

Lektsia_3_membrana.ppt
- Количество слайдов: 77
 УО ”Международный государственный университет им. А. Д. Сахарова” Факультет экологической медицины кафедра экологической и молекулярной генетики
УО ”Международный государственный университет им. А. Д. Сахарова” Факультет экологической медицины кафедра экологической и молекулярной генетики
 n Cтарший преподаватель кафедры n Лозинская Ольга Владиславовна
n Cтарший преподаватель кафедры n Лозинская Ольга Владиславовна
") Мембранная система клетки n Плазматическая мембрана n Внутренние мембраны n Мембраны ядерной оболочки (кариолеммы) n Эндоплазматический ретикулум n Пластинчатый комплекс n Лизосомы n Пероксисомы n Везикулы и др. n Наружные мембраны митохондрий и пластид n Сопрягающие мембраны митохондрий и пластид
Мембранная система клетки n Плазматическая мембрана n Внутренние мембраны n Мембраны ядерной оболочки (кариолеммы) n Эндоплазматический ретикулум n Пластинчатый комплекс n Лизосомы n Пероксисомы n Везикулы и др. n Наружные мембраны митохондрий и пластид n Сопрягающие мембраны митохондрий и пластид
 Общая характеристика клеточных мембран n 1. Разные типы мембран различаются по своей толщине, но в большинстве случаев толщина мембран составляет 6 -10 нм; 2. Мембраны - это липопротеиновые структуры (липид 25 -60%, + белок 40 -75%. ). на внешних поверхностях присоединены углеводные компоненты 2 -10%(гликозильные группы). n 3. Липиды спонтанно образуют бислой, т. к. молекулы имеют полярные головки и неполярные хвосты. n 4. Способность к слиянию и смыканию фрагментов с образованием замкнутых пузырьков n 5. Мембранные белки выполняют разнообразные функции. n 6. Гликозильные группы связаны с механизмом распознавания. n 7. Мембрана асимметрична n 8. Неспособность возникать в клетке de novo без участия уже существующих мембранных образований.
Общая характеристика клеточных мембран n 1. Разные типы мембран различаются по своей толщине, но в большинстве случаев толщина мембран составляет 6 -10 нм; 2. Мембраны - это липопротеиновые структуры (липид 25 -60%, + белок 40 -75%. ). на внешних поверхностях присоединены углеводные компоненты 2 -10%(гликозильные группы). n 3. Липиды спонтанно образуют бислой, т. к. молекулы имеют полярные головки и неполярные хвосты. n 4. Способность к слиянию и смыканию фрагментов с образованием замкнутых пузырьков n 5. Мембранные белки выполняют разнообразные функции. n 6. Гликозильные группы связаны с механизмом распознавания. n 7. Мембрана асимметрична n 8. Неспособность возникать в клетке de novo без участия уже существующих мембранных образований.

 Модели биологических мембран n В 1935 г. модель Даниэля Доусона унитарная модель био мембран. Липидный бислой – структурная основа. Наружный и внутренний слои – глобулярные белки. Симметричная модель. n Модель Робертсона (середина 60 х г). Мембрана представляет собой 3 х слойную структуру, средний слой из липидов. Белковые молекулы развернуты на поверхности двойного липидного слоя вследствие электростатических взаимодействий заряженными головками фосфолипидов. Модель Робертсона ассимметрична, так как на наружной поверхности мембраны – гликопротеиды. n В группе моделей предполагается наличие белков матрицы. Модель Лючи (середина 60 х г. ) – белковокристаллическая модель. n Модель _ (1970) сохраняется концепция липидного бислоя, однако этот слой прирастается участками симметрично расположенных белков, они жестко фиксированны пространственно за счет дальнодействующих белок-белковых свойств. n Модель Сингера и Николсона. 60 -70 г. Основа – липидный бислой, в который включены молекулы интегральных и периферических белков. n Жидкомозаичная модель. С ее помощью объясняется проницаемость мембран.
Модели биологических мембран n В 1935 г. модель Даниэля Доусона унитарная модель био мембран. Липидный бислой – структурная основа. Наружный и внутренний слои – глобулярные белки. Симметричная модель. n Модель Робертсона (середина 60 х г). Мембрана представляет собой 3 х слойную структуру, средний слой из липидов. Белковые молекулы развернуты на поверхности двойного липидного слоя вследствие электростатических взаимодействий заряженными головками фосфолипидов. Модель Робертсона ассимметрична, так как на наружной поверхности мембраны – гликопротеиды. n В группе моделей предполагается наличие белков матрицы. Модель Лючи (середина 60 х г. ) – белковокристаллическая модель. n Модель _ (1970) сохраняется концепция липидного бислоя, однако этот слой прирастается участками симметрично расположенных белков, они жестко фиксированны пространственно за счет дальнодействующих белок-белковых свойств. n Модель Сингера и Николсона. 60 -70 г. Основа – липидный бислой, в который включены молекулы интегральных и периферических белков. n Жидкомозаичная модель. С ее помощью объясняется проницаемость мембран.
 Жидкостно-мозаичная модель биологической мембраны n Жидкостность – способность мембраны находится подвижном состоянии и выполнять поддержку для структур(липиды) n Мозаичность – погружение структур на разные уровни в жидкую поддерживающую фазу (белки)
Жидкостно-мозаичная модель биологической мембраны n Жидкостность – способность мембраны находится подвижном состоянии и выполнять поддержку для структур(липиды) n Мозаичность – погружение структур на разные уровни в жидкую поддерживающую фазу (белки)
 Биохимический состав биологических мембран Липиды мембран В составе мембран может входить более 1500 различных липидов. Лишь около 30 из них встречаются в значительных количествах. Большая часть их принадлежит к -фосфолипидам ( от 40 до 90%), - гликолипидам - нейтральным липидам, в том числе стероидам (холестерол). - Минорные липидные составляющие весьма вариабельны. В эту группу входят убихинон, токоферолы, полипрениловые спирты, каротиноиды, лизолипиды, свободные ЖК.
