

ВАКУОЛЯРНАЯ СИСТЕМА_ядерная оболочка.ppt
- Количество слайдов: 30
 ВАКУОЛЯРНАЯ СИСТЕМА КЛЕТКИ
ВАКУОЛЯРНАЯ СИСТЕМА КЛЕТКИ
 и экзоцитоз (б)") Везикулярный перенос: Эндоцитоз (а) и экзоцитоз (б)
Везикулярный перенос: Эндоцитоз (а) и экзоцитоз (б)
 и пиноцитоза (б)") Схема фагоцитоза (а) и пиноцитоза (б)
Схема фагоцитоза (а) и пиноцитоза (б)
: • неспецифический (конститутивный, постоянный); • специфический (рецепторный).") Эндоцитоз (пиноцитоз и фагоцитоз): • неспецифический (конститутивный, постоянный); • специфический (рецепторный).
Эндоцитоз (пиноцитоз и фагоцитоз): • неспецифический (конститутивный, постоянный); • специфический (рецепторный).
. а — пиноцитозная ямка и вакуоли; б — пиноцитоз") Эндоцитоз (фото С. М. Коломиной). а — пиноцитозная ямка и вакуоли; б — пиноцитоз на поверхности макрофага, видны выросты цитоплазмы, образующие складки 1 2 а 1 — пиноцитозная ямка; 2 — пиноцитозные вакуоли; 3 — эндосомы; 4 — ядро; 5 — аппарат Гольджи 2 3 5 4 б а - инвагинации плазматической мембраны (кишечный эпителий, эндотелий амебы) б - выросты в виде складок (фагоциты, фибробласты)
Эндоцитоз (фото С. М. Коломиной). а — пиноцитозная ямка и вакуоли; б — пиноцитоз на поверхности макрофага, видны выросты цитоплазмы, образующие складки 1 2 а 1 — пиноцитозная ямка; 2 — пиноцитозные вакуоли; 3 — эндосомы; 4 — ядро; 5 — аппарат Гольджи 2 3 5 4 б а - инвагинации плазматической мембраны (кишечный эпителий, эндотелий амебы) б - выросты в виде складок (фагоциты, фибробласты)
 Окаймленные ямки и окаймленные пузырьки Последовательные стадии образования и отщепления окаймленного пузырька 1 — плазматическая мембрана; 2 — интегральные белки; 3 — кэпирующие белковые частицы; 4 — клатрин; 5 — динамин а — вид со стороны цитозоля; б — трискелеоны на поверхности пузырька
Окаймленные ямки и окаймленные пузырьки Последовательные стадии образования и отщепления окаймленного пузырька 1 — плазматическая мембрана; 2 — интегральные белки; 3 — кэпирующие белковые частицы; 4 — клатрин; 5 — динамин а — вид со стороны цитозоля; б — трискелеоны на поверхности пузырька
 Фракция лизосом состоит из пузырьков размером 0, 2 - 0,") Лизосомы (Де Дюв, 1955) Фракция лизосом состоит из пузырьков размером 0, 2 - 0, 4 мкм, ограниченных одиночной мембраной (около 7 нм), с очень разнородным содержанием внутри. Содержат около 40 гидролитических ферментов: протеиназы, нуклеазы, гликозидазы, фосфорилазы, фосфатазы, сульфатазы. Оптимум действия при р. Н = 5. Надежным маркёром лизосом является фермент кислая фосфатаза
Лизосомы (Де Дюв, 1955) Фракция лизосом состоит из пузырьков размером 0, 2 - 0, 4 мкм, ограниченных одиночной мембраной (около 7 нм), с очень разнородным содержанием внутри. Содержат около 40 гидролитических ферментов: протеиназы, нуклеазы, гликозидазы, фосфорилазы, фосфатазы, сульфатазы. Оптимум действия при р. Н = 5. Надежным маркёром лизосом является фермент кислая фосфатаза
 Морфологическая неоднородность лизосом первичные лизосомы вторичные лизосомы остаточные тельца аутофагосомы
Морфологическая неоднородность лизосом первичные лизосомы вторичные лизосомы остаточные тельца аутофагосомы
 Вторичные лизосомы в клетках культуры СПЭВ. 1 — лизосомы; 2 — жировые капли
Вторичные лизосомы в клетках культуры СПЭВ. 1 — лизосомы; 2 — жировые капли
