

лекция ИММУНОЛОГИЯ.ppt
- Количество слайдов: 34
 В-лимфоциты Характеристика Дифференцировка
В-лимфоциты Характеристика Дифференцировка
 зависит от функционирования В-системы специфической иммунной") Гуморальный иммунный ответ в виде продукции антител (иммуноглобулинов) зависит от функционирования В-системы специфической иммунной защиты. Центральным органом системы является костный мозг – основное место генерации В-клеток. Клеточный состав системы представлен В-лимфоцитами различной степени зрелости, вплоть до заключительной клеточной формы в гистогенезе этих клеток – плазмоцита, активно синтезирующего и секретирующего специфические иммуноглобулины
Гуморальный иммунный ответ в виде продукции антител (иммуноглобулинов) зависит от функционирования В-системы специфической иммунной защиты. Центральным органом системы является костный мозг – основное место генерации В-клеток. Клеточный состав системы представлен В-лимфоцитами различной степени зрелости, вплоть до заключительной клеточной формы в гистогенезе этих клеток – плазмоцита, активно синтезирующего и секретирующего специфические иммуноглобулины
 В процессе развития В-лимфоцитов в костном мозге происходит реорганизация генов для синтеза тяжелых и легких цепей иммуноглобулинов, в результате которой каждая клетка синтезирует только одну по специфичности тяжелую и одну легкую цепи из множества возможных. Образующийся из такой клетки клон несет иммуноглобулиновый рецептор, способный реагировать только на один антигенный эпитоп. При этом клоны, экспрессирующие иммуноглобулиновые рецепторы к своим собственным антигенам, уничтожаются и не принимают участия в дальнейшем становлении системы. Создание толерантности к собственным антигенам осуществляется на территории костного мозга. В результате на периферии оказываются клетки, рецепторы которых реагируют только на чужеродные антигены.
В процессе развития В-лимфоцитов в костном мозге происходит реорганизация генов для синтеза тяжелых и легких цепей иммуноглобулинов, в результате которой каждая клетка синтезирует только одну по специфичности тяжелую и одну легкую цепи из множества возможных. Образующийся из такой клетки клон несет иммуноглобулиновый рецептор, способный реагировать только на один антигенный эпитоп. При этом клоны, экспрессирующие иммуноглобулиновые рецепторы к своим собственным антигенам, уничтожаются и не принимают участия в дальнейшем становлении системы. Создание толерантности к собственным антигенам осуществляется на территории костного мозга. В результате на периферии оказываются клетки, рецепторы которых реагируют только на чужеродные антигены.
,") Дифференцировка В-лимфоцитов характеризуется последовательными событиями, заключающимися – - формированием специфических рецепторов – иммуноглобулинов (Ig), - избирательной экспрессией мембранных и цитоплазматических маркеров. Она осуществляется в костном мозге, где Влимфоциты проходят этап антигеннезависимой дифференцировки
Дифференцировка В-лимфоцитов характеризуется последовательными событиями, заключающимися – - формированием специфических рецепторов – иммуноглобулинов (Ig), - избирательной экспрессией мембранных и цитоплазматических маркеров. Она осуществляется в костном мозге, где Влимфоциты проходят этап антигеннезависимой дифференцировки
 Самый ранний распознаваемый этап дифференцировки В-лимфоцитов носит название про-В-лимфоцита. На этой стадии происходит начальный этап перестройки генов тяжелых µ-цепей иммуноглобулинов и появляется на мембране CD 19, который является общим маркером для всех Влимфоцитов и участвует в процессах активации клеток. Появление СD 19 происходит в клетках, экспрессирующих молекулы HLA-DR, часто в сочетании с CD 38, CD 34 и Td. T
Самый ранний распознаваемый этап дифференцировки В-лимфоцитов носит название про-В-лимфоцита. На этой стадии происходит начальный этап перестройки генов тяжелых µ-цепей иммуноглобулинов и появляется на мембране CD 19, который является общим маркером для всех Влимфоцитов и участвует в процессах активации клеток. Появление СD 19 происходит в клетках, экспрессирующих молекулы HLA-DR, часто в сочетании с CD 38, CD 34 и Td. T