Биохимический состав биологических мембран Липиды мембран В составе мембран может входить более 1500 различных липидов. Лишь около 30 из них встречаются в значительных количествах. Большая часть их принадлежит к -фосфолипидам ( от 40 до 90%), - гликолипидам - нейтральным липидам, в том числе стероидам (холестерол). - Минорные липидные составляющие весьма вариабельны. В эту группу входят убихинон, токоферолы, полипрениловые спирты, каротиноиды, лизолипиды, свободные ЖК.
 n Первый элементный анализ Л. выполнен в нач. 19 в. А. Лавуазье, а первые исследования по выяснению хим. строения Л. принадлежат К. Шееле и М. Шеврёлю. Впервые синтезы триглицеридов осуществили М. Бертло в 1854 и Ш. Вюрц в 1859. Фосфолипиды выделены М. Гобли в 1847, а затем получены в более чистом виде Ф. А. Хоппе-Зейлером в 1877. К этому времени уже было установлено строение ряда важнейших жирных кислот. Дальнейшую историю изучения Л. можно разделить на три периода, различающиеся по методич. уровню исследований. На первом этапе (1880 -1950) Л. исследовали традиционными методами орг. химии, второй этап (1950 -1970) характеризуется широким применением методов хроматографии, а последний (7080 -е гг. ) - использованием таких физ. -хим. методов, как массспектрометрия, оптич. спектроскопия и радиоспектроскопия, флуоресцентный анализ и др.
n Первый элементный анализ Л. выполнен в нач. 19 в. А. Лавуазье, а первые исследования по выяснению хим. строения Л. принадлежат К. Шееле и М. Шеврёлю. Впервые синтезы триглицеридов осуществили М. Бертло в 1854 и Ш. Вюрц в 1859. Фосфолипиды выделены М. Гобли в 1847, а затем получены в более чистом виде Ф. А. Хоппе-Зейлером в 1877. К этому времени уже было установлено строение ряда важнейших жирных кислот. Дальнейшую историю изучения Л. можно разделить на три периода, различающиеся по методич. уровню исследований. На первом этапе (1880 -1950) Л. исследовали традиционными методами орг. химии, второй этап (1950 -1970) характеризуется широким применением методов хроматографии, а последний (7080 -е гг. ) - использованием таких физ. -хим. методов, как массспектрометрия, оптич. спектроскопия и радиоспектроскопия, флуоресцентный анализ и др.
 n Липидный состав клеточных мембран изменчив. В меньшей степени это проявляется в животных клетках, находящихся в условиях стабильной внутренней среды. Однако можно модифицировать состав липидов в некоторых мембранах, меняя пищевой рацион. n Липидный состав мембран растений заметно изменяется в зависимости от освещенности, температуры и р. Н. n Еще более изменчив состав бактериальных мембран. Он варьирует не только в зависимости от штамма, но и в пределах одного и того же штамма, а также от условий культивирования и фазы роста. n У вирусов, имеющих липопротеиновую оболочку, липидный состав мембран также не постоянен и определяется составом липидов клетки-хозяина.
n Липидный состав клеточных мембран изменчив. В меньшей степени это проявляется в животных клетках, находящихся в условиях стабильной внутренней среды. Однако можно модифицировать состав липидов в некоторых мембранах, меняя пищевой рацион. n Липидный состав мембран растений заметно изменяется в зависимости от освещенности, температуры и р. Н. n Еще более изменчив состав бактериальных мембран. Он варьирует не только в зависимости от штамма, но и в пределах одного и того же штамма, а также от условий культивирования и фазы роста. n У вирусов, имеющих липопротеиновую оболочку, липидный состав мембран также не постоянен и определяется составом липидов клетки-хозяина.
 Липидный бислой n Липиды мембран имеют в структуре две различные части: неполярный гидрофобный "хвост" и полярную гидрофильную "голову". Такую двойственную природу соединений называют амфифильной. Липиды мембран образуют двухслойную структуру. Каждый слой состоит из сложных липидов, расположенных таким образом, что неполярные (не несущие зарядов) гидрофобные "хвосты" молекул находятся в тесном контакте друг с другом. Так же контактируют гидрофильные части молекул. Все взаимодействия имеют нековалентный характер. Два монослоя ориентируются "хвост к хвосту" так, что образующаяся структура двойного слоя имеет внутреннюю неполярную часть и две полярные поверхности. n Полярные головки несут на себе отрицательные заряды
Липидный бислой n Липиды мембран имеют в структуре две различные части: неполярный гидрофобный "хвост" и полярную гидрофильную "голову". Такую двойственную природу соединений называют амфифильной. Липиды мембран образуют двухслойную структуру. Каждый слой состоит из сложных липидов, расположенных таким образом, что неполярные (не несущие зарядов) гидрофобные "хвосты" молекул находятся в тесном контакте друг с другом. Так же контактируют гидрофильные части молекул. Все взаимодействия имеют нековалентный характер. Два монослоя ориентируются "хвост к хвосту" так, что образующаяся структура двойного слоя имеет внутреннюю неполярную часть и две полярные поверхности. n Полярные головки несут на себе отрицательные заряды
 n Представление о том, что в основе клеточных мембран лежит двойной липидный слой, было получено еще в 20 -х гг. Было найдено, что если экстрагировать липиды из оболочки эритроцитов, а затем поместить липиды на поверхность водного мениска, то можно рассчитать площадь, занимаемую образовавшимся монослоем липидов. Оказалось, что эта площадь вдвое больше площади, занимаемой поверхностью эритроцитов, из которых были экстрагированы липиды. Было сделано предположение, что в мембранах эритроцитов липиды располагаются в два слоя.
n Представление о том, что в основе клеточных мембран лежит двойной липидный слой, было получено еще в 20 -х гг. Было найдено, что если экстрагировать липиды из оболочки эритроцитов, а затем поместить липиды на поверхность водного мениска, то можно рассчитать площадь, занимаемую образовавшимся монослоем липидов. Оказалось, что эта площадь вдвое больше площади, занимаемой поверхностью эритроцитов, из которых были экстрагированы липиды. Было сделано предположение, что в мембранах эритроцитов липиды располагаются в два слоя.
 Фосфолипиды n представляют собой сложные эфиры трехатомного спирта глицерина с двумя жирными кислотами и с фосфорной кислотой, которая в свою очередь может быть связана с различными химическими группами (холин, серин, инозит, этаноламин и др. ). Так, например, в структуру наиболее часто встречающегося в мембранах фосфолипида лецитина входят участки двух жирных кислот, глицерина, фосфорной кислоты и холина.
Фосфолипиды n представляют собой сложные эфиры трехатомного спирта глицерина с двумя жирными кислотами и с фосфорной кислотой, которая в свою очередь может быть связана с различными химическими группами (холин, серин, инозит, этаноламин и др. ). Так, например, в структуру наиболее часто встречающегося в мембранах фосфолипида лецитина входят участки двух жирных кислот, глицерина, фосфорной кислоты и холина.
 и") Фосфолипиды n Основное же количество фосфолипидов мембран приходится, как правило, на фосфатидилхолин (ФХ) и фосфатидилэтаноламин (ФЭА) И специфические липиды n фосфатидилсерин (ФС), сфингомиелин (СМ), фосфатидилинозитол (ФИ), фосфатидилглицерол (кардиолипин) (ФГ).