 Образование лизосом и их участие в клеточных процессах 1 — синтез гидролитических ферментов в ЭПР; 2 — переход их в АГ; 3 — образование первичных лизосом; 4 — выброс и использование (5) гидролаз при внеклеточном расщеплении; 6 — эндоцитозные вакуоли: 7 — слияние с ними первичных лизосом; 8 — образование вторичных лизосом; 9 —телолизосомы; 10 — экскреция остаточных телец; 11 — первичные лизосомы принимают участие в образовании аутофагосомы (12)
Образование лизосом и их участие в клеточных процессах 1 — синтез гидролитических ферментов в ЭПР; 2 — переход их в АГ; 3 — образование первичных лизосом; 4 — выброс и использование (5) гидролаз при внеклеточном расщеплении; 6 — эндоцитозные вакуоли: 7 — слияние с ними первичных лизосом; 8 — образование вторичных лизосом; 9 —телолизосомы; 10 — экскреция остаточных телец; 11 — первичные лизосомы принимают участие в образовании аутофагосомы (12)
 и листа табака (б)") Пероксисомы Строение пероксисом в клетках печени (а) и листа табака (б)
Пероксисомы Строение пероксисом в клетках печени (а) и листа табака (б)
 САМОСТОЯТЕЛЬНО: Специализация пероксисом у растений Сферосомы Протеосомы Вакуоли растительных клеток Включения
САМОСТОЯТЕЛЬНО: Специализация пероксисом у растений Сферосомы Протеосомы Вакуоли растительных клеток Включения
 Общая схема вакуолярной системы клетки 1 — ядерная оболочка; 2 — гранулярный эндоплазматический ретикулум (ЭПР); 3 — переходная зона (ЭПР-АГ-комплекс); 4 — перенос от ЭПР к аппарату Гольджи; 5 — проксимальные участки (цис) АГ; 6 — средняя часть (мед) АГ; 7 — дистальная (транс) часть АГ; 8 — транс-сеть АГ; 9 — возвратный путь вакуолей АГ; 10 — отделение первичных лизосом; 11 — постоянная экскреция (секреция); 12 — сигнальная секреция; 13 — эндоцитоз; 14 — эндосома; 15 — вторичная лизосома; 16 — возврат лизосомных мембан; 17 — возврат рецепторов в плазматическую мембрану; 18 — гладкий эндоплазматический ретикулум
Общая схема вакуолярной системы клетки 1 — ядерная оболочка; 2 — гранулярный эндоплазматический ретикулум (ЭПР); 3 — переходная зона (ЭПР-АГ-комплекс); 4 — перенос от ЭПР к аппарату Гольджи; 5 — проксимальные участки (цис) АГ; 6 — средняя часть (мед) АГ; 7 — дистальная (транс) часть АГ; 8 — транс-сеть АГ; 9 — возвратный путь вакуолей АГ; 10 — отделение первичных лизосом; 11 — постоянная экскреция (секреция); 12 — сигнальная секреция; 13 — эндоцитоз; 14 — эндосома; 15 — вторичная лизосома; 16 — возврат лизосомных мембан; 17 — возврат рецепторов в плазматическую мембрану; 18 — гладкий эндоплазматический ретикулум
 ЯДРО Браун, 1833 г.
ЯДРО Браун, 1833 г.
 ЯДЕРНАЯ ОБОЛОЧКА В состав ядерной оболочки входят наружная и внутренняя мембраны, ядерный поровый комплекс и ламина (или ядерная пластинка).
ЯДЕРНАЯ ОБОЛОЧКА В состав ядерной оболочки входят наружная и внутренняя мембраны, ядерный поровый комплекс и ламина (или ядерная пластинка).
 Наружная ядерная мембрана является частью гранулярной эндоплазматической сети. На её поверхности расположены рибосомы
Наружная ядерная мембрана является частью гранулярной эндоплазматической сети. На её поверхности расположены рибосомы
 Внутренняя ядерная мембрана Снаружи граничит с перинуклеарной цистерной, изнутри отделена от содержимого ядра ядерной пластинкой или ламиной. В местах слияния двух мембран расположены ядерные поры.