 и") Отличительной чертой следующего этапа дифференцировки В-лимфоцитов является появление на мембране CD 10 (CALLA) и цитоплазматического CD 22 антигенов. Эта стадия носит название пре-Влимфоцита
Отличительной чертой следующего этапа дифференцировки В-лимфоцитов является появление на мембране CD 10 (CALLA) и цитоплазматического CD 22 антигенов. Эта стадия носит название пре-Влимфоцита
 • На следующем этапе происходит появление цитоплазматических µ-цепей иммуноглобулинов и молекулы CD 20: клетки приобретают иммунофенотип пре-Влимфоцитов. • На этой стадии происходит перестройка генов легких цепей, которая завершает процесс генетических преобразований в Влимфоцитах. Легкие цепи иммуноглобулинов ещё не синтезируются в пре-В-клетках. Клетки с непродуктивно перестроенными генами иммуноглобулинов гибнут в костном мозге
• На следующем этапе происходит появление цитоплазматических µ-цепей иммуноглобулинов и молекулы CD 20: клетки приобретают иммунофенотип пре-Влимфоцитов. • На этой стадии происходит перестройка генов легких цепей, которая завершает процесс генетических преобразований в Влимфоцитах. Легкие цепи иммуноглобулинов ещё не синтезируются в пре-В-клетках. Клетки с непродуктивно перестроенными генами иммуноглобулинов гибнут в костном мозге
 Следствием реаранжировки генов легких цепей является экспрессия полноценного мембранного Ig. M в сочетании с другими мембранными маркерами. Этот этап соответствует стадии незрелой В-клетки. Процесс антигеннезависимой дифференцировки В-лимфоцитов завершается экспрессией Ig. D, который сосуществует с Ig. Mрецептором. Присутствие на мембране Ig. M+Ig. D, CD 19, CD 20 антигенов позволяет считать В-лимфоцит зрелой (наивной) клеткой. С момента завершения формирования рецепторного комплекса В-клетка приобретает способность взаимодействовать с антигеном.
Следствием реаранжировки генов легких цепей является экспрессия полноценного мембранного Ig. M в сочетании с другими мембранными маркерами. Этот этап соответствует стадии незрелой В-клетки. Процесс антигеннезависимой дифференцировки В-лимфоцитов завершается экспрессией Ig. D, который сосуществует с Ig. Mрецептором. Присутствие на мембране Ig. M+Ig. D, CD 19, CD 20 антигенов позволяет считать В-лимфоцит зрелой (наивной) клеткой. С момента завершения формирования рецепторного комплекса В-клетка приобретает способность взаимодействовать с антигеном.
 Для завершения дифференцировки В-клеток на последнем этапе их развития в костном мозге, то есть для трансформации незрелых В-клеток с поверхностным иммуноглобулином М в зрелые В-клетки периферии экспрессирующие два иммуноглобулина М и D, требуется несколько дней. За это время происходит одно из главных событий в становлении В-системы иммунитета – отбор клеток, реагирующих только на чужеродные антигены. Вклетки, иммуноглобулиновые рецепторы которых способны взаимодействовать с собственными антигенами, либо погибают в результате апоптоза, либо приходят в состояние ареактивности (анергии). Анергия – функциональная блокада В-клеток. Апоптоз развивается обычно в тех случаях, когда распознавание антигена как «своего» происходит на поверхности клетки. Распознавание свободного (гуморального) антигена приводит к анергии
Для завершения дифференцировки В-клеток на последнем этапе их развития в костном мозге, то есть для трансформации незрелых В-клеток с поверхностным иммуноглобулином М в зрелые В-клетки периферии экспрессирующие два иммуноглобулина М и D, требуется несколько дней. За это время происходит одно из главных событий в становлении В-системы иммунитета – отбор клеток, реагирующих только на чужеродные антигены. Вклетки, иммуноглобулиновые рецепторы которых способны взаимодействовать с собственными антигенами, либо погибают в результате апоптоза, либо приходят в состояние ареактивности (анергии). Анергия – функциональная блокада В-клеток. Апоптоз развивается обычно в тех случаях, когда распознавание антигена как «своего» происходит на поверхности клетки. Распознавание свободного (гуморального) антигена приводит к анергии
