4Уровни упаковки хроматина.ppt
- Количество слайдов: 34
 Уровни упаковки ДНК
Уровни упаковки ДНК
, комплекс ДНК и белков. Образует хромосомы") ХРОМАТИН • (от греч. Chroma — цвет, краска), комплекс ДНК и белков. Образует хромосомы клеток эукариот, входит в состав нуклеоида у прокариот. • В. Флемминг (1880) • Состав хроматина: – ДНК (30— 45%), – гистоны (30— 50%): Н 1, H 2 A, H 2 B, H 3 и H 4 – негистоновые белки (4— 33% ) • Эухроматин и Гетерохроматин (факультативный и конститутивный)
ХРОМАТИН • (от греч. Chroma — цвет, краска), комплекс ДНК и белков. Образует хромосомы клеток эукариот, входит в состав нуклеоида у прокариот. • В. Флемминг (1880) • Состав хроматина: – ДНК (30— 45%), – гистоны (30— 50%): Н 1, H 2 A, H 2 B, H 3 и H 4 – негистоновые белки (4— 33% ) • Эухроматин и Гетерохроматин (факультативный и конститутивный)
 Структура диффузный конденсированный Синтез РНК") СВОЙСТВА ГЕТЕРОХРОМАТИН КОНСТИТУТИВНЫЙ ЭУХРОМАТИН АКТИВНЫЙ НЕАКТИВНЫЙ ГЕТЕРОХРОМАТИН (ФАКУЛЬТАТИВНЫЙ ) Структура диффузный конденсированный Синтез РНК + - Синтез ДНК + + Тип нуклеотидный последовательностей ДНК Уникальные, умеренные повторы Высокоповторяющ аяся, сателлитная ДНК Локализация Плечи хромосом Центромера, теломеры, интеркалярный - + поздняя репликация
СВОЙСТВА ГЕТЕРОХРОМАТИН КОНСТИТУТИВНЫЙ ЭУХРОМАТИН АКТИВНЫЙ НЕАКТИВНЫЙ ГЕТЕРОХРОМАТИН (ФАКУЛЬТАТИВНЫЙ ) Структура диффузный конденсированный Синтез РНК + - Синтез ДНК + + Тип нуклеотидный последовательностей ДНК Уникальные, умеренные повторы Высокоповторяющ аяся, сателлитная ДНК Локализация Плечи хромосом Центромера, теломеры, интеркалярный - + поздняя репликация
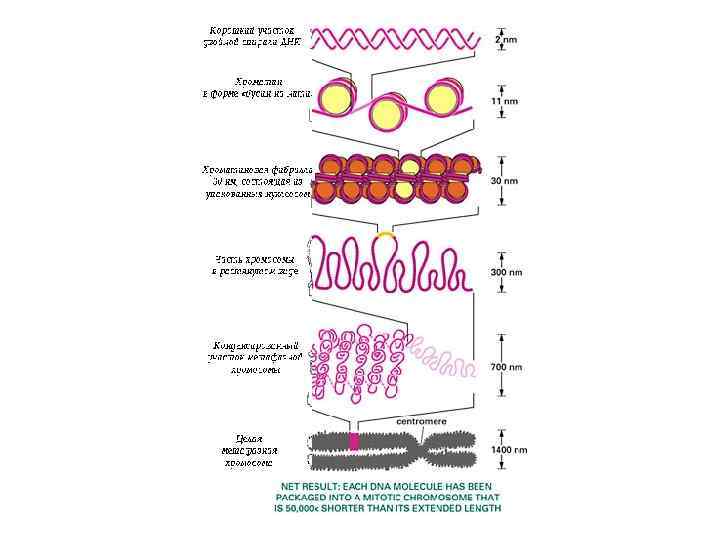
 УРОВНИ УПАКОВКИ ХРОМАТИНА • Длина ДНК в диплоидном наборе хромосом человека - 2 м, а всех метафазных хромосом - 150 мкм • ДНК соединяется с гистоновыми (основными) и негистоновыми (кислыми) белками, образуя нуклеопротеидные фибриллы • Компактизация генетического материала достигается спирализацией, (конденсацией) и связью ДНК с белками
УРОВНИ УПАКОВКИ ХРОМАТИНА • Длина ДНК в диплоидном наборе хромосом человека - 2 м, а всех метафазных хромосом - 150 мкм • ДНК соединяется с гистоновыми (основными) и негистоновыми (кислыми) белками, образуя нуклеопротеидные фибриллы • Компактизация генетического материала достигается спирализацией, (конденсацией) и связью ДНК с белками