Фосфолипиды n Основное же количество фосфолипидов мембран приходится, как правило, на фосфатидилхолин (ФХ) и фосфатидилэтаноламин (ФЭА) И специфические липиды n фосфатидилсерин (ФС), сфингомиелин (СМ), фосфатидилинозитол (ФИ), фосфатидилглицерол (кардиолипин) (ФГ).
 функции структурная транспорт ионов и метаболитов, влияют на активность мембраносвязанных ферментов, межклеточные взаимодействия и рецепция. дипальмитоиллецитин является основным элементом сурфактанта (ПАВ) легких n Фосфолипиды, содержащие инозитол , являются предшественниками вторых посредников при действии гормонов, n алкилфосфолипид - тромбоцит-активирующим фактором ( ФАТ ). n n n
функции структурная транспорт ионов и метаболитов, влияют на активность мембраносвязанных ферментов, межклеточные взаимодействия и рецепция. дипальмитоиллецитин является основным элементом сурфактанта (ПАВ) легких n Фосфолипиды, содержащие инозитол , являются предшественниками вторых посредников при действии гормонов, n алкилфосфолипид - тромбоцит-активирующим фактором ( ФАТ ). n n n
 Гликолипиды n В молекулах гликолипидов моно n n n - и олигосахариды присоединены взамен фосфатных групп. Наиболее распространен-ными из гликолипидов являются: у животных – производные сфингозина, у растений – глицерола (по большей части галактозилдиглицериды). У бактерий встречаются в основном глицерогликолипиды.
Гликолипиды n В молекулах гликолипидов моно n n n - и олигосахариды присоединены взамен фосфатных групп. Наиболее распространен-ными из гликолипидов являются: у животных – производные сфингозина, у растений – глицерола (по большей части галактозилдиглицериды). У бактерий встречаются в основном глицерогликолипиды.
 участвуют в межклеточных взаимодействиях, n 2) являются рецепторами бактериальных токсинов ,") Функции n 1) участвуют в межклеточных взаимодействиях, n 2) являются рецепторами бактериальных токсинов , например холерного токсина, n 3) рецепторы или корецепторы гормонов, n 4) являются соединениями, определяющими группы крови (система АВО).
Функции n 1) участвуют в межклеточных взаимодействиях, n 2) являются рецепторами бактериальных токсинов , например холерного токсина, n 3) рецепторы или корецепторы гормонов, n 4) являются соединениями, определяющими группы крови (система АВО).
 Холестерол Жесткие молекулы холестерола погружены в мембрану между молекул фосфолипидов. Гидрофобное четырехчленное стероидное кольцо молекулы холестерола взаимодействует с цепями остатков жирных кислот, входящих в состав фосфолипидов мембраны. В эукариотических клетках холестерол ограничивает текучесть мембраны при температуре 370 С. при более низких температурах он, наоборот, способствует поддержанию текучести мембраны, препятствуя слипанию углеводородных цепей.
Холестерол Жесткие молекулы холестерола погружены в мембрану между молекул фосфолипидов. Гидрофобное четырехчленное стероидное кольцо молекулы холестерола взаимодействует с цепями остатков жирных кислот, входящих в состав фосфолипидов мембраны. В эукариотических клетках холестерол ограничивает текучесть мембраны при температуре 370 С. при более низких температурах он, наоборот, способствует поддержанию текучести мембраны, препятствуя слипанию углеводородных цепей.
 Холестерол n Доля, приходящаяся на стеролы, может достигать 17 – 20% по n n весу. В миелиновых мембранах – до 50% от веса всех липидов. холестерола много в плазматических мембранах животных, у высших растений преобладающими стероидами плазмолеммы являются ситостерол и стигмостерол, у грибов – эргостерол.
Холестерол n Доля, приходящаяся на стеролы, может достигать 17 – 20% по n n весу. В миелиновых мембранах – до 50% от веса всех липидов. холестерола много в плазматических мембранах животных, у высших растений преобладающими стероидами плазмолеммы являются ситостерол и стигмостерол, у грибов – эргостерол.
 ХОЛЕСТЕРОЛ n Придает механическую прочность бислою, заполняя свободное пространство, n большую текучесть, способен свободно перетекать из слоя в слой, обеспечивая изменение формы мембран при сжатии и растяжении. n Холестерол в некоторых мембранах может занимать до 25% по массе. n Мембраны прокариотических клеток не содержат холестерол.
ХОЛЕСТЕРОЛ n Придает механическую прочность бислою, заполняя свободное пространство, n большую текучесть, способен свободно перетекать из слоя в слой, обеспечивая изменение формы мембран при сжатии и растяжении. n Холестерол в некоторых мембранах может занимать до 25% по массе. n Мембраны прокариотических клеток не содержат холестерол.
 Особенности липидного состава мембран различного функционального назначения n Наиболее изменчив липидный состав плазматической мембраны, поскольку количество холестерола, обилием которого она отличается, в значительной степени зависит от особенностей питания организма. n Миелиновые мембраны имеют необычно высокую концентрацию сфинголипидов и гликосфинголипидов. n Мембраны ядра и ЭПР, содержат, фосфоглицеролы и, в меньшем количестве, сфинголипиды и холестерол. n ПК по этому показателю занимает промежуточное положение между ЭПС и плазмолеммой, характеризуясь меньшим содержанием фосфоглицеролов и большим – холестерола. n Особняком стоят мембраны митохондрий: если наружная мембрана отличается от мембраны ЭПС только появлением ФГ, то внутренняя обеднена СМ и обогащена ФГ.
Особенности липидного состава мембран различного функционального назначения n Наиболее изменчив липидный состав плазматической мембраны, поскольку количество холестерола, обилием которого она отличается, в значительной степени зависит от особенностей питания организма. n Миелиновые мембраны имеют необычно высокую концентрацию сфинголипидов и гликосфинголипидов. n Мембраны ядра и ЭПР, содержат, фосфоглицеролы и, в меньшем количестве, сфинголипиды и холестерол. n ПК по этому показателю занимает промежуточное положение между ЭПС и плазмолеммой, характеризуясь меньшим содержанием фосфоглицеролов и большим – холестерола. n Особняком стоят мембраны митохондрий: если наружная мембрана отличается от мембраны ЭПС только появлением ФГ, то внутренняя обеднена СМ и обогащена ФГ.
