Молекулярная биотехнология 3.ppt
- Количество слайдов: 169



Важнейшие методы молекулярной биотехнологии http: //molbiotech. ucoz. ru/

Важнейшие методы молекулярной биотехнологии • Химический синтез дезоксирибополинуклеотидов • Молекулярная гибридизация • Полимеразная цепная реакция • Секвенирование ДНК

Химический синтез ДНК • Общий принцип • ДНК-синтезаторы • Подход удобен для синтеза олигонуклеотидов, используемых в различных работах по генетической инженерии • Возможен химический синтез генов

Химический синтез ДНК: 1 -й нуклеотид

Структура нуклеозид-фосфорамидита

Присоединение 2 -го нуклеотида В 1

ДНК-синтезаторы • Это аппараты, в которых синтез ДНК осуществляется автоматически под контролем компьютера
 Праймеры 2) Зонды 3) Линкеры 4) Адапторы")
Для чего нужен химический синтез ДНК? 1) Праймеры 2) Зонды 3) Линкеры 4) Адапторы 5) Синтетические гены
, используемые для старта матричного синтеза ДНК с помощью")
Синтетические праймеры Олигонуклеотиды (20 -25 нт), используемые для старта матричного синтеза ДНК с помощью ДНК-полимераз 3’ 5’

Синтетические зонды • Зонд – меченый олигонуклеотид, специфически связывающийся с определённой нуклеотидной последовательностью и позволяющий ее обнаружить • Радиоактивные зонды • Флуоресцирующие зонды * 5’ ___________ 3’

Синтетические линкеры • Короткие двуцепочечные олигонуклеотиды, позволяющие соединить две молекулы ДНК, имеющие разные концы • ___________

Синтетические адапторы • Короткие дц олигонуклеотиды, позволяющие создать липкий конец в молекуле ДНК _________
 •")
Синтетические гены • Химический синтез ДНК с помощью ДНКсинтезаторов (олигонуклеотиды до 50 нт) • Сборка молекулы из фрагментов d. NTP, ДНК-полимераза, лигаза • Включение синтетического гена в плазмиду
 • Синтез")
Работы К. Вентера по синтетической геномике • Синтез генома микоплазмы (1000 тпн) • Синтез митохондриального генома (20 тпн)
 Это метод амплификации (увеличения количества) определённой нуклеотидной последовательности")
Полимеразная цепная реакция (ПЦР) Это метод амплификации (увеличения количества) определённой нуклеотидной последовательности

Разработка ПЦР • Химический синтез праймеров • Выделение термостабильной ДНКполимеразы из термофильной бактерии Thermus aquaticus – Taq I – А. Каледин и С. Городецкий (1980) • Разработка принципа ПЦР К. Муллисом (1986)

Главные реактивы для ПЦР • • • Исходная ДНК Праймеры Термостабильная ДНК-полимераза Дезоксирибонуклеотиды (d. NTP) Буферный раствор
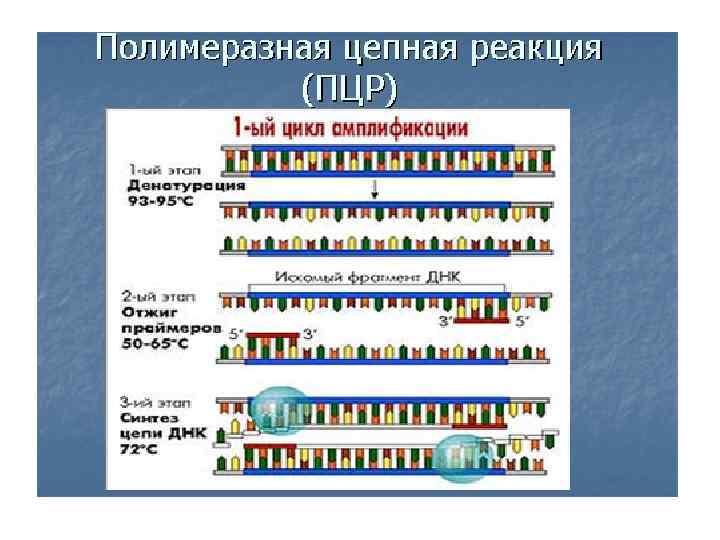

Принцип ПЦР

• pcr_animation. MOV

Электрофорез продуктов ПЦР в агарозе 1 - маркер 2, 3, 4, 5 – gag 1/gag 2 6, 7, 8, 9 – pol 1/pol 2

ПЦР в реальном времени Это ПЦР, продукты которой выявляют не электрофоретическим методом, а с помощью флуоресцирующей метки, которая нарастает по мере увеличения продуктов реакции
, который флуоресцирует")
Варианты ПЦР в реальном времени: метод интеркаляции • Используется флуоресцентный краситель (SYBR), который флуоресцирует после интеркаляции в дц. ДНК (чем больше ДНК, тем сильнее флуоресценция)
")
Real-Time PCR (ПЦР в реальном времени)

Варианты ПЦР в реальном времени: метод зондов • Использование меченых зондов, флуоресценция которых проявляется после связывания с ДНК (чем больше ДНК, тем сильнее флуоресценция)