 В-клетки покидают костный мозг, имея сформированный иммуноглобулиновый рецептор. Активационные антигены") • Зрелые (наивные) В-клетки покидают костный мозг, имея сформированный иммуноглобулиновый рецептор. Активационные антигены CD 23 и CD 5 на мембране отсутствуют. Содержание В-лимфоцитов в трепанобиоптатах костного мозга составляет у взрослых лиц 16 -40%, количество клеток с иммунофенотипом CD 20+CD 19+ 12 -22%. • С увеличением возраста происходит снижение общего количества В-лимфоцитов в костном мозге и их пролиферативного потенциала. • Большую роль в развитии В-лимфоцитов играет костномозговое микроокружение – клетки стромы и молекулы межклеточной адгезии, гуморальные факторы (ИЛ-7, ИЛ-3, ИЛ-1, ИЛ-4, γ-интерферон
• Зрелые (наивные) В-клетки покидают костный мозг, имея сформированный иммуноглобулиновый рецептор. Активационные антигены CD 23 и CD 5 на мембране отсутствуют. Содержание В-лимфоцитов в трепанобиоптатах костного мозга составляет у взрослых лиц 16 -40%, количество клеток с иммунофенотипом CD 20+CD 19+ 12 -22%. • С увеличением возраста происходит снижение общего количества В-лимфоцитов в костном мозге и их пролиферативного потенциала. • Большую роль в развитии В-лимфоцитов играет костномозговое микроокружение – клетки стромы и молекулы межклеточной адгезии, гуморальные факторы (ИЛ-7, ИЛ-3, ИЛ-1, ИЛ-4, γ-интерферон
 Зрелые В-клетки покидают костный мозг, попадают в кровь и заносятся в периферические лимфоидные органы, где при встречи с антигеном они проходят этап антигензависимой дифференцировки. Локализация клеток в наружных слоях коры лимфатических узлов, краевой зоне и фолликулах белой пульпы селезенки. Продолжительность жизни большинства зрелых Влимфоцитов в отсутствии антигенной стимуляции составляет несколько месяцев. Основным источником обновления популяции В-лимфоцитов служит костный мозг. Зрелые В-лимфоциты располагают необходимыми мембранными молекулами, чтобы не только распознать антиген, но и эффективно контактировать с другими клетками иммунной системы, молекулами иммуноглобулинов, компонентами комплемента, цитокинами.
Зрелые В-клетки покидают костный мозг, попадают в кровь и заносятся в периферические лимфоидные органы, где при встречи с антигеном они проходят этап антигензависимой дифференцировки. Локализация клеток в наружных слоях коры лимфатических узлов, краевой зоне и фолликулах белой пульпы селезенки. Продолжительность жизни большинства зрелых Влимфоцитов в отсутствии антигенной стимуляции составляет несколько месяцев. Основным источником обновления популяции В-лимфоцитов служит костный мозг. Зрелые В-лимфоциты располагают необходимыми мембранными молекулами, чтобы не только распознать антиген, но и эффективно контактировать с другими клетками иммунной системы, молекулами иммуноглобулинов, компонентами комплемента, цитокинами.
 В лимфатических узлах зрелые В-лимфоциты поступают в первичные фолликулы, которые представлены в виде компактных шаровых образований, не имеющих светлого (зародышевого) центра. Вторичные фолликулы отличаются от первичных наличием зародышевого центра.
В лимфатических узлах зрелые В-лимфоциты поступают в первичные фолликулы, которые представлены в виде компактных шаровых образований, не имеющих светлого (зародышевого) центра. Вторичные фолликулы отличаются от первичных наличием зародышевого центра.
 Морфология клеток первичного фолликула соответствует малому лимфоциту, большинство из них не имеет признаков активации. Лимфоциты экспрессируют на мембране CD 19, CD 20, CD 21, CD 24, CD 37, т. е. имеют фенотип периферических В-лимфоцитов. Чаще всего эти клетки содержат мембранные иммуноглобулина М и D. Активационные антигены CD 23, CD 5, CD 38, а также CD 10, как правила, отсутствуют
Морфология клеток первичного фолликула соответствует малому лимфоциту, большинство из них не имеет признаков активации. Лимфоциты экспрессируют на мембране CD 19, CD 20, CD 21, CD 24, CD 37, т. е. имеют фенотип периферических В-лимфоцитов. Чаще всего эти клетки содержат мембранные иммуноглобулина М и D. Активационные антигены CD 23, CD 5, CD 38, а также CD 10, как правила, отсутствуют
 В отсутствии антигенной стимуляции клетки через несколько дней гибнут путем апоптоза. При взаимодействии с антигеном происходит активация и пролиферация В-лимфоцитов, отражением которой является появление на мембране молекулы CD 23, повышенная экспрессия HLA-DR, утрата мембранного Ig. D.