 Жирные кислоты липидов мембран Свойства n Тканеспецифичность n В мембране никогда не находятся в свободном состоянии, Длина углеродной цепи всегда в пределах 12 -22 атома углерода n ЖК мембран животных чётное количество атомов углерода. У растений, у прокариот, нечётное n Молекулы ЖК, как правило, линейны, Углеводородная часть ЖК может быть как насыщенной, так и ненасыщенной, содержащей одну (мононенасыщенные ЖК) или несколько (полиненасыщенные ЖК) двойных связей
Жирные кислоты липидов мембран Свойства n Тканеспецифичность n В мембране никогда не находятся в свободном состоянии, Длина углеродной цепи всегда в пределах 12 -22 атома углерода n ЖК мембран животных чётное количество атомов углерода. У растений, у прокариот, нечётное n Молекулы ЖК, как правило, линейны, Углеводородная часть ЖК может быть как насыщенной, так и ненасыщенной, содержащей одну (мононенасыщенные ЖК) или несколько (полиненасыщенные ЖК) двойных связей
 Асимметрия мембран n Свойством мембран является диспропорциональное распределение отдельных сортов фосфолипидов между двумя монослоями липидного бислоя n Наружный монослой плазматической мембраны обогащён ФХ либо СМ, тогда как внутренний – ФЭА и ФС.
Асимметрия мембран n Свойством мембран является диспропорциональное распределение отдельных сортов фосфолипидов между двумя монослоями липидного бислоя n Наружный монослой плазматической мембраны обогащён ФХ либо СМ, тогда как внутренний – ФЭА и ФС.
 Подвижность n Диапазон движений, происходящих в мембране, весьма широк: от молекулярных колебаний с частотой порядка 1014 с до трансмембранного флип-флоп-переноса липидов, характерное время которого может достигать несколько суток
Подвижность n Диапазон движений, происходящих в мембране, весьма широк: от молекулярных колебаний с частотой порядка 1014 с до трансмембранного флип-флоп-переноса липидов, характерное время которого может достигать несколько суток
 Подвижность мембранных липидов n Вращательное движение n Латеральное движение n Флип-флоп
Подвижность мембранных липидов n Вращательное движение n Латеральное движение n Флип-флоп
 Флип-флоп липидов в ЭПР
Флип-флоп липидов в ЭПР
, n -полуинтегральные (погруженные в липиды частично) n") Белки мембран n -интегральные (полностью пронизываю-щие мембрану), n -полуинтегральные (погруженные в липиды частично) n -периферические (ассоциированные с мембраной и взаимодействующие с полярными головками фосфолипидов).
Белки мембран n -интегральные (полностью пронизываю-щие мембрану), n -полуинтегральные (погруженные в липиды частично) n -периферические (ассоциированные с мембраной и взаимодействующие с полярными головками фосфолипидов).
 Интегральные белки n глобулярные белки. n Молекулярный вес 10 – 1. Монотопный белок, а-спиральный домен которого пронизывает мембрану. 2. Политопный белок, пронизывающий мембрану несколько раз. 500 к. Да n Много гидрофобных аминокислотных остатков (23 -24). n Достаточное количество полярных остатков.
Интегральные белки n глобулярные белки. n Молекулярный вес 10 – 1. Монотопный белок, а-спиральный домен которого пронизывает мембрану. 2. Политопный белок, пронизывающий мембрану несколько раз. 500 к. Да n Много гидрофобных аминокислотных остатков (23 -24). n Достаточное количество полярных остатков.
, активный насосный механизм n формирование межклеточных контактов") Функции n транспортная (ионные каналы), активный насосный механизм n формирование межклеточных контактов
Функции n транспортная (ионные каналы), активный насосный механизм n формирование межклеточных контактов
 Полуинтегральные белки n Глобулярные n белки могут экспонироваться только на одной поверхности n n n мембраны. В этом случае молекула белка состоит из двух участков – гидрофобного «якоря» и гидрофильного функционального участка. Функции специфические рецепторы транспортная (белки-переносчики) ферментные белки переносчики электронов
Полуинтегральные белки n Глобулярные n белки могут экспонироваться только на одной поверхности n n n мембраны. В этом случае молекула белка состоит из двух участков – гидрофобного «якоря» и гидрофильного функционального участка. Функции специфические рецепторы транспортная (белки-переносчики) ферментные белки переносчики электронов
 Периферические белки n Глобулярные и фибриллярные n взаимодействуют только с полярными компонентами – головками фосфолипидов и немембранными доменами интегральных белков. На поверхности мембран периферические белки удерживаются за счёт электростатических сил и солевых мостиков На наружной поверхности - ламинин и фибронектин, на внутренней - спектрин и альфа-актинин.
Периферические белки n Глобулярные и фибриллярные n взаимодействуют только с полярными компонентами – головками фосфолипидов и немембранными доменами интегральных белков. На поверхности мембран периферические белки удерживаются за счёт электростатических сил и солевых мостиков На наружной поверхности - ламинин и фибронектин, на внутренней - спектрин и альфа-актинин.
 Функции n Рецепторные n Адгезионные n Белки цитоскелета n Белки системы вторичных посредников n Антитела n Ферменты
Функции n Рецепторные n Адгезионные n Белки цитоскелета n Белки системы вторичных посредников n Антитела n Ферменты
 Асимметрия белкового состава n Структурная асимметрия тесно связана с функциональной. n На внешней стороне мембраны находятся рецепторные и маркерные молекулы. Внутренняя сторона обеспечивает выполнение исполнительских функций, на ней расположены ферменты, белки, связанные с цитоскелетом и т. д.
Асимметрия белкового состава n Структурная асимметрия тесно связана с функциональной. n На внешней стороне мембраны находятся рецепторные и маркерные молекулы. Внутренняя сторона обеспечивает выполнение исполнительских функций, на ней расположены ферменты, белки, связанные с цитоскелетом и т. д.
 всегда связаны ковалентными связями") Углеводные компоненты n Углеводные остатки различной длины (моно- и олигосахариды) всегда связаны ковалентными связями либо с липидами, либо с белками. n Углеводные цепи плазматической мембраны экспонированы во внеклеточную среду, располагаясь над плоскостью липидного бислоя. n Углеводные компоненты обнаруживаются и в составе мембран внутриклеточных органоидов. В этом случае они экспонированы во внутренний просвет органоида.
Углеводные компоненты n Углеводные остатки различной длины (моно- и олигосахариды) всегда связаны ковалентными связями либо с липидами, либо с белками. n Углеводные цепи плазматической мембраны экспонированы во внеклеточную среду, располагаясь над плоскостью липидного бислоя. n Углеводные компоненты обнаруживаются и в составе мембран внутриклеточных органоидов. В этом случае они экспонированы во внутренний просвет органоида.
 Углеводный состав n Среди мономеров в составе углеводных цепей - галактоза и манноза, n реже – ксилоза и глюкоза, и сиаловая кислота. Терминальным мономером является фукоза или сиаловая кислота. n В молекулах гликолипидов моно- и олигосахариды присоединены взамен фосфатных групп.
Углеводный состав n Среди мономеров в составе углеводных цепей - галактоза и манноза, n реже – ксилоза и глюкоза, и сиаловая кислота. Терминальным мономером является фукоза или сиаловая кислота. n В молекулах гликолипидов моно- и олигосахариды присоединены взамен фосфатных групп.
