

2 Структура клетки_СУРС 1_студ.ppt
- Количество слайдов: 101
 УО «Гомельский государственный университет имени Франциска Скорины» Кафедра ботаники и физиологии растений ФИЗИОЛОГИЯ РАСТЕНИЙ СУРС 1 Раздел 2 Физиология растительной клетки Тема 1 Особенности структуры растительной клетки (см. текст СУРС 1 Структура растительной клетки. pdf)
УО «Гомельский государственный университет имени Франциска Скорины» Кафедра ботаники и физиологии растений ФИЗИОЛОГИЯ РАСТЕНИЙ СУРС 1 Раздел 2 Физиология растительной клетки Тема 1 Особенности структуры растительной клетки (см. текст СУРС 1 Структура растительной клетки. pdf)
 «Для того чтобы понять жизнь растения. . . необходимо прежде ознакомиться с его формой; для того чтобы понять действие машины, нужно знать ее устройство» . К. А. Тимирязев Вопросы темы для самостоятельного изучения 1. Структурная организация клетки - основа ее биохимической активности и функционирования как целостной живой системы. 2. Структура, функция отдельных компонентов и органоидов клетки, особенности их строения в связи с их биологическими функциями. 3. Мембранный принцип организации поверхности протоплазмы и органоидов растительной клетки. Биохимическая и функциональная разнокачественность мембран. Основные функции мембран.
«Для того чтобы понять жизнь растения. . . необходимо прежде ознакомиться с его формой; для того чтобы понять действие машины, нужно знать ее устройство» . К. А. Тимирязев Вопросы темы для самостоятельного изучения 1. Структурная организация клетки - основа ее биохимической активности и функционирования как целостной живой системы. 2. Структура, функция отдельных компонентов и органоидов клетки, особенности их строения в связи с их биологическими функциями. 3. Мембранный принцип организации поверхности протоплазмы и органоидов растительной клетки. Биохимическая и функциональная разнокачественность мембран. Основные функции мембран.
 1. Структурная организация клетки основа ее биохимической активности и функционирования как целостной живой системы.
1. Структурная организация клетки основа ее биохимической активности и функционирования как целостной живой системы.
 Термин «клетка» от греч. «cytos» — клетка или лат. «cellula» — полость. Основные даты в истории открытия и изучения клетки: 1665 г. − обнаруживший в растительных тканях особые замкнутые образования Р. Гук вводит термин «клетка» . 1833 г. − открытие английским ученым Робертом Броуном ядра в эпидермисе листа орхидных растений. 1839 г. − М. Я. Шлейденом и Т. Шванном была сформулирована клеточная теория. 1898 г. − итальянский цитолог К. Гольджи обнаружил в нервных клетках совы и кошки сетчатые структуры, которые затем так и назвали — аппарат Гольджи. 1945 г. − американским ученым К. Портером посредством электронной микроскопии был обнаружен эндоплазматический ретикулум.
Термин «клетка» от греч. «cytos» — клетка или лат. «cellula» — полость. Основные даты в истории открытия и изучения клетки: 1665 г. − обнаруживший в растительных тканях особые замкнутые образования Р. Гук вводит термин «клетка» . 1833 г. − открытие английским ученым Робертом Броуном ядра в эпидермисе листа орхидных растений. 1839 г. − М. Я. Шлейденом и Т. Шванном была сформулирована клеточная теория. 1898 г. − итальянский цитолог К. Гольджи обнаружил в нервных клетках совы и кошки сетчатые структуры, которые затем так и назвали — аппарат Гольджи. 1945 г. − американским ученым К. Портером посредством электронной микроскопии был обнаружен эндоплазматический ретикулум.
 Рис. 1. Соотношение между отдельными видами структур во взрослой растительной клетке: Клетка Клеточная оболочка Ядро Микроскопические структуры Органоиды Вакуоль Протопласт Плазмалемма Цитоплазма Субмикроскопические структуры Гиалоплазма Пластиды Митохондрии Эндоплазматический ретикулюм Аппарат Гольджи Рибосомы Микротела Пероксисомы, глиоксисомы. Сферосомы Цитоскелет Микрофиламенты Микротрубочки
Рис. 1. Соотношение между отдельными видами структур во взрослой растительной клетке: Клетка Клеточная оболочка Ядро Микроскопические структуры Органоиды Вакуоль Протопласт Плазмалемма Цитоплазма Субмикроскопические структуры Гиалоплазма Пластиды Митохондрии Эндоплазматический ретикулюм Аппарат Гольджи Рибосомы Микротела Пероксисомы, глиоксисомы. Сферосомы Цитоскелет Микрофиламенты Микротрубочки
 Рис. 2. Схема строения растительной клетки
Рис. 2. Схема строения растительной клетки
.") Рис 3. Схема строения клетки мезофилла листа (по Полевому В. В. ).