В отсутствии антигенной стимуляции клетки через несколько дней гибнут путем апоптоза. При взаимодействии с антигеном происходит активация и пролиферация В-лимфоцитов, отражением которой является появление на мембране молекулы CD 23, повышенная экспрессия HLA-DR, утрата мембранного Ig. D.
 экзогенные антигены, ведет") Особая группа антигенов, включающая главным образом аутологичные и немногочисленные (преимущественно тимуснезависимые) экзогенные антигены, ведет к появлению экспрессии CD 5 в процессе активации В-лимфоцитов. Активированные CD 5, CD 23 В-лимфоциты мигрируют в фолликул, структура которого из-за быстрой пролиферации видоизменяется – появляется зародышевый центр и так называемая мантийная зона.
Особая группа антигенов, включающая главным образом аутологичные и немногочисленные (преимущественно тимуснезависимые) экзогенные антигены, ведет к появлению экспрессии CD 5 в процессе активации В-лимфоцитов. Активированные CD 5, CD 23 В-лимфоциты мигрируют в фолликул, структура которого из-за быстрой пролиферации видоизменяется – появляется зародышевый центр и так называемая мантийная зона.
 • В условиях микроокружения зародышевого центра происходит многоступенчатый процесс антигензависимого созревания и дифференцировки В-клеток. В темной зоне зародышевого центра активированные Вклетки утрачивают CD 23 и превращаются в центробласты, которые активно пролиферируют. • Для центробластов характерна экспрессия CD 77, CD 10, CD 19, CD 20, CD 38, практически полное отсутствие Ig. M и Ig. D. Большинство этих клеток гибнет путем апоптоза. Разрушенные погибшие клетки поглощаются макрофагами зародышевых центров
• В условиях микроокружения зародышевого центра происходит многоступенчатый процесс антигензависимого созревания и дифференцировки В-клеток. В темной зоне зародышевого центра активированные Вклетки утрачивают CD 23 и превращаются в центробласты, которые активно пролиферируют. • Для центробластов характерна экспрессия CD 77, CD 10, CD 19, CD 20, CD 38, практически полное отсутствие Ig. M и Ig. D. Большинство этих клеток гибнет путем апоптоза. Разрушенные погибшие клетки поглощаются макрофагами зародышевых центров
 Здесь же в зародышевом центре часть центробластов дифференцируются в центроциты – клетки с расщепленными ядрами. На центроцитах вновь появляются мембранные иммуноглобулины Ig. G, Ig. A или Ig. E. Синтез иммуноглобулинов с продукции Ig. M переключается на Ig. G, Ig. A и Ig. E. Переключение происходит преимущественно с синтеза Ig. M на Ig. G, обнаружение которого свидетельствует о том, что клетка подверглась антигенной стимуляции.
Здесь же в зародышевом центре часть центробластов дифференцируются в центроциты – клетки с расщепленными ядрами. На центроцитах вновь появляются мембранные иммуноглобулины Ig. G, Ig. A или Ig. E. Синтез иммуноглобулинов с продукции Ig. M переключается на Ig. G, Ig. A и Ig. E. Переключение происходит преимущественно с синтеза Ig. M на Ig. G, обнаружение которого свидетельствует о том, что клетка подверглась антигенной стимуляции.