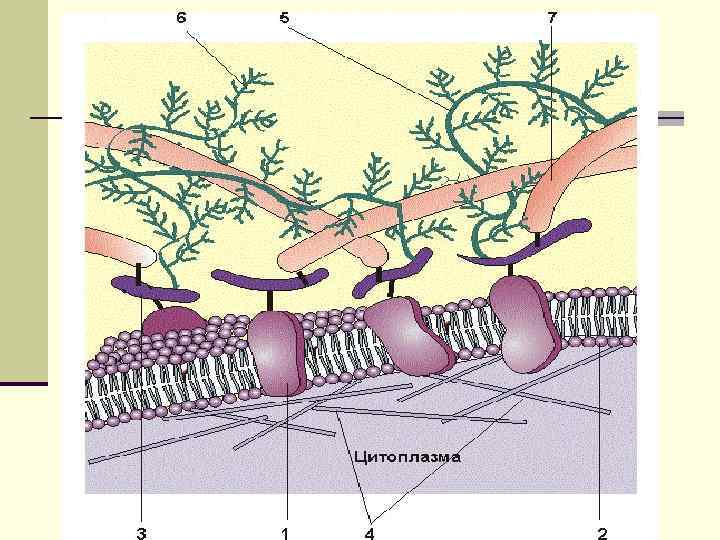
 Функции углеводов мембран n Углеводные цепи этих соединений формируют основную часть гликокаликса эукариотических клеток n Большая часть мембранных рецепторов – гликопротеиды или ганглиозиды, углеводная составляющая которых играет важную роль в реализации трансмембранной передачи сигнала. n Гликопротеидами являются и отдельные компоненты ионных насосов, например, малая субъединица Na+/K+-АТФазы. n Антигены групп крови АВ 0, локализованные на плазматической мембране эритроцитов, представляют собой углеводные компоненты гликопротеидов.
Функции углеводов мембран n Углеводные цепи этих соединений формируют основную часть гликокаликса эукариотических клеток n Большая часть мембранных рецепторов – гликопротеиды или ганглиозиды, углеводная составляющая которых играет важную роль в реализации трансмембранной передачи сигнала. n Гликопротеидами являются и отдельные компоненты ионных насосов, например, малая субъединица Na+/K+-АТФазы. n Антигены групп крови АВ 0, локализованные на плазматической мембране эритроцитов, представляют собой углеводные компоненты гликопротеидов.
 Неорганические компоненты мембран n Вода. 30% веса мембраны. Само существование липидного бислоя возможно как раз по причине водного окружения мембран. n Ca 2+, Mg 2+, K+ и Na+. связываются с участками кислых фосфолипидов, а также белков, включающих гидрофильные кислые домены. Они оказывают влияние на структурнофункциональныые характеристики мембран. Это влияние реализуется через изменение конформационного состояния белков, статического заряда поверхностей мембран и т. д. Обеспечивают ионный гомеостаз n Некоторые мембранные белки содержат ионы металлов.
Неорганические компоненты мембран n Вода. 30% веса мембраны. Само существование липидного бислоя возможно как раз по причине водного окружения мембран. n Ca 2+, Mg 2+, K+ и Na+. связываются с участками кислых фосфолипидов, а также белков, включающих гидрофильные кислые домены. Они оказывают влияние на структурнофункциональныые характеристики мембран. Это влияние реализуется через изменение конформационного состояния белков, статического заряда поверхностей мембран и т. д. Обеспечивают ионный гомеостаз n Некоторые мембранные белки содержат ионы металлов.
 Функции мембраны n n n n Транспортная Избирательная проницаемость Коммуникационная Межклеточная адгезия Межклеточные контакты Ферментативная Мембраны несут специфические рецепторы, связывающие биологически активные соединения
Функции мембраны n n n n Транспортная Избирательная проницаемость Коммуникационная Межклеточная адгезия Межклеточные контакты Ферментативная Мембраны несут специфические рецепторы, связывающие биологически активные соединения
 Функции n благодаря рецепторной и ферментативной активности мембран осуществляется трансмембранная передача сигналов n на мембранах находятся специфические молекулярные маркеры, позволяющие опознавать данный тип мембранного органоида (в случае плазмолеммы – тип клетки) n мембраны обеспечивают сопряжение процессов окисления и фоторецепции с синтезом макроэргических соединений n мембранные пузырьки образуются в результате отшнуровки от существующих мембранных образований и способны сливаться с другими мембранами
Функции n благодаря рецепторной и ферментативной активности мембран осуществляется трансмембранная передача сигналов n на мембранах находятся специфические молекулярные маркеры, позволяющие опознавать данный тип мембранного органоида (в случае плазмолеммы – тип клетки) n мембраны обеспечивают сопряжение процессов окисления и фоторецепции с синтезом макроэргических соединений n мембранные пузырьки образуются в результате отшнуровки от существующих мембранных образований и способны сливаться с другими мембранами
 Транспорт Пассивный транспорт n Диффузия n Облегченная диффузия n Осмос Активный транспорт Экзо - или эндоцитоз.
Транспорт Пассивный транспорт n Диффузия n Облегченная диффузия n Осмос Активный транспорт Экзо - или эндоцитоз.
 Пассивный транспорт: диффузия Движение молекул по градиенту концентрации n Холестерин, бензол n Небольшие неполярные молекулы О 2 и N 2 n Небольшие полярные Н 2 О, С О 2 , этанол
Пассивный транспорт: диффузия Движение молекул по градиенту концентрации n Холестерин, бензол n Небольшие неполярные молекулы О 2 и N 2 n Небольшие полярные Н 2 О, С О 2 , этанол
 Потенциалзависимые, механочувствительные, рецепторзависимые, кальций-зависимые)") Облегченная диффузия Ионный канал n Ионные каналы (натриевые, кальциевые, аквапорины) Потенциалзависимые, механочувствительные, рецепторзависимые, кальций-зависимые)
Облегченная диффузия Ионный канал n Ионные каналы (натриевые, кальциевые, аквапорины) Потенциалзависимые, механочувствительные, рецепторзависимые, кальций-зависимые)
 Лиганд-зависимые ионные каналы n Эти каналы открываются, когда медиатор, связываясь с их наружными сторонами, меняет их конформацию. Открываясь, они впускают ионы, изменяя этим мембранный потенциал. Лигандзависимые каналы почти нечувствительны к изменению мембранного потенциала.
Лиганд-зависимые ионные каналы n Эти каналы открываются, когда медиатор, связываясь с их наружными сторонами, меняет их конформацию. Открываясь, они впускают ионы, изменяя этим мембранный потенциал. Лигандзависимые каналы почти нечувствительны к изменению мембранного потенциала.