Молекулярный маяк
-ПЦР • С помощью обратной транскриптазы синтезируют к. ДНК-копии молекул РНК • В смеси")
ОТ(RT)-ПЦР • С помощью обратной транскриптазы синтезируют к. ДНК-копии молекул РНК • В смеси образовавшихся молекул к. ДНК с помощью ПЦР амплифицируется заданная последовательность с помощью подобранной пары праймеров РНК оц. ДНК дц. ДНК амплификация определённого фрагмента ДНК

Возможности ПЦР • Получение нужных вставок для генетической инженерии и клонирование генов, введение нужных сайтов рестрикции в концы амплифицированной последовательности • Получение материала для секвенирования ДНК

Клонирование генов с помощью ПЦР • Выбор праймеров на основании данных банка нуклеотидных последовательностей • Приготовление препарата РНК • Постановка ОТ-ПЦР с праймерами, содержащими нужный сайт рестрикции • Клонирование полученного продукта в нужный вектор
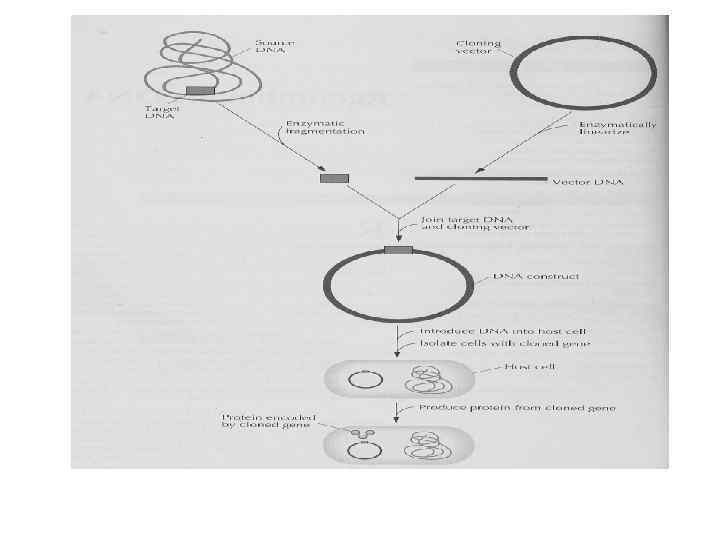

Диагностическое значение ПЦР • Индикация определённых фрагментов ДНК • Диагностика инфекционных болезней • Определение вида пищевого сырья

Выявление ДНК-провируса бычьего иммунодефицита в клетках крови КРС
")
Секвенирование ДНК Это определение порядка нуклеотидов в молекуле ДНК (чтение генетического текста)

Принцип секвенирования ДНК

Радиоактивная метка 5’-конца
 • Модификация пуринов (G+A) – пиперидинформат • Модификация гуанина")
Химическое секвенирование ДНК (метод Максама-Гилберта) • Модификация пуринов (G+A) – пиперидинформат • Модификация гуанина (G) – диметилсульфат • Модификация пиримидинов (C+T) - гидразин • Модификация цитозина (C) – гидразин +Na. Cl

Принцип химического секвенирования

Результат химического секвенирования
 Для секвенирования необходимы: • ДНК-полимераза • d. NTP (A,")
Ферментативное секвенирование ДНК (метод Сэнгера) Для секвенирования необходимы: • ДНК-полимераза • d. NTP (A, G, C, T) • Дидезоксинуклеотиды (dd. ATP, dd. GTP, dd. CTP, dd. TTP)

Принцип ферментативного секвенирования

Ферментативное секвенирование ДНК

Метка dd. NTP • Радиоактивная • Флуоресцентная

ДНК-секвенаторы • Аппараты для автоматического секвенирования ДНК

ГЕНОМНЫЕ ПРОЕКТЫ • • ГЕНОМ ДРОЗОФИЛЫ ГЕНОМ H. INFLUENZAE ГЕНОМ ЧЕЛОВЕКА ГЕНОМ МЫШИ ГЕНОМ E. COLI ГЕНОМ ДРОЖЖЕЙ ГЕНОМ C. ELEGANS ГЕНОМЫ МНОГИХ ВИРУСОВ

Тaqman

Экспрессия рекомбинантных генов в прокариотических клетках

Одна из главных целей молекулярной биотехнологии – получение продуцентов биологически активных белков с помощью генетической инженерии

Рекомбинантная ДНК и рекомбинантные гены • Рекомбинантная ДНК – это ДНК, состоящая из вектора и вставки • Встроенные гены в составе рекомбинантной ДНК называются рекомбинантными генами
 • Синтез")
Экспрессия рекомбинантных генов • Синтез м. РНК на матрице рекомбинантных генов (транскрипция) • Синтез белков, кодируемых рекомбинантными генами (трансляция)

Рекомбинантные белки • Белки, экспрессируемые генами в составе рекомбинантной ДНК • Некоторые из этих белков полностью соответствуют природному белку • Некоторые рекомбинантные белки являются слитыми белками • Некоторые рекомбинантные белки имеют замены по отдельным аминокислотам

Что необходимо для эффективного получения рекомбинантных белков • Правильная биологическая система, позволяющая использовать ферментёры (биореакторы) • Эффективный аппарат транскрипции и трансляции • Правильная посттрансляционная модификация рекомбинантного белка • Эффективные методы очистки рекомбинантного белка

Ферментёры, или биореакторы • Это аппараты для промышленного культивирования микроорганизмов или клеток многоклеточных организмов • В ферментёрах поддерживается оптимальная температура, аэрация, р. Н, обеспечивается автоматическая подача питательной среды, получение и сбор клеток, подготовка аппарата к следующему циклу
 • Эукариоты (дрожжи) •")
Биологические системы, пригодные для использования в ферментёрах • Прокариоты (микроорганизмы) • Эукариоты (дрожжи) • Эукариотические клетки многоклеточных организмов в культуре

