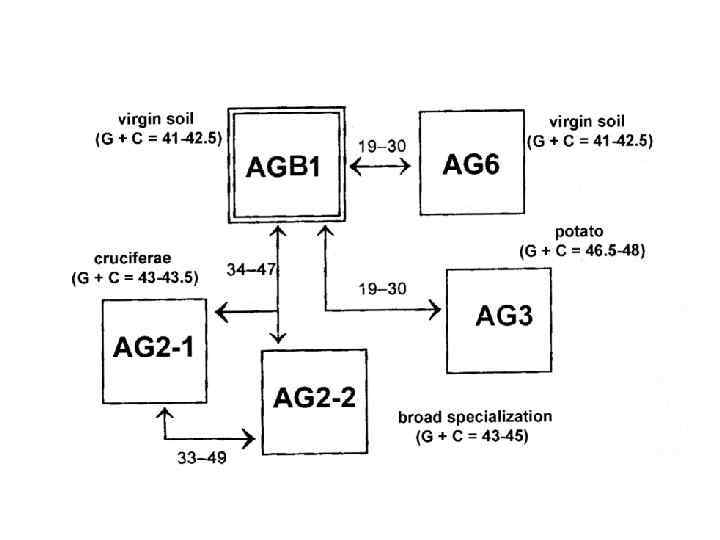
Расхождением по растениям-хозяевам Анастомозные группы Rhizoctonia solani


Поражением растений, находящихся на разных фазах развития

Во всех перечисленных случаях основным механизмом, приводящим к расхождению популяций и видообразованию, является ВН между адаптированными к разным условиям штаммами. Единственный путь генетических обменов в агамных грибных популяциях – миграция ядер между рядом растущими штаммами вследствие анастомозов гиф. В возникающих таким путем гетерокариотичных клетках возможны генетические рекомбинации (парасексуальный процесс). Миграцию ядер ограничивает ВН, препятствующая слияниям гиф или вызывающая гибель слившихся клеток штаммов, гетероаллельных по het-генам.

Благодаря этому грибная популяция разбивается на большое число групп, взаимно генетически изолированных и имеющих независимую эволюционную судьбу. Мозаичная структура популяций описана для многих грибов.

 предположили, что механизмом ассортативных")
А. В. Марков и А. М. Куликов (Известия РАН, 2006) предположили, что механизмом ассортативных скрещиваний, приводящих в симпатрическому видообразованию у животных, является иммунная система, белки которой могут наряду с половыми феромонами или в качестве последних влиять на выбор партнера и тем самым ограничивать гибридизацию между особями, адаптированными к разным условиям жизни. В соответствии с этой гипотезой можно полагать, что у грибов впервые возникли механизмы симпатрического видообразования, основанные на эксплуатации простейшей иммунной системы.

СПАСИБО ЗА ВНИМАНИЕ
incompat.ppt