 Облегченная диффузия Белки-переносчики n Белки-переносчики (глюкоза, аминокислоты, анионообменники – обмен Cl- на бикарбонат, фосфаты, обмен Na+ на Н+, норадреналин) По градиенту концентрации без затраты энергии, м. б. сочетанным, специфичен
Облегченная диффузия Белки-переносчики n Белки-переносчики (глюкоза, аминокислоты, анионообменники – обмен Cl- на бикарбонат, фосфаты, обмен Na+ на Н+, норадреналин) По градиенту концентрации без затраты энергии, м. б. сочетанным, специфичен
 Активный транспорт n Энергозависимый трансмембранный перенос против электрохимического градиента, происходит при участии АТФаз n Na+К+-, H+, K+- и Ca 2+- АТФазы
Активный транспорт n Энергозависимый трансмембранный перенос против электрохимического градиента, происходит при участии АТФаз n Na+К+-, H+, K+- и Ca 2+- АТФазы
 Активный транспорт: ионные насосы
Активный транспорт: ионные насосы
 Системы антипорта n или контр-транспорта – это система вторично активного транспорта, функционирующая на основе переносчика обменника, обеспечивающего выведение из клетки транспортирующего вещества против его концентрационного градиента в обмен на сопряженный, пассивно поступающий в клетки поток ионов Na+. Движущей силой является потенциальная энергия концентрационного градиента ионов Na+. n Антипортная система клеток проксимальных отделов нефрона, обменивающая Nа+ на Н+. Из мочи к клетки проксимальных отделов нефрона выделяется Nа+, взамен выводится Н+. Система не совершает электрическую работу, поэтому не надо тратить энергию.
Системы антипорта n или контр-транспорта – это система вторично активного транспорта, функционирующая на основе переносчика обменника, обеспечивающего выведение из клетки транспортирующего вещества против его концентрационного градиента в обмен на сопряженный, пассивно поступающий в клетки поток ионов Na+. Движущей силой является потенциальная энергия концентрационного градиента ионов Na+. n Антипортная система клеток проксимальных отделов нефрона, обменивающая Nа+ на Н+. Из мочи к клетки проксимальных отделов нефрона выделяется Nа+, взамен выводится Н+. Система не совершает электрическую работу, поэтому не надо тратить энергию.
 Везикулярный перенос n Крупные молекулы биополимеров практически не проникают сквозь плазмолемму. В ряде случаев макромолекулы и даже их агрегаты, а часто и крупные частицы попадают внутрь клетки в результате процесса эндоцитоза . Эндоцитоз формально разделяют на фагоцитоз (захват и поглощение клеткой крупных частиц, например бактерий или фрагментов других клеток) и пиноцитоз (захват отдельных молекул и макромолекулярных соединений).
Везикулярный перенос n Крупные молекулы биополимеров практически не проникают сквозь плазмолемму. В ряде случаев макромолекулы и даже их агрегаты, а часто и крупные частицы попадают внутрь клетки в результате процесса эндоцитоза . Эндоцитоз формально разделяют на фагоцитоз (захват и поглощение клеткой крупных частиц, например бактерий или фрагментов других клеток) и пиноцитоз (захват отдельных молекул и макромолекулярных соединений).
 Макромолекулы такие как белки, нуклеиновые кислоты, полисахариды, липопротеидные комплексы и другие сквозь клеточные мембраны не проходят, в противовес тому как транспортируются ионы и мономеры. Транспорт микромолекул, их комплексов, частиц внутрь клетки и из нее происходит совершенно иным путем - посредством везикулярного переноса. Т. е макромолекулы, не могут попадать в клетку сквозь плазматическую мембрану. В клетку они попадают заключенными внутри вакуолей или везикул. Такой везикулярный перенос делят на два вида: экзоцитоз - вынос из клетки макромолекулярных продуктов, и эндоцитоз – поглощение клеткой макромолекул.
Макромолекулы такие как белки, нуклеиновые кислоты, полисахариды, липопротеидные комплексы и другие сквозь клеточные мембраны не проходят, в противовес тому как транспортируются ионы и мономеры. Транспорт микромолекул, их комплексов, частиц внутрь клетки и из нее происходит совершенно иным путем - посредством везикулярного переноса. Т. е макромолекулы, не могут попадать в клетку сквозь плазматическую мембрану. В клетку они попадают заключенными внутри вакуолей или везикул. Такой везикулярный перенос делят на два вида: экзоцитоз - вынос из клетки макромолекулярных продуктов, и эндоцитоз – поглощение клеткой макромолекул.
 Эндоцитоз n Эндоцитоз начинается с сорбции на поверхности плазмолеммы поглощаемых веществ. Связывание их с плазмолеммой определяется наличием на ее поверхности рецепторных молекул. После сорбции веществ на поверхности плазмолемма начинает образовывать сначала небольшие впячивания внутрь клетки. Они могут иметь вид еще не замкнутых округлых пузырьков или представлять собой глубокие инвагинации, впячивания внутрь клетки. Затем такие локальные впячивания отшнуровываются от плазмолеммы и в виде пузырьков свободно располагаются под ней.
Эндоцитоз n Эндоцитоз начинается с сорбции на поверхности плазмолеммы поглощаемых веществ. Связывание их с плазмолеммой определяется наличием на ее поверхности рецепторных молекул. После сорбции веществ на поверхности плазмолемма начинает образовывать сначала небольшие впячивания внутрь клетки. Они могут иметь вид еще не замкнутых округлых пузырьков или представлять собой глубокие инвагинации, впячивания внутрь клетки. Затем такие локальные впячивания отшнуровываются от плазмолеммы и в виде пузырьков свободно располагаются под ней.
 В такую первичную вакуоль, или в эндосому, могут попадать любые биополимеры, макромолекулярные комплексы, части клеток или даже целые клетки, где затем и распадаются, деполимеризуются до мономеров, которые путем трансмембранного переноса попадают в гиалоплазму. n Клетка эпителия тонкого кишечника образует до 1000 пиносом в секунду, а макрофаги образуют около 125 пиносом в минуту. n Размер пиносом составляет 60 -130 нм
В такую первичную вакуоль, или в эндосому, могут попадать любые биополимеры, макромолекулярные комплексы, части клеток или даже целые клетки, где затем и распадаются, деполимеризуются до мономеров, которые путем трансмембранного переноса попадают в гиалоплазму. n Клетка эпителия тонкого кишечника образует до 1000 пиносом в секунду, а макрофаги образуют около 125 пиносом в минуту. n Размер пиносом составляет 60 -130 нм
 эндоцитоз
эндоцитоз
 Фагоцитоз захват клеткой крупных частицх, встречается среди клеток животных, как одноклеточных (амебы, некоторые хищные инфузории), так и в специализированных клетках многоклеточных животных. Специализированные клетки- фагоциты - характерны для беспозвоночных животных (амебоциты крови или полостной жидкости), и для позвоночных (нейтрофилы и макрофаги).