 Одновременно с пролиферацией дифференцировкой центробластов в центроциты происходит соматические гипермутации в вариабельных участков генов иммуноглобулинов. В процессе соматических мутаций в генах иммуноглобулинов некоторых клеток возникают различные поломки, в результате чего клетка не может продуцировать иммуноглобулины и подвергается апоптозу. Из центроцитов формируются В-клетки памяти и плазматические клетки. Это происходит в апикальной светлой зоне зародышевого центра фолликула, содержащей густую сеть CD 23+ фолликулярных дендритных клеток. Они содержат значительное количество антигена, к которому формируется иммунный ответ в данном зародышевом центре. Антиген на фолликулярных дендритных клетках презентируется В-лимфоциту
Одновременно с пролиферацией дифференцировкой центробластов в центроциты происходит соматические гипермутации в вариабельных участков генов иммуноглобулинов. В процессе соматических мутаций в генах иммуноглобулинов некоторых клеток возникают различные поломки, в результате чего клетка не может продуцировать иммуноглобулины и подвергается апоптозу. Из центроцитов формируются В-клетки памяти и плазматические клетки. Это происходит в апикальной светлой зоне зародышевого центра фолликула, содержащей густую сеть CD 23+ фолликулярных дендритных клеток. Они содержат значительное количество антигена, к которому формируется иммунный ответ в данном зародышевом центре. Антиген на фолликулярных дендритных клетках презентируется В-лимфоциту
 Направленность дифференцировки В-лимфоцитов – в клетки памяти или плазматические клетки регулируется в апикальной светлой зоне зародышевых центров. Связывание молекулы CD 40 на В-клетках с соответствующим лигандом Тлимфоцитов ведет к формированию В-клеток памяти. Плазмоцитарная дифференцировка Влимфоцитов происходит после их взаимодействия с антигеном CD 23. Зрелые плазматические клетки выполняют основную функцию – синтез и секрецию иммуноглобулинов, обеспечивающих гуморальную защиту организма. При этом плазматические клетки теряют большинство В-клеточных рецепторов, на их поверхности определяется только CD 38.
Направленность дифференцировки В-лимфоцитов – в клетки памяти или плазматические клетки регулируется в апикальной светлой зоне зародышевых центров. Связывание молекулы CD 40 на В-клетках с соответствующим лигандом Тлимфоцитов ведет к формированию В-клеток памяти. Плазмоцитарная дифференцировка Влимфоцитов происходит после их взаимодействия с антигеном CD 23. Зрелые плазматические клетки выполняют основную функцию – синтез и секрецию иммуноглобулинов, обеспечивающих гуморальную защиту организма. При этом плазматические клетки теряют большинство В-клеточных рецепторов, на их поверхности определяется только CD 38.
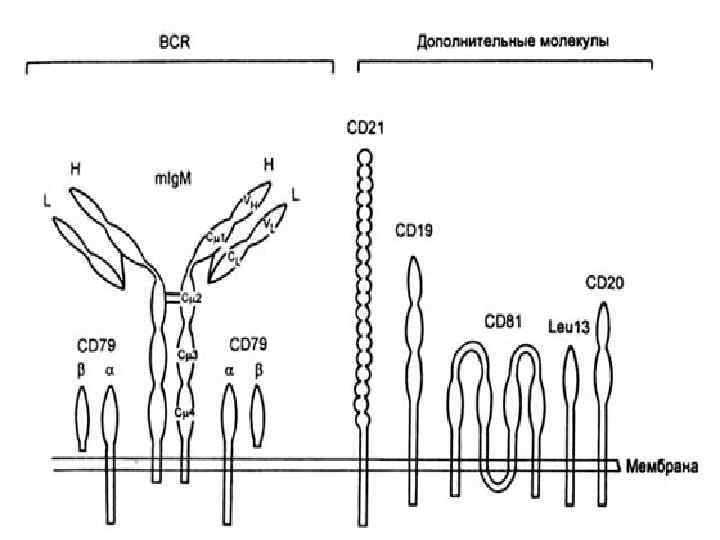
 Рецепторный комплекс В-клеток
Рецепторный комплекс В-клеток
 Большинство В-клеток периферической крови человека экспрессируют на своей поверхности иммуноглобулины двух изотипов М и G. На каждой отдельной В-клетке антигенсвязывающие центры у этих изотипов идентичны. Менее 10% В-клеток циркулирующей крови экспрессируют Ig. D, Ig. A, Ig. E, но в определенных областях организма такие клетки встречаются с большей частотой, например, В-клеток, несущих иммуноглобулин А, много в слизистой оболочке кишечника. Ассоциируя с другими молекулами на поверхности В-клеток, иммуноглобулин образует антигенраспознающий рецепторный комплекс В-клетки. К этим другим, «вспомогательным» молекулам относятся соединенные дисульфидными связями гетеродимеры, состоящие из Igα (CD 79 a) и Igβ (CD 79 b). Эти гетеродимеры, взаимодействуя (подобно составным частям Тк. Р – CD 3 комплекса Т-клеток) с трансмембранными сегментами иммуноглобулинового рецептора, участвуют в процессе активации В-клеток.