Прокариоты • Наиболее удобная биологическая система для промышленного получения продуктов – прокариоты • Наиболее отработанные прокариотические системы – E. coli (кишечная палочка) и B. subtilis (сенная палочка)
")
Про- и эукариотические клетки (схема)

Преимущества прокариотических систем • Дешёвая питательная среда • Быстрое размножение клеток • Возможность культивирования в больших объёмах среды • Максимальный экономический эффект

Недостатки прокариотических систем • Невозможность экспрессии интронированных эукариотических генов из-за отсутствия сплайсинга у прокариот • Невозможность правильной посттрансляционной модификации некоторых белков (протеолиз, фосфорилирование, гликозилирование, ацетилирование и др. ) • Нестабильность многих чужеродных белков в прокариотах

Е. coli • • Грамотрицательная бактерия, 1 х0. 5 мкм Геном – кольцевая дц. ДНК, 4 х106 п. н. Время деления – от 30 мин. Хорошо охарактеризована генетически, имеются коллекции мутантов • Имеются плазмиды и вирусы (фаги) для работы с Е. coli

Штамм E. coli K 12 • Специальный штамм E. coli, неспособный сохраняться во внешней среде • Имеется коллекция мутантов • Используется в работах по генетической инженерии

Этапы получения рекомбинантных белков • Подготовка гена • Подготовка вектора с необходимыми регуляторными элементами • Получение и накопление рекомбинантной плазмиды • Трансфекция, трансформация и селекция трансформированных клеток • Получение рекомбинанта-продуцента • Наращивание бактериальной массы • Индукция синтеза целевого белка • Очистка рекомбинантного белка

Как подготовить гены для их экспрессии в прокариотах • Используют клонированные гены • Прокариотические гены используются в их натуральном виде (за исключением промотора) • Эукариотические гены используются в виде к. ДНК • Гены получают из соответствующих библиотек или с помощью ПЦР

Два типа векторов • Клонирующие векторы • Экспрессирующие векторы

Векторы экспрессии • Для экспрессии генов используют те же векторы, что и для клонирования генов (плазмиды, фаги) • Специальное внимание уделяется регуляторным элементам, которые должны обеспечивать эффективную экспрессию целевого гена

Экспрессирующий прокариотический вектор

Регуляторные элементы для экспрессии генов в прокариотах • • • Промотор Оператор Последовательность Шайн-Дельгарно Инициаторный ATG кодон Терминирующий кодон (ТАА, ТАG, ТGА) Терминатор транскрипции

Регуляторные элементы для экспрессии генов в прокариотах P O ATG SD TC TT Gene м. РНК Полипептид

Последовательность Шайн. Дельгарно

Прокариотический промотор старт 5’TAGTG TATTGACA TGATAGAAGCACTCTAC TATATT CTCAATAACG 3’ -35 последов. -10 последов. Прибнов бокс -1 +1

Поиск промоторов

Два типа промоторов • Конститутивные – работают постоянно • Индуцибельные – работают при определенных условиях • Для наработки белков необходимо использовать индуцибельные промоторы, потому что накапливаемый в большом количестве белок помешает бактериям размножаться
 • UV 5 (модифицированный промотор lacоперона) •")
Индуцибельные прокариотические промоторы • Lac (промотор lac-оперона) • UV 5 (модифицированный промотор lacоперона) • Trp (промотор триптофанового оперона) • Tac (-35 область из trp, -10 из lacпромотора • PL (промотор фага лямбда)

Lac-оперон
 • Позитивная регуляция")
Два способа регуляции lacоперона • Негативная регуляция – lac-репрессор (ингибитор, I) • Позитивная регуляция – CAP-c. AMP (комплекс белка, активирующего катаболизм, с циклическим АМФ)

Регуляция lac-оперона

Регуляторная область lac-оперона

Образование циклического АМР

Условия активации lac-промотора

Фаговые PL - промотор и с. I-репрессор • Активность PL - промотора подавляется с. I-репрессором • Используют мутантный с. I-репрессор, теряющий активность при 420

Триптофановый промотор • Индуцибельный промотор, содержит оператор • Контролирует работу триптофанового оперона, продукты которого осуществляют синтез триптофана • Ген ингибитора находится далеко от оперона • При наличии триптофана в среде ингибитор+Trp выключает оперон

Искусственное выключение промотора • Lac-промотор: среда с глюкозой и без лактозы • Trp-промотор: среда с триптофаном • PL -промотор – культивирование при 370
 • Trp-промотор: 3 -β-индолилакриловая кислота •")
Искусственная индукция промоторов • Lac-промотор: изопропил-β -Dтиогалактозид (IPTG) • Trp-промотор: 3 -β-индолилакриловая кислота • PL -промотор – нагревание до 420

Нестабильность рекомбинантных белков в бактериях • Многие рекомбинантные белки в бактериях подвергаются ускоренному протеолитическому распаду • Для устранения ускоренного протеолиза заменяют аминокислоту на N-конце полипептидной цепи (правило N-конца) или • Получают т. н. слитый белок