Рис 3. Схема строения клетки мезофилла листа (по Полевому В. В. ).
, – комплекс Гольджи. Их совместная деятельность обеспечивает") Транспортная система клетки: – эндоплазматический ретикулум (ЭР), – комплекс Гольджи. Их совместная деятельность обеспечивает • • синтез и преобразование веществ в клетке, их изоляцию, накопление и транспорт
Транспортная система клетки: – эндоплазматический ретикулум (ЭР), – комплекс Гольджи. Их совместная деятельность обеспечивает • • синтез и преобразование веществ в клетке, их изоляцию, накопление и транспорт
 В отличие от клеток других эукариотических организмов для растительной клетки характерны: 1. пластидная система, возникшая в связи с фототрофным способом питания, 2. полисахаридная клеточная стенка, окружающая клетку, 3. центральная вакуоль в зрелых клетках. Кроме того, • у делящейся растительной клетки нет центриолей.
В отличие от клеток других эукариотических организмов для растительной клетки характерны: 1. пластидная система, возникшая в связи с фототрофным способом питания, 2. полисахаридная клеточная стенка, окружающая клетку, 3. центральная вакуоль в зрелых клетках. Кроме того, • у делящейся растительной клетки нет центриолей.
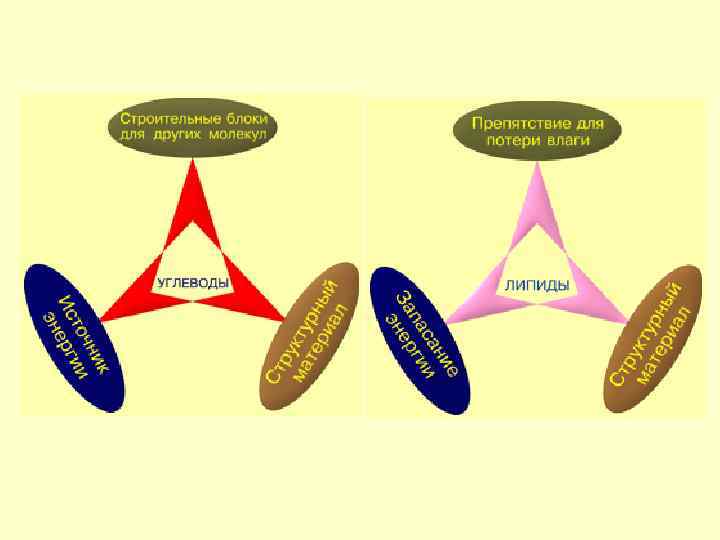
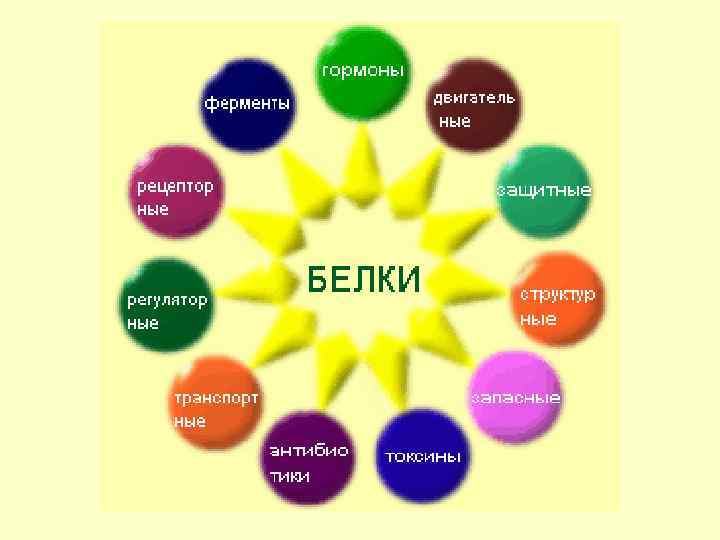
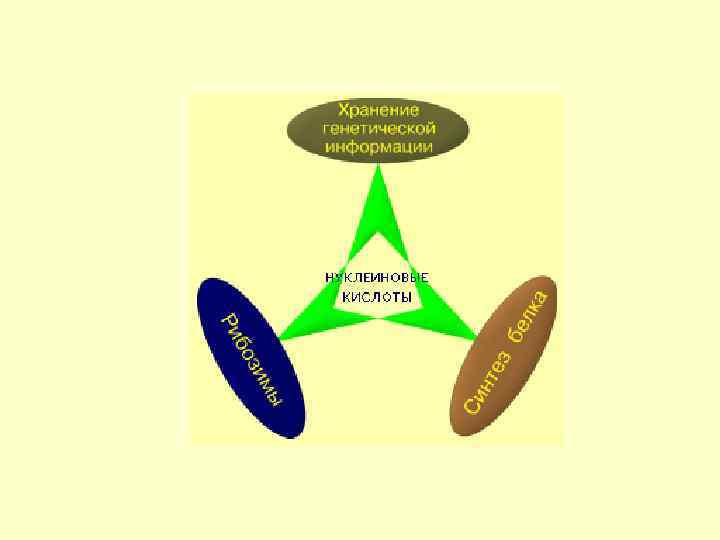
 Вода 75 -85 Белки") Содержание в клетке химических соединений (в % на сырую массу) Вода 75 -85 Белки 10 -20 Жиры 1 -5 Углеводы НК 0, 2 -2 1 -2 Низкомолекулярные органические соединения 0, 1 - 0, 5 Неорганические вещества 1 - 1, 5
Содержание в клетке химических соединений (в % на сырую массу) Вода 75 -85 Белки 10 -20 Жиры 1 -5 Углеводы НК 0, 2 -2 1 -2 Низкомолекулярные органические соединения 0, 1 - 0, 5 Неорганические вещества 1 - 1, 5
 2. Структура, функции отдельных компонентов и органоидов клетки. Особенности их строения в связи с их биологическими функциями.
2. Структура, функции отдельных компонентов и органоидов клетки. Особенности их строения в связи с их биологическими функциями.
 2. 1 Клеточная стенка плотная полисахаридная оболочка, выстланной изнутри плазмалеммой. Формируется на стадии телофазы во время митоза. Клеточную стенку делящихся и растущих растяжением клеток называют первичной. После прекращения роста клетки на первичную клеточную стенку изнутри откладываются новые слои и возникает прочная вторичная клеточная стенка.