Большинство В-клеток периферической крови человека экспрессируют на своей поверхности иммуноглобулины двух изотипов М и G. На каждой отдельной В-клетке антигенсвязывающие центры у этих изотипов идентичны. Менее 10% В-клеток циркулирующей крови экспрессируют Ig. D, Ig. A, Ig. E, но в определенных областях организма такие клетки встречаются с большей частотой, например, В-клеток, несущих иммуноглобулин А, много в слизистой оболочке кишечника. Ассоциируя с другими молекулами на поверхности В-клеток, иммуноглобулин образует антигенраспознающий рецепторный комплекс В-клетки. К этим другим, «вспомогательным» молекулам относятся соединенные дисульфидными связями гетеродимеры, состоящие из Igα (CD 79 a) и Igβ (CD 79 b). Эти гетеродимеры, взаимодействуя (подобно составным частям Тк. Р – CD 3 комплекса Т-клеток) с трансмембранными сегментами иммуноглобулинового рецептора, участвуют в процессе активации В-клеток.
 Таким образом основной функцией Влимфоцитов является реализация гуморального иммунного ответа. Основой его служит активация В-клеток и их дифференцировка в антителообразующие плазматические клетки. В процессе ответа происходит переключение синтеза антител c Ig. M на Ig. Gантитела, а при иммунном ответе в слизистых оболочках - на Ig. A-антитела. Антитела нейтрализуют свободные антигены, образуя иммунные комплексы, опсанизируют клетки-мишени фагоцитов и естественных киллеров, активируют комплемент
Таким образом основной функцией Влимфоцитов является реализация гуморального иммунного ответа. Основой его служит активация В-клеток и их дифференцировка в антителообразующие плазматические клетки. В процессе ответа происходит переключение синтеза антител c Ig. M на Ig. Gантитела, а при иммунном ответе в слизистых оболочках - на Ig. A-антитела. Антитела нейтрализуют свободные антигены, образуя иммунные комплексы, опсанизируют клетки-мишени фагоцитов и естественных киллеров, активируют комплемент
 Идентификационные и функциональные маркеры В-лимфоцитов
Идентификационные и функциональные маркеры В-лимфоцитов
 • CD 19 является ключевой молекулой трансдукции сигналов, регулирующих развитие, активацию и дифференцировку Влимфоцитов. Поскольку CD 19 -рецептор экспрессирован практически на всех клетках В-ряда (начиная с ранних перегруппировок иммуноглобулиновых генов и кончая образованием В-зрелых лимфоцитов) и не обнаруживается на лимфоидных клетках других типов, то данный маркер рекомендуется для количественной характеристики общей популяции В-лимфоцитов. • Молекулы CD 20, представлены практически на всех клетках В-онтогенетического ряда (начиная от про-В-лимфоцитов до плазматических клеток). Присоединение антител к CD 20 угнетает митогенную пролиферацию В-лимфоцитов и Вклеточную дифференцировку. Молекулы CD 20, представляющие собой субстрат для протеинкиназы С, являются своеобразным рецептором, способствующим включению цепи по проведению специфического сигнала (при стимуляции антигеном) с поверхности внутрь клетки к ядру.
• CD 19 является ключевой молекулой трансдукции сигналов, регулирующих развитие, активацию и дифференцировку Влимфоцитов. Поскольку CD 19 -рецептор экспрессирован практически на всех клетках В-ряда (начиная с ранних перегруппировок иммуноглобулиновых генов и кончая образованием В-зрелых лимфоцитов) и не обнаруживается на лимфоидных клетках других типов, то данный маркер рекомендуется для количественной характеристики общей популяции В-лимфоцитов. • Молекулы CD 20, представлены практически на всех клетках В-онтогенетического ряда (начиная от про-В-лимфоцитов до плазматических клеток). Присоединение антител к CD 20 угнетает митогенную пролиферацию В-лимфоцитов и Вклеточную дифференцировку. Молекулы CD 20, представляющие собой субстрат для протеинкиназы С, являются своеобразным рецептором, способствующим включению цепи по проведению специфического сигнала (при стимуляции антигеном) с поверхности внутрь клетки к ядру.