Вектор для получения слитого белка

Очистка рекомбинантного белка • При накоплении белка в бактериях их нужно разрушить ультразвуком • Осадить дебрис центрифугированием • Фракционировать смесь полученных белков с помощью колоночной хроматографии
 • Гель-фильтрация (делит молекулы")
Виды хроматографии • Ионообменная хроматография (делит молекулы белка по заряду) • Гель-фильтрация (делит молекулы белка по размеру) • Аффинная хроматография (делит по сродству к определённому веществулиганду, закреплённому на матрице)
 • Катионообменники (фосфоцеллюлоза)")
Ионообменная хроматография • Разделение по заряду • Анионообменники (ДЕАЕ-целлюлоза) • Катионообменники (фосфоцеллюлоза)
 Гели на основе")
Гель-фильтрация • • Разделение по размеру Гели на основе декстрана (сефадексы) Гели на основе агарозы Гели на основе синтетических веществ (полиакриламид, трисакрил и др. )

Аффинная хроматография • Разделение по связыванию с лигандом, фиксированным на матрице • Матрицы (целлюлоза, агароза и др. ) • Лиганды (нуклеиновые к-ты, антитела и др. )

Прокариотические белки, получаемые в рекомбинантных бактериях • ДНК-полимераза I Е. coli • ДНК-лигаза Е. coli • Рестриктазы и др.

Эукариотические белки, получаемые в рекомбинантных бактериях • • Обратная транскриптаза Гормоны роста Интерфероны (альфа-, бета-, гамма-) Инсулин Цитокины (IL-2, TNF и др. ) Факторы свёртываемости крови Тканевой активатор плазминогена и урокиназа • Вирусные антигены и др.

Экспрессия рекомбинантных генов в эукариотических клетках
")
Про- и эукариотические клетки (схема)

Недостатки прокариотических систем • Невозможность экспрессии интронированных эукариотических генов из-за отсутствия сплайсинга у прокариот • Невозможность правильной пострансляционной модификации белков (протеолиз, фосфорилирование, гликозилирование, ацетилирование и др. ) • Нестабильность многих чужеродных белков в прокариотах
 •")
Эукариотические клетки, используемые как продуценты рекомбинантных белков • Дрожжи (пекарские дрожжи, метилотрофные дрожжи) • Культуры клеток насекомых • Культуры клеток млекопитающих и птиц

Преимущества эукариотических клеток • Обеспечивается правильное посттрансляционное процессирование рекомбинантных белков, особенно в клетках животных • Дрожжи сочетают достоинства эукариотических и прокариотических клеток

Недостатки эукариотических клеток • Использование эукариотических клеток, особенно клеток животных, менее технологично из-за более медленного размножения клеток и требует больших затрат Время удвоения: • Бактерии – 20 -30 мин • Дрожжи – 1, 5 -2, 5 ч. • Клетки животных – 8 -12 ч.

Пекарские дрожжи. Saccharomyces cerevisiae • • Размер клетки ~10 мкм Геном 1. 4 x 107 пн 16 хромосом Размножается почкованием и половым способом • Время удвоения – 1, 5 -2, 5 ч. • Образует колонии на агаре, легко культивируется в биореакторах

Векторы для дрожжевой системы • Вирусы дрожжей не описаны • Обнаружен один вид плазмиды – 2 мкм ДНК • На основе этой плазмиды получены векторы для дрожжей, в том числе челночные плазмиды

Челночный вектор • Способен размножаться и в бактерии и в эукариотической клетке • Содержит два участка ori – про- и эукариотический • Содержит селективные маркеры для прокариотических и для эукариотических клеток

Челночный вектор для дрожжей

Векторы на основе ARS элементов

Трансфекция дрожжевых клеток • Клетки дрожжей имеют полисахаридную клеточную стенку, окружающую цитоплазматическую мембрану. Эта стенка не пропускает молекулы ДНК. • Клеточную стенку разрушают полисахаридрасщепляющим ферментом и получают сферопласты, которые обрабатывают Ca. Cl 2 и трансфицируют ДНК. Клеточная стенка восстанавливается при культивировании.

Неинтегративная и интегративная трансформация • Неинтегративная трансформация клетки достигается за счет размножения плазмиды в клеточном ядре. • Интегративная трансформация – результат интеграции части плазмиды с интересующим геном в хромосому.

Стабильная и нестабильная трансформация клеток • В отсутствие селективного давления автономные плазмиды легко утрачиваются (нестабильная трансформация) • Интегрированный ген обычно стабильно сохраняется в геноме, если не влияет негативно на жизнеспособность клетки (стабильная трансформация)

Стабильная трансформация дрожжевых клеток
")
Искусственная дрожжевая хромосома (YAC)

Системы селекции для дрожжей • Использование мутантов дрожжей, дефектных по генам синтеза аминокислот (лейцин, гистидин, триптофан) и азотистых оснований. Маркерами являются гены LEU, HIS, TRP, URA и др. • Использование гена NEO устойчивости к антибиотику G 418 (генетицину)

Регуляторные элементы, контролирующие эукариотические рекомбинантные гены ATG TC Poly-A signal Промотор AAAAA м. РНК Полипептид

Регуляторные элементы для экспрессии генов в прокариотах P O ATG SD TC TT Gene м. РНК Полипептид
 • Промотор гена алкоголь-оксидазы")
Дрожжевые промоторы • Промотор гена CYC 1 (кодирует цитохром С) • Промотор гена алкоголь-оксидазы (AOX 1)