Внутренняя ядерная мембрана Снаружи граничит с перинуклеарной цистерной, изнутри отделена от содержимого ядра ядерной пластинкой или ламиной. В местах слияния двух мембран расположены ядерные поры.
 Ядерная пластинка или ламина Толщина ламины – 80 -300 нм. Ламина сформирована последовательностью одинаково ориентированных промежуточных филаментов, называемых ламинами. Молекулярная масса ламинов — примерно 60 -75 к. Да, это димеры, организующиеся в полимеры.
Ядерная пластинка или ламина Толщина ламины – 80 -300 нм. Ламина сформирована последовательностью одинаково ориентированных промежуточных филаментов, называемых ламинами. Молекулярная масса ламинов — примерно 60 -75 к. Да, это димеры, организующиеся в полимеры.
 Функции ламины: Ламины могут связываться с белками хроматина. Они необходимы для пространственной организации комплексов ядерных пор. Вовлечены в процессы конденсации хроматина и репрессии генов вблизи ядерной периферии. Ламины В-типа участвуют в процессе роста ядерной оболочки.
Функции ламины: Ламины могут связываться с белками хроматина. Они необходимы для пространственной организации комплексов ядерных пор. Вовлечены в процессы конденсации хроматина и репрессии генов вблизи ядерной периферии. Ламины В-типа участвуют в процессе роста ядерной оболочки.
 Мутации гена, кодирующего ламин А являются причиной тяжелейшего генетического заболевания, характеризующегося комплексом изменений кожи и внутренних органов, обусловленных преждевременным старением организма – детской прогерии.
Мутации гена, кодирующего ламин А являются причиной тяжелейшего генетического заболевания, характеризующегося комплексом изменений кожи и внутренних органов, обусловленных преждевременным старением организма – детской прогерии.
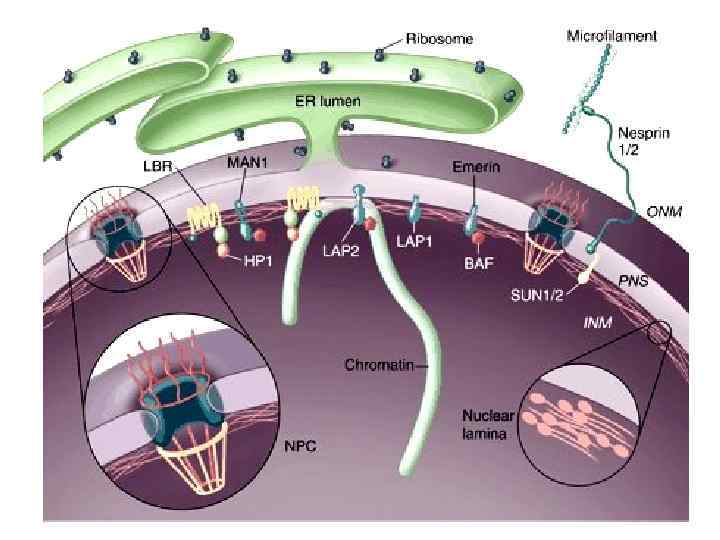
 Белки, связанные с ламинами В состав ядерной ламины входят белки, связанные с ламинами, такие как рецептор ламина B (LBR), ламинассоциированные полипептиды LAP 1 и LAP 2, имерин, MAN 1, отефин, несприн, UNC-84 и др. Некоторые из этих трансмембранных полипептидов связывают белки хроматина или ДНК. Важной функцией полипептидов ламины является дезинтеграция ядерной оболочки в процессе митоза
Белки, связанные с ламинами В состав ядерной ламины входят белки, связанные с ламинами, такие как рецептор ламина B (LBR), ламинассоциированные полипептиды LAP 1 и LAP 2, имерин, MAN 1, отефин, несприн, UNC-84 и др. Некоторые из этих трансмембранных полипептидов связывают белки хроматина или ДНК. Важной функцией полипептидов ламины является дезинтеграция ядерной оболочки в процессе митоза
 Структура ядерного порового комплекса Ядерный поровый комплекс состоит из различных белков нуклеопоринов и образует канал, регулирующий ядерно-цитоплазматический транспорт различных типов РНК, мембранных белков и растворимых белков. Крупные молекулы через ЯПК транспортируются специальными белками кариоферинами
Структура ядерного порового комплекса Ядерный поровый комплекс состоит из различных белков нуклеопоринов и образует канал, регулирующий ядерно-цитоплазматический транспорт различных типов РНК, мембранных белков и растворимых белков. Крупные молекулы через ЯПК транспортируются специальными белками кариоферинами
") Схематическое изображение ядерного порового комплекса (Сорокин и др. , 2007)
Схематическое изображение ядерного порового комплекса (Сорокин и др. , 2007)
 Ядерная оболочка ооцита of Xenopus laevis. Изображение получено с помощью Поровые комплексы в ядерной атомного силового микроскопа. оболочке дрожжевой клетки Область 600 x 600 нм. Толщина изображения (z-axis) – 15 нм. Желтые структуры – ядерные поры. Синий цвет – билипидный слой мембраны.