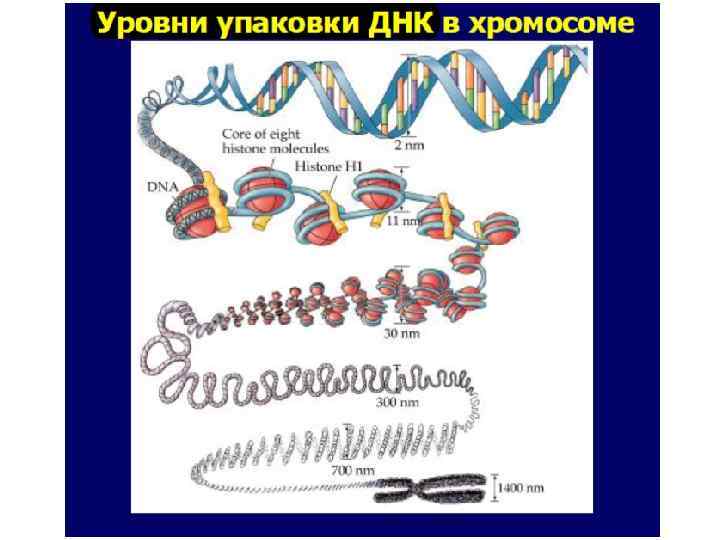
 НУКЛЕОСОМНЫЙ УРОВЕНЬ Нуклеосома - дисковидная структура, являющаяся элементарной единицей упаковки хромосомной ДНК в хроматине. Описана в 1974 году Р. Корнбергом Представляет собой белковую глобулу (октаэдр), содержащую по 2 молекулы четырех гистонов - (Н 2 А, Н 2 В, Н 3, Н 4), вокруг которой двойная спираль ДНК образует 1, 67 витка (146 + 10 -100 пар нуклеотидов). Диаметр нуклеосомной нити - около 11 нм Длина ДНК уменьшается в 5 -7 раз.
НУКЛЕОСОМНЫЙ УРОВЕНЬ Нуклеосома - дисковидная структура, являющаяся элементарной единицей упаковки хромосомной ДНК в хроматине. Описана в 1974 году Р. Корнбергом Представляет собой белковую глобулу (октаэдр), содержащую по 2 молекулы четырех гистонов - (Н 2 А, Н 2 В, Н 3, Н 4), вокруг которой двойная спираль ДНК образует 1, 67 витка (146 + 10 -100 пар нуклеотидов). Диаметр нуклеосомной нити - около 11 нм Длина ДНК уменьшается в 5 -7 раз.
 Н 1 (очень богатая лизином) 29% 1% 23000") Фракция Лизин Аргинин Мол. вес (Да) Н 1 (очень богатая лизином) 29% 1% 23000 Н 2 В (умеренно богатая лиз) 16% 6% 13774 Н 2 А (умеренно богатая лиз и арг) 11% 9% 13960 Н 4 (богатая арг и гли) 11% 14% 11282 Н 3 (очень богатая арг); есть цис, а в других – нет 10% 13% 15348
Фракция Лизин Аргинин Мол. вес (Да) Н 1 (очень богатая лизином) 29% 1% 23000 Н 2 В (умеренно богатая лиз) 16% 6% 13774 Н 2 А (умеренно богатая лиз и арг) 11% 9% 13960 Н 4 (богатая арг и гли) 11% 14% 11282 Н 3 (очень богатая арг); есть цис, а в других – нет 10% 13% 15348
 СТРУКТУРА НУКЛЕОСОМЫ ДНК+ Гистоновый октамер: 2 H 2 A 2 H 2 B 2 H 3 2 H 4
СТРУКТУРА НУКЛЕОСОМЫ ДНК+ Гистоновый октамер: 2 H 2 A 2 H 2 B 2 H 3 2 H 4
") Структура нуклеосомы (рентгеноструктурный анализ)
Структура нуклеосомы (рентгеноструктурный анализ)

 НУКЛЕОНЕМНЫЙ УРОВЕНЬ, 30 -нм фибрилла Дальнейшая компактизация нуклеосомной нити обеспечивается пистоном H 1, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом. В результате образуется более компактная структура, построенная по типу соленоида. Такая Хроматиновая фибрилла, имеет диаметр 20— 30 нм. Виток спирали содержит 5 -6 нуклеосом ДНП укорачивается в 6 раз