Метилотрофные дрожжи • Метилотрофные дрожжи используют метиловый спирт в качестве источника углерода • Они накапливаются в большом количестве • Удобны для наработки рекомбинантных белков под контролем промотора гена алкоголь-оксидазы (АОХ 1)

Использование метилотрофных дрожжей Pichia pastoris для наработки белков

Стабильная трансформация метилотрофных дрожжей плазмидой, содержащей ген HBs. Ag
")
Рекомбинантные продукты, полученные в дрожжевых системах • • HBs. Ag (вакцина против гепатита В) Белки малярийного плазмодия Оболочечный антиген HIV Лекарственные белки человека (эпидермальный фактор роста, альфа 1 антитрипсин, факторы свёртываемости крови и др. )

Клетки животных в культуре • Первичные и вторичные культуры • Диплоидные штаммы клеток • Перевиваемые линии клеток

Векторы для клеток животных • Вирусы (поксвирусы, аденовирусы, полиома- и папилломавирусы, ретровирусы, бакуловирусы и др. ) • Прокариотические плазмиды

Схема плазмидного вектора для клеток млекопитающих

Введение плазмидных векторов в клетки • Кальций-фосфатный метод (Ca. Cl 2+ Na 3 PO 4 Ca 3(PO 4)2 • Полибрен, ДЕАЕ-декстран • Липосомы • Электропорация • Бомбардировка частичками золота
 • Ген устойчивости к")
Селекция трансформированных клеток • Ген устойчивости к G 418 (генетицину) • Ген устойчивости к пуромицину • Устойчивость к аминоптерину или метотрексату • Устойчивость к микофеноловой кислоте • Окраска (lac. Z) или флуоресценция (флуоресцентные белки)

Транзиторная экспрессия генов в эукариотических клетках • В большинстве трансфицированных клеток происходит транзиторная (временная, проходящая) экспрессия трансфицированного гена, достигающая максимума через 2 -3 дня и спадающая к 6 -7 дню после трансфекции. • Небольшая часть трансфицированных клеток становится стабильно трансформированной.

Стабильная трансформация клеток животных рекомбинантными плазмидами • Попав в клеточное ядро, рекомбинантные плазмиды способны встраиваться в геном клетки, что ведет к стабильной трансформации • Встройка обычно происходит по принципу случайной рекомбинации • Существуют способы увеличения числа копий встроенного гена

Использование гена дигидрофолатредуктазы для усиление экспрессии целевого гена

Увеличения числа копий введенного гена • Выращивание клеток в присутствии аминоптерина ведет к амплификации участка генома, содержащего ген дигидрофолатредуктазы (ДГФР) и прилежащие гены • При этом резко увеличивается число копий гена ДГФР и расположенных рядом с ним генов. Количество продукта значительно увеличивается.

Эритропоэтин • Важное лекарство – эритропоэтин – было получено в клетках СНО после амплификации гена эритропоэтина, встроенного рядом с геном ДГФР

Геном вируса SV 40

Размножение плазмид в клетках животных • Обезьяньи СOS клетки Я. Глузмана • Эти клетки продуцируют Т антиген вируса SV 40 • T антиген, связываясь с ori SV 40, вызывает репликацию вирусной ДНК клеточной репликационной машиной • Если ori SV 40 включен в плазмиду, она будет реплицироваться в COS клетках

Бакуловирусная система • Вирус множественного ядерного полиэдроза – ДНК-содержащий вирус с кольцевой дц. ДНК ~120 тпн, поражает насекомых свыше 30 видов, вызывает образование полиэдров в ядрах клеток • Полиэдры построены из вирусного белка полиэдрина. • Ген полиэдрина имеет мощный промотор

Плазмидный бакуловирусный вектор

Получение рекомбинантного бакуловируса • Клетки заражают «диким» бакуловирусом • Трансфицируют зараженные клетки плазмидой, содержащей участок вирусной ДНК и ген интереса • В результате гомологичной рекомбинации ген интереса встраивается в вирусную ДНК • Затем отбирают и клонируют рекомбинантные вирионы методом бляшек

Встройка целевого гена в бакуловирусную ДНК

Получение рекомбинантного белка в бакуловирусной системе • Культуру клеток насекомых заражают рекомбинантным бакуловирусом • Целевой белок образуется в заражённых клетках и может быть выделен и очищен из культуральной жидкости или клеточного экстракта

Рекомбинантные белки, полученные в бакуловирусной системе • В системе получены сотни рекомбинантных белков различного назначения • Недавно была разработана вакцина против папилломавирусных инфекций, вызывающих рак шейки матки (антигены для этой вакцины нарабатываются в бакуловирусной системе)

Трансгенные животные

Биологические системы, используемые молекулярной биотехнологией • • Вирусы Прокариоты Дрожжи Культура клеток животных и человека (клетки насекомых, птиц, млекопитающих), растений • Животные • Растения

Трансгенные животные • Трансгенные животные - это животные, в геном которых искусственно встроен чужеродный генетический материал • Чужеродный ген содержится во всех клетках трансгенного организма