Ядерная оболочка ооцита of Xenopus laevis. Изображение получено с помощью Поровые комплексы в ядерной атомного силового микроскопа. оболочке дрожжевой клетки Область 600 x 600 нм. Толщина изображения (z-axis) – 15 нм. Желтые структуры – ядерные поры. Синий цвет – билипидный слой мембраны.
 Сканирующий электронный микроскоп. Ядро ооцита Xenopus 1 scale bar – 200 nm. 2 scale bar – 50 nm
Сканирующий электронный микроскоп. Ядро ооцита Xenopus 1 scale bar – 200 nm. 2 scale bar – 50 nm
 САМОСТОЯТЕЛЬНО Ядерные транскрипты и их транспорт
САМОСТОЯТЕЛЬНО Ядерные транскрипты и их транспорт
 Схема строения клеточного ядра 1 — ядерная оболочка (внутренняя и внешняя мембраны и перинуклеарное пространство); 2 — ядерная пора; 3 — конденсированный хроматин; 4 — диффузный хроматин; 5 — ядрышко (гранулярный и фибриллярный компоненты); 6 — интерхроматиновые гранулы (РНП); 7 — перихроматиновые гранулы (РНП); 8 — перихроматиновые фибриллы (РНП); 9 — кариоплазма.
Схема строения клеточного ядра 1 — ядерная оболочка (внутренняя и внешняя мембраны и перинуклеарное пространство); 2 — ядерная пора; 3 — конденсированный хроматин; 4 — диффузный хроматин; 5 — ядрышко (гранулярный и фибриллярный компоненты); 6 — интерхроматиновые гранулы (РНП); 7 — перихроматиновые гранулы (РНП); 8 — перихроматиновые фибриллы (РНП); 9 — кариоплазма.
 Ядерный аппарат эукариотических клеток имеет ряд отличий от прокариотических • У эукариот ДНК-содержащий компонент отделен от цитоплазмы специальной оболочкой. • В ядрах не происходит синтеза белков, т. е. в эукариотических клетках процессы синтеза ДНК и РНК разобщены от процесса синтеза белков. • Количество ДНК в ядрах эукариот в тысячи раз больше, чем в составе нуклеоидов бактерий. • В состав ядер эукариот входят несколько физически не связанных хромосом, каждая из которых содержит одну линейную молекулу ДНК. • Эукариотические хромосомы состоят сложного нуклеопротеидного комплекса – хроматина. Хроматин эукариот может пребывать в двух альтернативных состояниях: деконденсированном в интерфазе и в максимально уплотненном во время митоза (в составе митотических хромосом).
Ядерный аппарат эукариотических клеток имеет ряд отличий от прокариотических • У эукариот ДНК-содержащий компонент отделен от цитоплазмы специальной оболочкой. • В ядрах не происходит синтеза белков, т. е. в эукариотических клетках процессы синтеза ДНК и РНК разобщены от процесса синтеза белков. • Количество ДНК в ядрах эукариот в тысячи раз больше, чем в составе нуклеоидов бактерий. • В состав ядер эукариот входят несколько физически не связанных хромосом, каждая из которых содержит одну линейную молекулу ДНК. • Эукариотические хромосомы состоят сложного нуклеопротеидного комплекса – хроматина. Хроматин эукариот может пребывать в двух альтернативных состояниях: деконденсированном в интерфазе и в максимально уплотненном во время митоза (в составе митотических хромосом).