2. 1 Клеточная стенка плотная полисахаридная оболочка, выстланной изнутри плазмалеммой. Формируется на стадии телофазы во время митоза. Клеточную стенку делящихся и растущих растяжением клеток называют первичной. После прекращения роста клетки на первичную клеточную стенку изнутри откладываются новые слои и возникает прочная вторичная клеточная стенка.
 2) 3) 4) структурные компоненты (целлюлоза у растений,") В состав клеточной стенки входят 1) 2) 3) 4) структурные компоненты (целлюлоза у растений, хитин у грибов), компоненты матрикса стенки (гемицеллюлозы, пектин, белки), инкрустирующие компоненты (лигнин, суберин) вещества, откладывающиеся на поверхности стенки (кутин и воска). Клеточные стенки могут содержать также силикаты и карбонаты кальция.
В состав клеточной стенки входят 1) 2) 3) 4) структурные компоненты (целлюлоза у растений, хитин у грибов), компоненты матрикса стенки (гемицеллюлозы, пектин, белки), инкрустирующие компоненты (лигнин, суберин) вещества, откладывающиеся на поверхности стенки (кутин и воска). Клеточные стенки могут содержать также силикаты и карбонаты кальция.
 • Углеводные компоненты клеточных стенок целлюлоза, гемицеллюлозы и пектиновые вещества. Целлюлоза и пектиновые вещества адсорбируют воду, обеспечивая оводненность клеточной стенки. Пектиновые вещества, содержащие много карбоксильных групп, связывают ионы двухвалентных металлов, которые способны обмениваться на другие катионы (Н + , К+ и т. д. ). Это обусловливает катионообменную способность клеточных стенок растений. • В состав матрикса клеточной стенки входит структурный белок экстенсин (гликопротеин), который по аминокислотному составу сходен с межклеточным белком животных — коллагеном.
• Углеводные компоненты клеточных стенок целлюлоза, гемицеллюлозы и пектиновые вещества. Целлюлоза и пектиновые вещества адсорбируют воду, обеспечивая оводненность клеточной стенки. Пектиновые вещества, содержащие много карбоксильных групп, связывают ионы двухвалентных металлов, которые способны обмениваться на другие катионы (Н + , К+ и т. д. ). Это обусловливает катионообменную способность клеточных стенок растений. • В состав матрикса клеточной стенки входит структурный белок экстенсин (гликопротеин), который по аминокислотному составу сходен с межклеточным белком животных — коллагеном.