НУКЛЕОНЕМНЫЙ УРОВЕНЬ, 30 -нм фибрилла Дальнейшая компактизация нуклеосомной нити обеспечивается пистоном H 1, который, соединяясь с линкерной ДНК и двумя соседними белковыми телами, сближает их друг с другом. В результате образуется более компактная структура, построенная по типу соленоида. Такая Хроматиновая фибрилла, имеет диаметр 20— 30 нм. Виток спирали содержит 5 -6 нуклеосом ДНП укорачивается в 6 раз
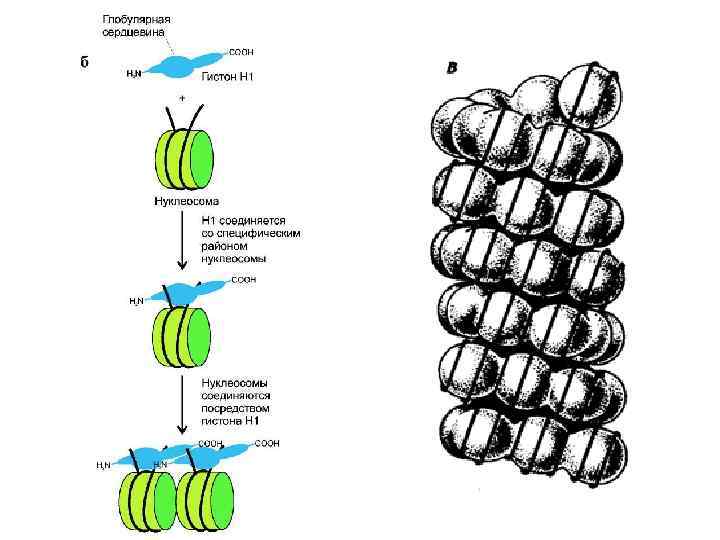
 Модели организации 30 -нм фибриллы
Модели организации 30 -нм фибриллы
 Хвосты гистонов – регуляторы активности генов и компактизации хроматина
Хвосты гистонов – регуляторы активности генов и компактизации хроматина
 Ацетилирование гистонов определяет компактность хроматиновой фибриллы
Ацетилирование гистонов определяет компактность хроматиновой фибриллы
 В период синтеза ДНК происходит ацетилирование гистонов сердцевины с последующим деацилированием. Основное фосфорилирование гистона HI происходит в начале митоза; в конце митоза гистон HI дефосфорилируется. Новые и старые гистоны ацетилированы; Н 1 может фосфорилироваться 1 -2 раза Из гистонов удаляются ацетильные группы Н 1 фосфорилирован 4 раза Н 1 дефосфорилирован
В период синтеза ДНК происходит ацетилирование гистонов сердцевины с последующим деацилированием. Основное фосфорилирование гистона HI происходит в начале митоза; в конце митоза гистон HI дефосфорилируется. Новые и старые гистоны ацетилированы; Н 1 может фосфорилироваться 1 -2 раза Из гистонов удаляются ацетильные группы Н 1 фосфорилирован 4 раза Н 1 дефосфорилирован
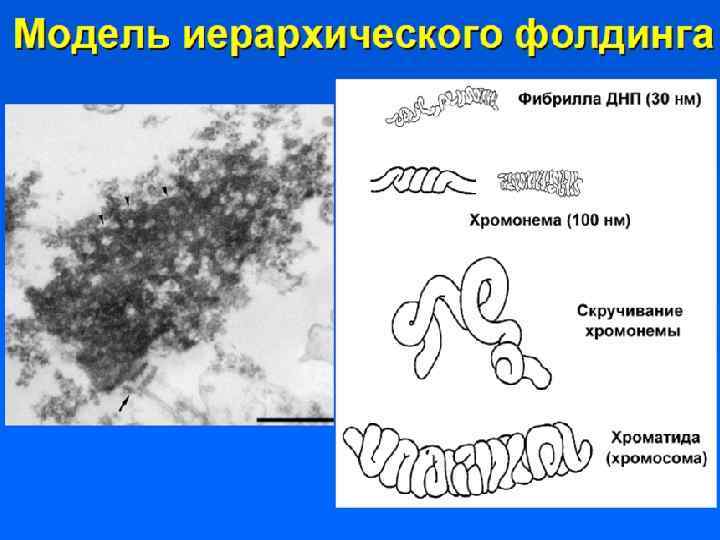
 Хромонемный уровень Следующий уровень структурной организации генетического материала обусловлен укладкой хроматиновой фибриллы в петли. В их образовании принимают участие негистоновые белки, способные узнавать специфические нуклеотидные последовательности вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы. Участок ДНК, соответствующий одной петле, содержит от 20 до 80 т. п. н. Возможно, каждая петля является функциональной единицей генома. В результате такой упаковки хроматиновая фибрилла диаметром 20 — 30 нм преобразуется в структуру диаметром 100— 200 нм, называемую интерфазной хромонемой.