Фагоцитоз захват клеткой крупных частицх, встречается среди клеток животных, как одноклеточных (амебы, некоторые хищные инфузории), так и в специализированных клетках многоклеточных животных. Специализированные клетки- фагоциты - характерны для беспозвоночных животных (амебоциты крови или полостной жидкости), и для позвоночных (нейтрофилы и макрофаги).
. n Неспецифический эндоцитоз протекает автоматически") Эндоцитоз может быть неспецифическим и специфическим, опосредуемым рецепторами (рецепторным). n Неспецифический эндоцитоз протекает автоматически и часто приводит к захвату и поглощению совершенно чуждых или безразличных для клетки веществ( сажа или красители).
Эндоцитоз может быть неспецифическим и специфическим, опосредуемым рецепторами (рецепторным). n Неспецифический эндоцитоз протекает автоматически и часто приводит к захвату и поглощению совершенно чуждых или безразличных для клетки веществ( сажа или красители).
 Рецепторный эндоцитоз Специальные рецепторы плазматической мембраны, диффузно расположенные на поверхности различных клеток, узнают вещество, и образуют специфический комплекс рецептор-лиганд. Вслед за этим такой комплекс перемещается в зону окаймленных ямок и интернализуется - окружается мембраной и погружается вглубь цитоплазмы. Со стороны цитоплазмы плазматическая мембрана покрыта тонким (около 20 нм) волокнистым слоем, окаймляет небольшие ямки. Занимают около 2% клеточной поверхности. Состоит в основном из белка клатрина.
Рецепторный эндоцитоз Специальные рецепторы плазматической мембраны, диффузно расположенные на поверхности различных клеток, узнают вещество, и образуют специфический комплекс рецептор-лиганд. Вслед за этим такой комплекс перемещается в зону окаймленных ямок и интернализуется - окружается мембраной и погружается вглубь цитоплазмы. Со стороны цитоплазмы плазматическая мембрана покрыта тонким (около 20 нм) волокнистым слоем, окаймляет небольшие ямки. Занимают около 2% клеточной поверхности. Состоит в основном из белка клатрина.
 Клатриновый слой одевает весь периметр отделяющихся первичных эндоцитозных вакуолей - окаймленных пузырьков. Эти белки связываются с интегральными белкамирецепторами со стороны цитоплазмы и образуют одевающий слой по периметру возникающей пиносомы, - “окаймленного” пузырька. Затем окаймленный пузырек отделится от плазмолеммы и переносится вглубь цитоплазмы, клатриновый слой распадается. После потери клатринового слоя эндосомы начинают сливаться друг с другом.
Клатриновый слой одевает весь периметр отделяющихся первичных эндоцитозных вакуолей - окаймленных пузырьков. Эти белки связываются с интегральными белкамирецепторами со стороны цитоплазмы и образуют одевающий слой по периметру возникающей пиносомы, - “окаймленного” пузырька. Затем окаймленный пузырек отделится от плазмолеммы и переносится вглубь цитоплазмы, клатриновый слой распадается. После потери клатринового слоя эндосомы начинают сливаться друг с другом.
 Основное биологическое значение эндоцитоза получение строительных блоков за счет внутриклеточного переваривания, которое осуществляется на втором этапе эндоцитоза после слияния первичной эндосомы с лизосомой.
Основное биологическое значение эндоцитоза получение строительных блоков за счет внутриклеточного переваривания, которое осуществляется на втором этапе эндоцитоза после слияния первичной эндосомы с лизосомой.
 Экзоцитоз процесс, обратный эндоцитозу. Внутриклеточные продукты, заключенные в вакуоли и отграниченные от гиалоплазмы мембраной, подходят к плазматической мембране. В местах их контактов плазматическая мембрана и мембрана вакуоли сливаются, и пузырек опустошается в окружающую среду. С экзоцитозом связано выделение синтезированных в клетке разнообразных веществ. Секретирующие, выделяющие вещества во внешнюю среду, клетки могут вырабатывать и выбрасывать низкомолекулярные соединения (ацетилхолин, биогенные амины и др. ), а также в большинстве случаев макромолекулы (пептиды, белки, липопротеиды, пептидогликаны и др. ). Экзоцитоз происходит в ответ на внешний сигнал (нервный импульс, гормоны, медиаторы.
Экзоцитоз процесс, обратный эндоцитозу. Внутриклеточные продукты, заключенные в вакуоли и отграниченные от гиалоплазмы мембраной, подходят к плазматической мембране. В местах их контактов плазматическая мембрана и мембрана вакуоли сливаются, и пузырек опустошается в окружающую среду. С экзоцитозом связано выделение синтезированных в клетке разнообразных веществ. Секретирующие, выделяющие вещества во внешнюю среду, клетки могут вырабатывать и выбрасывать низкомолекулярные соединения (ацетилхолин, биогенные амины и др. ), а также в большинстве случаев макромолекулы (пептиды, белки, липопротеиды, пептидогликаны и др. ). Экзоцитоз происходит в ответ на внешний сигнал (нервный импульс, гормоны, медиаторы.
 Значение экзоцтоза n секреция веществ для других клеток (секреция молока, пищеварительных соков, гормонов и др. ). n секреция для собственных нужд. (рост плазматической мембраны, элементы гликокаликса ) n экзоцитоз гидролитических ферментов для пристеночного пищеварения
Значение экзоцтоза n секреция веществ для других клеток (секреция молока, пищеварительных соков, гормонов и др. ). n секреция для собственных нужд. (рост плазматической мембраны, элементы гликокаликса ) n экзоцитоз гидролитических ферментов для пристеночного пищеварения
 Надмембранные структуры
Надмембранные структуры
 Гликокаликс представляет собой внешний по отношению к мембране слой, содержащий олиго- и полисахаридные цепочки мембранных интегральных белков - гликопротеидов. Эти цепочки содержат такие углеводы как манноза, глюкоза, n-ацетилгликозамин, сиаловая кислота и др. Такие углеводные гетерополимеры образуют ветвящиеся цепочки, между которыми могут располагаться выделенные из клетки гликолипиды и протеогликаны.
Гликокаликс представляет собой внешний по отношению к мембране слой, содержащий олиго- и полисахаридные цепочки мембранных интегральных белков - гликопротеидов. Эти цепочки содержат такие углеводы как манноза, глюкоза, n-ацетилгликозамин, сиаловая кислота и др. Такие углеводные гетерополимеры образуют ветвящиеся цепочки, между которыми могут располагаться выделенные из клетки гликолипиды и протеогликаны.