ТРАНСГЕНЫ И ТРАНСГЕНЕЗ • ТРАНСГЕНЫ: ВСТРОЕННЫЙ ГЕНЕТИЧЕСКИЙ МАТЕРИАЛ ОБОЗНАЧАЕТСЯ ТЕРМИНОМ “ТРАНСГЕН” • ТРАНСГЕНЕЗ: ПРОЦЕСС ПОЛУЧЕНИЯ ТРАНСГЕННЫХ ОРГАНИЗМОВ ОБОЗНАЧАЕТСЯ ТЕРМИНОМ “ТРАНСГЕНЕЗ”
 – такие же, как для")
Подготовка генной конструкции для трансгенеза • Экспрессирующие векторы (плазмиды) – такие же, как для экспрессии генов в клетках животных. Они должны содержать регуляторные элементы (промоторы, энхенсеры и др. ), соответствующие поставленной задаче • Специальные конструкции для введения гена в заданную область генома

Регуляторные элементы, контролирующие эукариотические рекомбинантные гены ATG TC Poly-A signal Промотор AAAAA м. РНК Полипептид

Стабильная трансформация клеток животных трансгеном • В качестве трансгенов используют линейную ДНК, освобожденную от плазмидных последовательностей • Попав в клеточное ядро, трансгены способны встроиться в геном клетки, что ведет к стабильной трансформации трансфицированной клетки • Встройка обычно происходит по принципу случайной (негомологичной) рекомбинации

ВИДЫ ЖИВОТНЫХ, ИСПОЛЬЗУЕМЫЕ ДЛЯ ТРАНСГЕНЕЗА • • • ДРОЗОФИЛЫ ЗЕМНОВОДНЫЕ МЫШИ КРОЛИКИ КОЗЫ ОВЦЫ СВИНЬИ КОРОВЫ КУРЫ РЫБЫ

ОБЩАЯ СХЕМА ПОЛУЧЕНИЯ ТРАНСГЕННЫХ ЖИВОТНЫХ • • • ПОЛУЧЕНИЕ ОПЛОДОТВОРЕННЫХ ЯЙЦЕКЛЕТОК ВВЕДЕНИЕ КЛОНИРОВАННОГО ГЕНА В ЯДРО ОПЛОДОТВОРЕННОЙ ЯЙЦЕКЛЕТКИ ИМПЛАНТАЦИЯ ТАКИХ ЯЙЦЕКЛЕТОК В МАТКУ ПОДГОТОВЛЕННОЙ САМКИ ВЫЯВЛЕНИЕ ЖИВОТНЫХ, СОДЕРЖАЩИХ ТРАНСГЕН, СРЕДИ РОДИВШЕГОСЯ ПОТОМСТВА СКРЕЩИВАНИЕ ОСОБЕЙ, СОДЕРЖАЩИХ ТРАНСГЕН, ДЛЯ ВЫВЕДЕНИЯ ЛИНИИ ТРАНСГЕННЫХ ЖИВОТНЫХ

Введение трансгена • Введение трансгена с помощью ретровируса • Микроинъекции трансгена в яйцеклетку, микроманипуляторы

ТРАНСГЕННЫЕ МЫШИ • • ПОЛУЧЕНИЕ ОПЛОДОТВОРЕННЫХ ЯЙЦЕКЛЕТОК ОТ СУПЕРОВУЛИРОВАННЫХ САМОК (ДО 35 ЯЙЦЕКЛЕТОК ВМЕСТО ОБЫЧНЫХ 5 -10) МИКРОИНЪЕКЦИЯ ТРАНСГЕНА В МУЖСКОЙ ПРОНУКЛЕУС ИМПЛАНТАЦИЯ ИНОКУЛИРОВАННЫХ ЯЙЦЕКЛЕТОК В МАТКУ ПОДГОТОВЛЕННЫХ САМОК ПОЛУЧЕНИЕ И АНАЛИЗ ПОТОМСТВА НА НАЛИЧИЕ ТРАНСГЕНА (БЛОТГИБРИДИЗАЦИЯ И ПЦР)

Получение трансгенных мышей
 КЛЕТКИ • ЭС КЛЕТКИ – НЕДИФФЕРЕНЦИРОВАННЫЕ КЛЕТКИ, ДАЮЩИЕ НАЧАЛО ВСЕМ ТКАНЯМ")
ЭМБРИОНАЛЬНЫЕ СТВОЛОВЫЕ (ЭС) КЛЕТКИ • ЭС КЛЕТКИ – НЕДИФФЕРЕНЦИРОВАННЫЕ КЛЕТКИ, ДАЮЩИЕ НАЧАЛО ВСЕМ ТКАНЯМ ОРГАНИЗМА • ИСПОЛЬЗОВАНИЕ ДЛЯ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ МЫШЕЙ • ВВЕДЕНИЕ ТРАНСГЕНА В ЗАДАННОЕ МЕСТО ГЕНОМА

ПОЛУЧЕНИЕ ЭС КЛЕТОК • • ЭС КЛЕТКИ ПОЛУЧАЮТ ИЗ ВНУТРЕННЕЙ КЛЕТОЧНОЙ МАССЫ БЛАСТОЦИСТА КУЛЬТИВИРОВАНИЕ ЭТИХ КЛЕТОК ПОЗВОЛЯЕТ ПОЛУЧИТЬ ПЕРЕВИВАЕМЫЕ КЛЕТОЧНЫЕ ЛИНИИ, ПРИГОДНЫЕ ДЛЯ ПОЛУЧЕНИЯ ТРАНСГЕННЫХ ЖИВОТНЫХ