Хромонемный уровень Следующий уровень структурной организации генетического материала обусловлен укладкой хроматиновой фибриллы в петли. В их образовании принимают участие негистоновые белки, способные узнавать специфические нуклеотидные последовательности вненуклеосомной ДНК, отдаленные друг от друга на расстояние в несколько тысяч пар нуклеотидов. Эти белки сближают указанные участки с образованием петель из расположенных между ними фрагментов хроматиновой фибриллы. Участок ДНК, соответствующий одной петле, содержит от 20 до 80 т. п. н. Возможно, каждая петля является функциональной единицей генома. В результате такой упаковки хроматиновая фибрилла диаметром 20 — 30 нм преобразуется в структуру диаметром 100— 200 нм, называемую интерфазной хромонемой.
 Супернуклеосомная нить образует петли и изгибы, составляя основу хроматиды. Обнаруживается в профазе. Диаметр петель = 50 нм, диаметр хроматиды 300 нм. Нить ДНП укорачивается еще в 10 -20 раз.
Супернуклеосомная нить образует петли и изгибы, составляя основу хроматиды. Обнаруживается в профазе. Диаметр петель = 50 нм, диаметр хроматиды 300 нм. Нить ДНП укорачивается еще в 10 -20 раз.
 Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации, образуя структурные блоки, объединяющие соседние петли с одинаковой организацией. Они выявляются в интерфазном ядре в виде глыбок хроматина. Существование таких структурных блоков обусловливает картину неравномерного распределения некоторых красителей в метафазных хромосомах, что используют в цитогенетических исследованиях
Отдельные участки интерфазной хромонемы подвергаются дальнейшей компактизации, образуя структурные блоки, объединяющие соседние петли с одинаковой организацией. Они выявляются в интерфазном ядре в виде глыбок хроматина. Существование таких структурных блоков обусловливает картину неравномерного распределения некоторых красителей в метафазных хромосомах, что используют в цитогенетических исследованиях
 Хромосомный уровень Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией хроматина. Отдельные хромосомы становятся хорошо различимы. Этот процесс начинается в профазе, достигая своего максимального выражения в метафазе митоза и анафазе. В телофазе митоза происходит декомпактизация вещества хромосом, которое приобретает структуру интерфазного хроматина. Митотическая суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена в анафазе митоза. Толщина плеча хромосомы – 700 нм. Укорочение ДНП еще в 20 раз.
Хромосомный уровень Вступление клетки из интерфазы в митоз сопровождается суперкомпактизацией хроматина. Отдельные хромосомы становятся хорошо различимы. Этот процесс начинается в профазе, достигая своего максимального выражения в метафазе митоза и анафазе. В телофазе митоза происходит декомпактизация вещества хромосом, которое приобретает структуру интерфазного хроматина. Митотическая суперкомпактизация облегчает распределение хромосом к полюсам митотического веретена в анафазе митоза. Толщина плеча хромосомы – 700 нм. Укорочение ДНП еще в 20 раз.