 Слой гликокаликса сильно обводнен, имеет желеподобную консистенцию, что значительно снижает в этой зоне скорость диффузии различных веществ. Здесь располагаются выделенные клеткой гидролитические ферменты, участвующие во внеклеточном расщеплении полимеров (внеклеточное пищеварение) до мономерных молекул, которые затем транспортируются в цитоплазму через плазматическую мембрану.
Слой гликокаликса сильно обводнен, имеет желеподобную консистенцию, что значительно снижает в этой зоне скорость диффузии различных веществ. Здесь располагаются выделенные клеткой гидролитические ферменты, участвующие во внеклеточном расщеплении полимеров (внеклеточное пищеварение) до мономерных молекул, которые затем транспортируются в цитоплазму через плазматическую мембрану.
 В электронном микроскопе, при специальных методах контрастирования полисахаридов, гликокаликс имеет вид рыхлого волокнистого слоя, толщиной 3 -4 нм, покрывающего всю поверхность клетки. Особенно хорошо гликокаликс выражен в щеточной каемке клеток всасывающего эпителия кишечника (энтероциты)
В электронном микроскопе, при специальных методах контрастирования полисахаридов, гликокаликс имеет вид рыхлого волокнистого слоя, толщиной 3 -4 нм, покрывающего всю поверхность клетки. Особенно хорошо гликокаликс выражен в щеточной каемке клеток всасывающего эпителия кишечника (энтероциты)
 Гликокаликс
Гликокаликс
 Состав n Толщина – 45 -55 нм. n олигосахариды, n периферические белки n гликопротеиды n гликолипиды n сиаловая кислота
Состав n Толщина – 45 -55 нм. n олигосахариды, n периферические белки n гликопротеиды n гликолипиды n сиаловая кислота
 Строение n Ячеистая структура сети гликокаликса обусловлена наличием анионнокатионных подвижных мостиков между нитями. Размер ячеек сети гликокаликса уменьшается по направлению к плазмалемме.
Строение n Ячеистая структура сети гликокаликса обусловлена наличием анионнокатионных подвижных мостиков между нитями. Размер ячеек сети гликокаликса уменьшается по направлению к плазмалемме.
 Функции Он предназначен для восприятия, распознавания, передачи, модуляции сигналов клетки в виде вещества, энергии, информации Выступающие разветвленные части гликокаликса обеспечивают: n межклеточное узнавание n индивидуальность клетки; n иммунологическую индивидуальность клетки (роль антигенов); n Межклеточные взаимодействия соединение клеток при образовании тканей.
Функции Он предназначен для восприятия, распознавания, передачи, модуляции сигналов клетки в виде вещества, энергии, информации Выступающие разветвленные части гликокаликса обеспечивают: n межклеточное узнавание n индивидуальность клетки; n иммунологическую индивидуальность клетки (роль антигенов); n Межклеточные взаимодействия соединение клеток при образовании тканей.
 Межклеточное узнавание n Это специфическое взаимодействие клетки с другой клеткой или с внеклеточным матриксом. После узнавания - Прекращение миграции клеток - Адгезия клеток - Образование контактов - Образование клеточных ансамблей - Взаимодействие клеток между собой и клетками других структур
Межклеточное узнавание n Это специфическое взаимодействие клетки с другой клеткой или с внеклеточным матриксом. После узнавания - Прекращение миграции клеток - Адгезия клеток - Образование контактов - Образование клеточных ансамблей - Взаимодействие клеток между собой и клетками других структур
 n Семейство кадгеринов – трансмембранные гликопротеины n Иммуноглобулины. n интегрины n распознавание клетками друга при формировании тканевых структур, взаимодействие клеток с компонентами межклеточного вещества и прикрепление к ним.
n Семейство кадгеринов – трансмембранные гликопротеины n Иммуноглобулины. n интегрины n распознавание клетками друга при формировании тканевых структур, взаимодействие клеток с компонентами межклеточного вещества и прикрепление к ним.
 Пристеночное пищеварение n В гликокаликсе энтероцитов начинается примембранное пищеварение которое сопрягает процессы полостного пищеварения и всасывания питательных веществ в тонкой кишке.
Пристеночное пищеварение n В гликокаликсе энтероцитов начинается примембранное пищеварение которое сопрягает процессы полостного пищеварения и всасывания питательных веществ в тонкой кишке.
 Индивидуализация n Углеводный компонент гликокаликса играет важную роль в индивидуализации разных типов клеток. Презентация– лимфоциты которые в зависимости от дифференцировки на них антигенов подразделяются на несколько типов – В-, Т -лимфоциты и НК-клетки
Индивидуализация n Углеводный компонент гликокаликса играет важную роль в индивидуализации разных типов клеток. Презентация– лимфоциты которые в зависимости от дифференцировки на них антигенов подразделяются на несколько типов – В-, Т -лимфоциты и НК-клетки
 Создание поверхностного заряда n В эритроцитах млекопитающих развит углеводный компонент интегрального гликопротеина – гликофорина, который необходим для создания отрицательного заряда на поверхности эритроцитов
Создание поверхностного заряда n В эритроцитах млекопитающих развит углеводный компонент интегрального гликопротеина – гликофорина, который необходим для создания отрицательного заряда на поверхности эритроцитов
 Субмембранный слой
Субмембранный слой
 слой n Кортикальный (cortex -кора) слой цитоплазмы, лежащий в тесном контакте с") Субмембранный (кортикальный) слой n Кортикальный (cortex -кора) слой цитоплазмы, лежащий в тесном контакте с мембраной. Основным фибриллярным компонентом кортикального слоя является сеть актиновых микрофибрилл. Здесь же располагается ряд вспомогательных белков, необходимых для движения участков цитоплазмы.
Субмембранный (кортикальный) слой n Кортикальный (cortex -кора) слой цитоплазмы, лежащий в тесном контакте с мембраной. Основным фибриллярным компонентом кортикального слоя является сеть актиновых микрофибрилл. Здесь же располагается ряд вспомогательных белков, необходимых для движения участков цитоплазмы.
 Компоненты цитоскелета n 1. микротрубочки 22± 2 нм n 2. микрофиламенты 5 -7 нм (актин, миозин, актинин, тропомиозин) n 3. промежуточные филаменты -10 нм (прекератин, виментин, десмин) –
Компоненты цитоскелета n 1. микротрубочки 22± 2 нм n 2. микрофиламенты 5 -7 нм (актин, миозин, актинин, тропомиозин) n 3. промежуточные филаменты -10 нм (прекератин, виментин, десмин) –
 Функции n Поддержание формы клетки n Изменение формы клетки
Функции n Поддержание формы клетки n Изменение формы клетки