Источник стволовых клеток

ИСПОЛЬЗОВАНИЕ ЭС КЛЕТОК ДЛЯ ТРАНСГЕНЕЗА • ПОДГОТОВКА ЭС КЛЕТОК • ТРАНСФЕКЦИЯ ЭС КЛЕТОК ТРАНСГЕНОМ, ОТБОР ТРАНСФОРМИРОВАННЫХ КЛЕТОК • ВВЕДЕНИЕ ТРАНСФОРМИРОВАННЫХ ТРАНСГЕНОМ ЭС КЛЕТОК В БЛАСТОЦИСТЫ • ИМПЛАНТАЦИЯ БЛАСТОЦИСТОВ ПОДГОТОВЛЕННЫМ САМКАМ • ПОЛУЧЕНИЕ ПОТОМСТВА И ЕГО АНАЛИЗ НА НАЛИЧИЕ ТРАНСГЕНА

Введение трансгенов в ЭС клетки • Кальций-фосфатный метод (Ca. Cl 2+ Na 3 PO 4 Ca 3(PO 4)2 • Полибрен, ДЕАЕ-декстран • Липосомы • Электропорация

Получение трансгенных мышей с помощью стволовых клеток

ВВЕДЕНИЕ ТРАНСГЕНА В ЗАДАННОЕ МЕСТО ГЕНОМА • ОБЫЧНО ТРАНСГЕН НЕ ИМЕЕТ ОПРЕДЕЛЕННОГО АДРЕСА ДЛЯ ВСТРОЙКИ • ДЛЯ ВВЕДЕНИЯ ТРАНСГЕНА В ЗАДАННЫЙ УЧАСТОК ГЕНОМА ИСПОЛЬЗУЮТ ПРИНЦИП ГОМОЛОГИЧНОЙ РЕКОМБИНАЦИИ
 ВИРУСА ПРОСТОГО ГЕРПЕСА • TK ГЕНЫ ФОСФОРИЛИРУЮТ ТИМИДИН, ПРЕВРАЩАЯ ЕГО В")
ГЕН ТИМИДИНКИНАЗЫ (tk) ВИРУСА ПРОСТОГО ГЕРПЕСА • TK ГЕНЫ ФОСФОРИЛИРУЮТ ТИМИДИН, ПРЕВРАЩАЯ ЕГО В ТМР (Td. R-PO 4) • TK ГЕН ВПГ ОТЛИЧАЕТСЯ НИЗКОЙ СПЕЦИФИЧНОСТЬЮ И СПОСОБЕН ФОСФОРИЛИРОВАТЬ ЦИТИДИН И «НЕПРАВИЛЬНЫЕ» НУКЛЕОЗИДЫ (НАПР. , ГАНЦИКЛОВИР). • ФОСФОРИЛИРОВАННЫЙ ГАНЦИКЛОВИР УБИВАЕТ КЛЕТКУ

Целенаправленная встройка трансгена-I

Целенаправленная встройка трансгена-II
 Neo G 418 Neo")
Позитивная и негативная селекция Ген Ингибитор Рост клеток (позитивная селекция) Neo G 418 Neo + G 418 + (негативная селекция) tk HSV ганцикловир + tk HSV+ ганцикловир -

ИСПОЛЬЗОВАНИЕ ТРАНСГЕННЫХ МЫШЕЙ • • ИЗУЧЕНИЕ ФУНКЦИИ ГЕНОВ ПОИСК ТКАНЕСПЕЦИФИЧЕСКИХ ПРОМОТОРОВ

НОКАУТ ГЕНОВ У МЫШЕЙ • Нокаут гена – это его инактивация во всех клетках организма • Это достигается встройкой маркерного гена внутрь нокаутируемого гена

ТКАНЕСПЕЦИФИЧЕСКАЯ ЭКСПРЕССИЯ ГЕНОВ У ТРАНСГЕННЫХ МЫШЕЙ • Выявление промоторов, способных экспрессироваться в строго определенных тканях (в поджелудочной железе, в мозгу, в молочной железе и т. д. ) • Поставив онкоген под соответствующий промотор, можно вызвать опухоли у трансгенных мышей в определенной ткани или органе

ИСПОЛЬЗОВАНИЕ ТРАНСГЕННЫХ МЫШЕЙ • ИЗУЧЕНИЕ ФУНКЦИИ ГЕНОВ • ПОИСК ТКАНЕСПЕЦИФИЧЕСКИХ ПРОМОТОРОВ • СОЗДАНИЕ МОДЕЛЕЙ БОЛЕЗНЕЙ ЧЕЛОВЕКА (БОЛЕЗНЬ АЛЬЦГЕЙМЕРА ГЕН ПРЕДШЕСТВЕННИКА БЕТААМИЛОИДНОГО БЕЛКА; МЫШИ, ЧУВСТВИТЕЛЬНЫЕ К ВИРУСУ ПОЛИОМИЕЛИТА)

Болезнь Альцгеймера • Накопление сенильных бляшек в синапсисах нейронов • Бляшки содержат амилоид бета (Аβ), 4 к. Д • Ген предшественника Аβ кодирует белок предшественник Аβ (АРР) • Мутации в гене АРР ведут к накоплению Аβ • Мыши, трансгенные по мутантному гену АРР, проявляют симптомы болезни и характерные патологические изменения

МЫШИ, ЧУВСТВИТЕЛЬНЫЕ К ВИРУСУ ПОЛИОМИЕЛИТА • Рецептор полиовируса отсутствует в клетках мышей, поэтому они нечувствительны к этому вирусу • Мыши, трансгенные по гену рецептора полиовируса, чувствительны к нему и могут использоваться для контроля полиомиелитной вакцины вместо обезьян