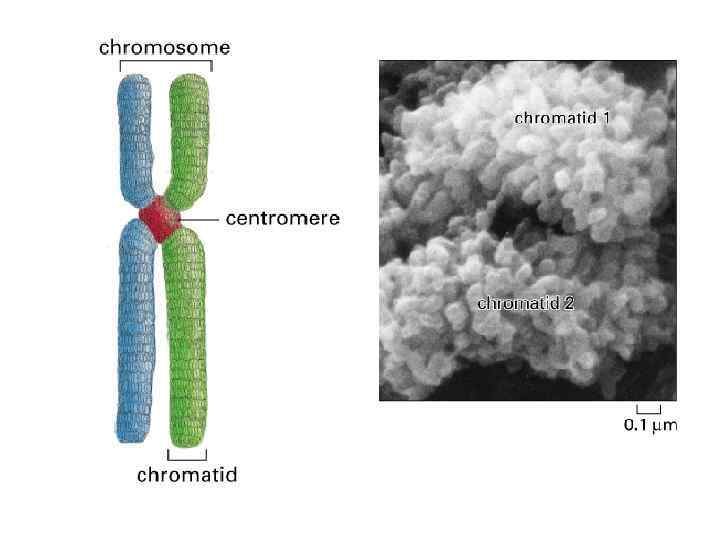
 ХРОМОСОМЫ В ИНТЕРФАЗЕ И В МЕТАФАЗЕ
ХРОМОСОМЫ В ИНТЕРФАЗЕ И В МЕТАФАЗЕ
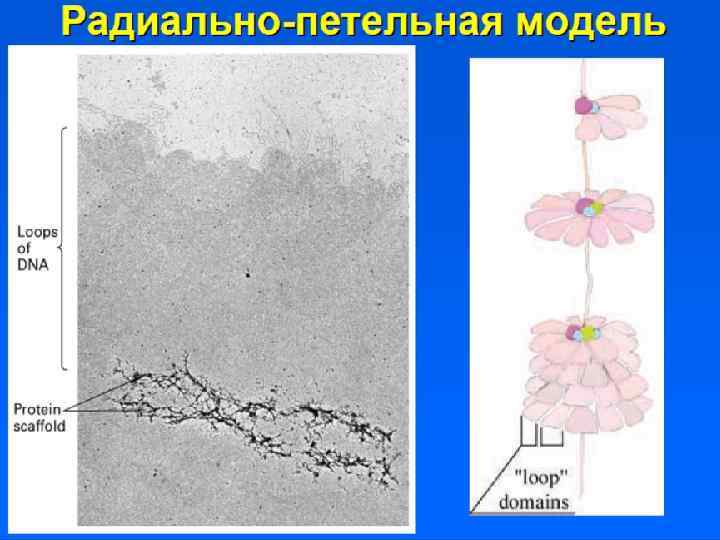

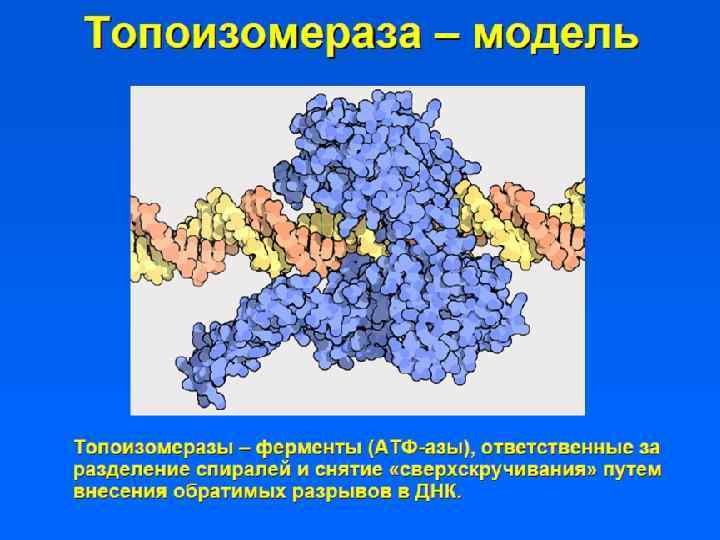
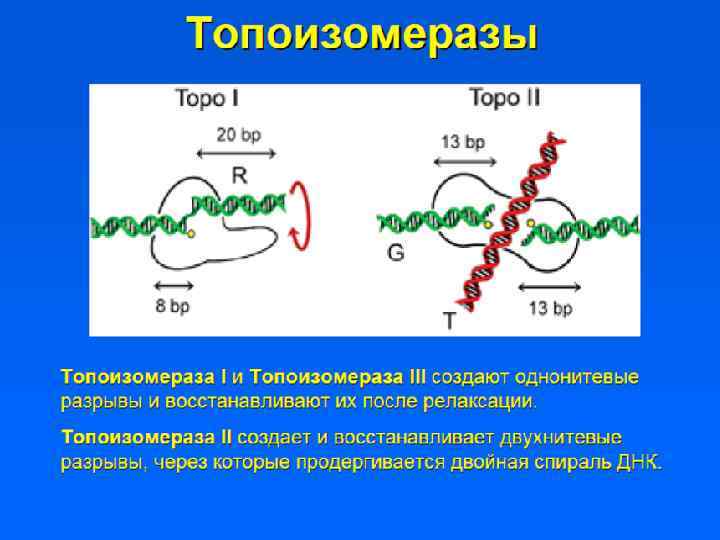
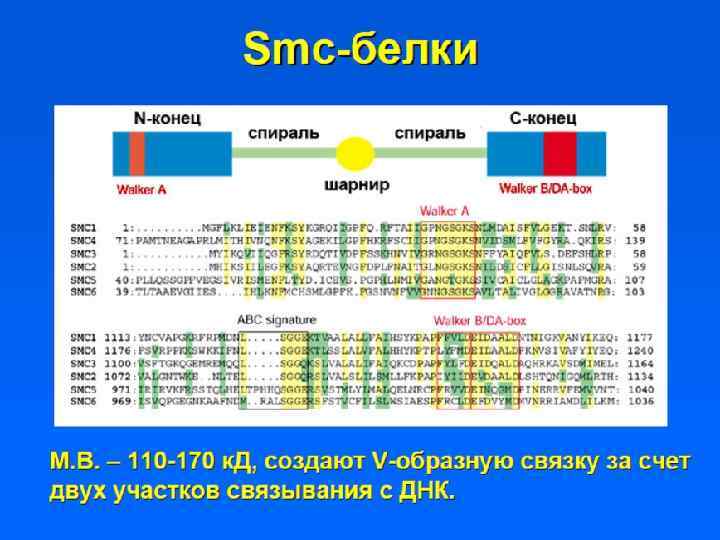
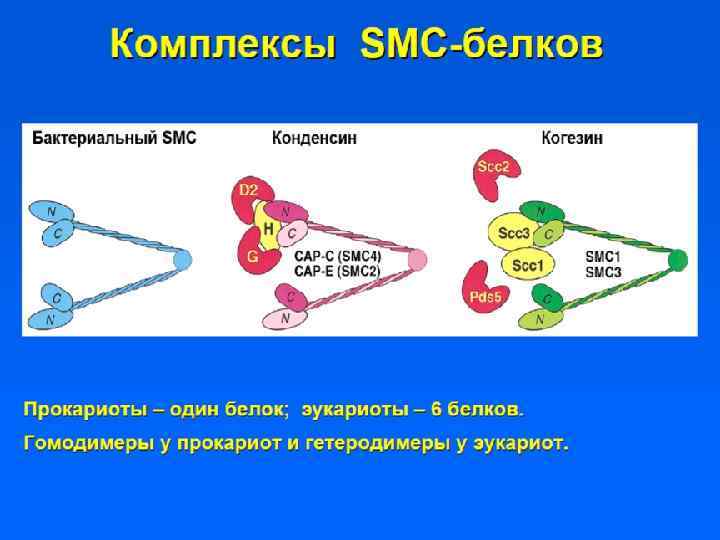
") • Участки ДНК, способные связываться с ядерным матриксом, называются MARs (Matrix assosiated regions) • Белки ядерного матрикса: – Инсуляторные белки – Ламины – ТОРО II – Белки транскрипции и репликации
• Участки ДНК, способные связываться с ядерным матриксом, называются MARs (Matrix assosiated regions) • Белки ядерного матрикса: – Инсуляторные белки – Ламины – ТОРО II – Белки транскрипции и репликации
 Инсуляторные белки – белки, связывающиеся с инсуляторами - Инсуляторы — последовательности ДНК, обладающие способностью блокировать сигналы, исходящие от окружения. 1. Блокируют взаимодействие между энхансером и промотором, если находится между ними. 2. инсулятор выполняет барьерную функцию для распространяющегося конденсированного хроматина.
Инсуляторные белки – белки, связывающиеся с инсуляторами - Инсуляторы — последовательности ДНК, обладающие способностью блокировать сигналы, исходящие от окружения. 1. Блокируют взаимодействие между энхансером и промотором, если находится между ними. 2. инсулятор выполняет барьерную функцию для распространяющегося конденсированного хроматина.