 • СD 21 – рецептор для компонента комплемента С 3 b. Эта молекула является также рецептором для вируса Эпштейна -Барра. Она экспрессирована на всех зрелых В-клетках и фолликулярных дендритных клетках. • СD 22 – рецептор иммуноглобулиновой природы. В своем цитоплазматическом домене содержит иммунорецепторный ингибитор. Участвует в негативной регуляции В-клеток. Мутации этого рецептора вызывают гиперпролиферацию Влимфоцитов. • CD 23 – низкоаффинный рецептор иммуноглобулина Е. В слабой степени экспрессирован на зрелых В-лимфоцитах. Высокий уровень характерен для фолликулярных дендритных клеток и особенно для трансформированных вирусом Эпштейна-Барра В-лимфобластов. Для В-клеток CD 23 является маркером дифференцировки, который утрачивается иммуноглобулинсекретирующими клетками
• СD 21 – рецептор для компонента комплемента С 3 b. Эта молекула является также рецептором для вируса Эпштейна -Барра. Она экспрессирована на всех зрелых В-клетках и фолликулярных дендритных клетках. • СD 22 – рецептор иммуноглобулиновой природы. В своем цитоплазматическом домене содержит иммунорецепторный ингибитор. Участвует в негативной регуляции В-клеток. Мутации этого рецептора вызывают гиперпролиферацию Влимфоцитов. • CD 23 – низкоаффинный рецептор иммуноглобулина Е. В слабой степени экспрессирован на зрелых В-лимфоцитах. Высокий уровень характерен для фолликулярных дендритных клеток и особенно для трансформированных вирусом Эпштейна-Барра В-лимфобластов. Для В-клеток CD 23 является маркером дифференцировки, который утрачивается иммуноглобулинсекретирующими клетками
 Клетки иммунологической памяти
Клетки иммунологической памяти
 В результате иммунного ответа образуются Т- и В-клетки памяти, которые обеспечивают ускоренное развитие реакции на повторное попадание в организм тех же антигенов – вторичный иммунный ответ. Клетки памяти представляют собой малые лимфоциты, обладающие двумя важными признаками: - способностью к рециркуляции - и большой продолжительностью жизни, которая обуславливает длительное сохранение иммунитета к возбудителям инфекционных заболеваний и другим чужеродным агентам
В результате иммунного ответа образуются Т- и В-клетки памяти, которые обеспечивают ускоренное развитие реакции на повторное попадание в организм тех же антигенов – вторичный иммунный ответ. Клетки памяти представляют собой малые лимфоциты, обладающие двумя важными признаками: - способностью к рециркуляции - и большой продолжительностью жизни, которая обуславливает длительное сохранение иммунитета к возбудителям инфекционных заболеваний и другим чужеродным агентам
 В-клетки памяти образуются в зародышевых центрах лимфатических узлов, селезенки и лимфоидных образованиях некоторых органов, например кишечника. Презентация антигена может происходить без предварительной внутриклеточной его переработки. Для В-клеток памяти характерны наличие Ig. G или Ig. A и высокая экспрессия ингибитора апоптоза Bcl-2. В популяции Вклеток с иммуноглобулином М, клеток памяти практически нет. При поступлении в кровь В-клетки памяти рециркулируют и накапливаются в костном мозге.
В-клетки памяти образуются в зародышевых центрах лимфатических узлов, селезенки и лимфоидных образованиях некоторых органов, например кишечника. Презентация антигена может происходить без предварительной внутриклеточной его переработки. Для В-клеток памяти характерны наличие Ig. G или Ig. A и высокая экспрессия ингибитора апоптоза Bcl-2. В популяции Вклеток с иммуноглобулином М, клеток памяти практически нет. При поступлении в кровь В-клетки памяти рециркулируют и накапливаются в костном мозге.