ИСПОЛЬЗОВАНИЕ ТРАНСГЕННЫХ МЫШЕЙ • ИЗУЧЕНИЕ ФУНКЦИИ ГЕНОВ • ПОИСК ТКАНЕСПЕЦИФИЧЕСКИХ ПРОМОТОРОВ • СОЗДАНИЕ МОДЕЛЕЙ БОЛЕЗНЕЙ ЧЕЛОВЕКА (БОЛЕЗНЬ АЛЬЦГЕЙМЕРА - ГЕН ПРЕДШЕСТВЕННИКА БЕТА-АМИЛОИДНОГО БЕЛКА; МЫШИ, ЧУВСТВИТЕЛЬНЫЕ К ВИРУСУ ПОЛИОМИЕЛИТА) • МОДЕЛЬНЫЕ ЭКСПЕРИМЕНТЫ ПО ПРОДУКЦИИ БИОЛОГИЧЕСКИ АКТИВНЫХ ВЕЩЕСТВ (БАВ)

ПРОДУКЦИЯ БАВ В МОЛОЧНОЙ ЖЕЛЕЗЕ ТРАНСГЕННЫХ ЖИВОТНЫХ • МОЛОЧНАЯ ЖЕЛЕЗА - ФАБРИКА ПО • • • СИНТЕЗУ БЕЛКОВ (ДО 600 КГ/КОРОВА/ПЕРИОД ЛАКТАЦИИ). ГЛАВНЫЕ БЕЛКИ АЛЬФА-S 1 -КАЗЕИН АЛЬФА-S 2 -КАЗЕИН, БЕТА-КАЗЕИН КАППА-КАЗЕИН ЛАКТАЛЬБУМИН

ПРОМОТОРЫ, СПЕЦИФИЧНЫЕ ДЛЯ МОЛОЧНОЙ ЖЕЛЕЗЫ • ГЕНЫ КАЗЕИНОВ И ЛАКТАЛЬБУМИНА ИМЕЮТ ПРОМОТОРЫ, СПЕЦИФИЧНЫЕ ДЛЯ МОЛОЧНОЙ ЖЕЛЕЗЫ • ГЕН, КОНТРОЛИРУЕМЫЙ ТАКИМ ПРОМОТОРОМ, ЭКСПРЕССИРУЕТСЯ ИЗБИРАТЕЛЬНО В МОЛОЧНОЙ ЖЕЛЕЗЕ • ЕГО БЕЛКОВЫЙ ПРОДУКТ ВЫДЕЛЯЕТСЯ В МОЛОКО, ОТКУДА ОН МОЖЕТ БЫТЬ ПОЛУЧЕН В ЧИСТОМ ВИДЕ

• • • ТРАНСГЕННЫЕ СЕЛЬСКОХОЗЯЙСТВЕННЫЕ ЖИВОТНЫЕ КОЗЫ ОВЦЫ СВИНЬИ КОРОВЫ КУРЫ

НЕКОТОРЫЕ БИОЛОГИЧЕСКИ АКТИВНЫЕ БЕЛКИ, ПОЛУЧАЕМЫЕ С ПОМОЩЬЮ ТРАНСГЕННЫХ ЖИВОТНЫХ БЕЛОК • • • ЖИВОТНОЕ ТКАНЕВОЙ АКТИВАТОР ПЛАЗМИНОГЕНА - КОЗА УРОКИНАЗА КОЗА ФАКТОР IX СВЕРТЫВАНИЯ КРОВИ ОВЦА ЛАКТОФЕРРИН КОЗА КОРОВ А • ХИМОЗИН ОВЦА

ИЗМЕНЕНИЕ БИОЛОГИЧЕСКИХ СВОЙСТВ ТРАНСГЕННЫХ ЖИВОТНЫХ • УСТОЙЧИВОСТЬ К ИНФЕКЦИЯМ, • УСКОРЕНИЕ РОСТА (ГЕН ГОРМОНА РОСТА) • ДРУГИЕ СВОЙСТВА

НАУЧНОЕ И БИОТЕХНОЛОГИЧЕСКОЕ ЗНАЧЕНИЕ ТРАНСГЕННЫХ ЖИВОТНЫХ • • ОПРЕДЕЛЕНИЕ ФУНКЦИИ ТРАНСГЕНА СОЗДАНИЕ МОДЕЛЕЙ НЕКОТОРЫХ БОЛЕЗНЕЙ ЧЕЛОВЕКА ПОЛУЧЕНИЕ БИОЛОГИЧЕСКИ АКТИВНЫХ БЕЛКОВ УЛУЧШЕНИЕ СВОЙСТВ СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ

ПЕРСПЕКТИВЫ • СОЗДАНИЕ НОВЫХ ПОРОД СЕЛЬСКОХОЗЯЙСТВЕННЫХ ЖИВОТНЫХ С РАЗЛИЧНЫМИ ПОЛЕЗНЫМИ СВОЙСТВАМИ • СУЩЕСТВЕННОЕ РАСШИРЕНИЕ АССОРТИМЕНТА БАВ, ПОЛУЧАЕМЫХ В ТРАНСГЕННЫХ ЖИВОТНЫХ
Молекулярная биотехнология 3.ppt