 При контакте В- и Т-клеток памяти с антигеном возникает вторичный иммунный ответ, основные признаки которого отличные от признаков первичного иммунного ответа. Характеристика вторичного иммунного ответа • более раннее развитие иммунного ответа на повторное введение антигена. Сокращение латентного периода на 2 -4 дня. • уменьшение дозы антигена, необходимой для достижения оптимального ответа. • Увеличение силы и продолжительности иммунного ответа. • Усиление образования антител: • увеличение количества антителообразующих клеток • активация Th 2 и усиление выработки их цитокинов • образование антител преимущественно за счет иммуноглобулина А и G • повышение аффинности антител • усиление гуморального иммунитета
При контакте В- и Т-клеток памяти с антигеном возникает вторичный иммунный ответ, основные признаки которого отличные от признаков первичного иммунного ответа. Характеристика вторичного иммунного ответа • более раннее развитие иммунного ответа на повторное введение антигена. Сокращение латентного периода на 2 -4 дня. • уменьшение дозы антигена, необходимой для достижения оптимального ответа. • Увеличение силы и продолжительности иммунного ответа. • Усиление образования антител: • увеличение количества антителообразующих клеток • активация Th 2 и усиление выработки их цитокинов • образование антител преимущественно за счет иммуноглобулина А и G • повышение аффинности антител • усиление гуморального иммунитета
 Усиление клеточных реакций: • увеличение числа антигенспецифических Ткиллеров и Т-эффекторов гиперчувствительности замедленного типа • активация Th 1 и усиление выработки их цитокинов • повышение аффинности антигенспецифических рецепторов Т-клеток • усиление защитных клеточных реакций • сокращение сроков отторжения пересаженной аллогенной ткани на 3 -5 дней
Усиление клеточных реакций: • увеличение числа антигенспецифических Ткиллеров и Т-эффекторов гиперчувствительности замедленного типа • активация Th 1 и усиление выработки их цитокинов • повышение аффинности антигенспецифических рецепторов Т-клеток • усиление защитных клеточных реакций • сокращение сроков отторжения пересаженной аллогенной ткани на 3 -5 дней
 Природа Т-клеток памяти изучена в меньшей степени. Неизвестно, являются ли Т-клетки памяти долгоживущей популяцией антигенспецифических эффекторных клеток или это особая популяция клеток, обладающих высокоаффинными антигенраспознающими рецепторами
Природа Т-клеток памяти изучена в меньшей степени. Неизвестно, являются ли Т-клетки памяти долгоживущей популяцией антигенспецифических эффекторных клеток или это особая популяция клеток, обладающих высокоаффинными антигенраспознающими рецепторами
 Процент клеток, которые становятся клетками памяти, крайне незначителен. В лимфатических узлах клетки памяти остаются в течение 18 -20 часов, а затем покидают их через эфферентные лимфатические сосуды. Клетки памяти находятся на G 1 - стадии клеточного цикла, то есть они вышли из стадии покоя G 0 и готовы к быстрому превращению в эффекторные клетки при очередном контакте с антигеном. Переход клеток памяти в состояние длительной иммунологической памяти происходит из-за прекращения активации антигенспецифических рецепторов клеток. Без антигенной стимуляции клетки памяти существуют как неделящаяся субпопуляция клеток
Процент клеток, которые становятся клетками памяти, крайне незначителен. В лимфатических узлах клетки памяти остаются в течение 18 -20 часов, а затем покидают их через эфферентные лимфатические сосуды. Клетки памяти находятся на G 1 - стадии клеточного цикла, то есть они вышли из стадии покоя G 0 и готовы к быстрому превращению в эффекторные клетки при очередном контакте с антигеном. Переход клеток памяти в состояние длительной иммунологической памяти происходит из-за прекращения активации антигенспецифических рецепторов клеток. Без антигенной стимуляции клетки памяти существуют как неделящаяся субпопуляция клеток
 • Клетки памяти способны рециркулировать в кровеносной и лимфатической системах, обеспечивая эндогенный надзор и защиту организма от проникновения чужеродных веществ. Клетки памяти постоянно присутствуют в лимфоидных органах и костном мозге. • Таким образом иммунологическая память связана прежде всего с клетками памяти, которые рециркулируют во всех тканях организма, где есть кровеносные сосуды, и выполняют по существу функцию эффекторных клеток.
• Клетки памяти способны рециркулировать в кровеносной и лимфатической системах, обеспечивая эндогенный надзор и защиту организма от проникновения чужеродных веществ. Клетки памяти постоянно присутствуют в лимфоидных органах и костном мозге. • Таким образом иммунологическая память связана прежде всего с клетками памяти, которые рециркулируют во всех тканях организма, где есть кровеносные сосуды, и выполняют по существу функцию эффекторных клеток.