 • Основным инкрустирующим веществом клеточной стенки является лигнин полимер с неразветвленной молекулой, состоящей из ароматических спиртов (п-кумарового, кониферилового, синапового). Интенсивная лигнификация клеточных стенок начинается после прекращения роста клетки. Разрушение и конденсация лигнина в почве — один из факторов образования гумуса. • Отложение суберина делает стенки клеток трудно проницаемыми для воды и растворов (например, в эндодерме, перидерме).
• Основным инкрустирующим веществом клеточной стенки является лигнин полимер с неразветвленной молекулой, состоящей из ароматических спиртов (п-кумарового, кониферилового, синапового). Интенсивная лигнификация клеточных стенок начинается после прекращения роста клетки. Разрушение и конденсация лигнина в почве — один из факторов образования гумуса. • Отложение суберина делает стенки клеток трудно проницаемыми для воды и растворов (например, в эндодерме, перидерме).
 • Поверхность эпидермальных клеток растений защищена гидрофобными веществами — кутином и восками. Предшественники этих соединений секретируются из цитоплазмы на поверхность, где и происходит их полимеризация. Слой кутина обычно пронизан полисахаридными компонентами стенки (целлюлозой, пектином) и образует кутикулу. Кутикула участвует в регуляции водного режима тканей и защищает клетки от повреждений и проникновения инфекции. Строение, заложение и рост клеточной оболочки повторить по учебнику Бавтуто Г. А. , Еремин В. М «Ботаника: морфология и анатомия растений» , Мн. , 1997, с. 95 -104.
• Поверхность эпидермальных клеток растений защищена гидрофобными веществами — кутином и восками. Предшественники этих соединений секретируются из цитоплазмы на поверхность, где и происходит их полимеризация. Слой кутина обычно пронизан полисахаридными компонентами стенки (целлюлозой, пектином) и образует кутикулу. Кутикула участвует в регуляции водного режима тканей и защищает клетки от повреждений и проникновения инфекции. Строение, заложение и рост клеточной оболочки повторить по учебнику Бавтуто Г. А. , Еремин В. М «Ботаника: морфология и анатомия растений» , Мн. , 1997, с. 95 -104.
 Поверхностный налет воска, который формирует верхний слой кутикулы, на листе цветной капусты (Brassica oleracea var. botrytis).
Поверхностный налет воска, который формирует верхний слой кутикулы, на листе цветной капусты (Brassica oleracea var. botrytis).
: А — продольный разрез; Б") Рис. 4. Схема строения плазмодесмы (Полевой В. В. ): А — продольный разрез; Б — поперечный разрез
Рис. 4. Схема строения плазмодесмы (Полевой В. В. ): А — продольный разрез; Б — поперечный разрез
 Рис. 5. Схема образования компонентов оболочки в растущей клетке растения Пл – плазмалемма, Д – диктиосома, ПГ – пузырьки Гольджи, Г – гиалоплазма, КО – клеточная оболочка
Рис. 5. Схема образования компонентов оболочки в растущей клетке растения Пл – плазмалемма, Д – диктиосома, ПГ – пузырьки Гольджи, Г – гиалоплазма, КО – клеточная оболочка
 2. 2 Строение биологических мембран Размеры. Толщина биомембран не превышает 6— 10 нм.
2. 2 Строение биологических мембран Размеры. Толщина биомембран не превышает 6— 10 нм.
 Структура. Жидкостно-мозаичная гипотеза строения биологических мембран: основу мембраны составляет двойной слой фосфолипидов с некоторым количеством других липидов (галактолипидов, стеринов, жирных кислот и др. ), причем липиды повернуты друг к другу своими гидрофобными концами (рис. 6). Основную роль в формировании мембран играют гидрофобные связи: липид — липид, липид — белок, белок — белок. Участки молекул полярных липидов, образованные ненасыщенными жирными кислотами, и стерины обеспечивают несколько разрыхленное (жидкое) состояние бислоя. Наружная и внутренняя стороны биологических мембран обращены в качественно разные гидрофильные среды. Это является причиной асимметричного строения мембран в наружном слое плазмалеммы содержится больше стеринов и гликолипидов.
Структура. Жидкостно-мозаичная гипотеза строения биологических мембран: основу мембраны составляет двойной слой фосфолипидов с некоторым количеством других липидов (галактолипидов, стеринов, жирных кислот и др. ), причем липиды повернуты друг к другу своими гидрофобными концами (рис. 6). Основную роль в формировании мембран играют гидрофобные связи: липид — липид, липид — белок, белок — белок. Участки молекул полярных липидов, образованные ненасыщенными жирными кислотами, и стерины обеспечивают несколько разрыхленное (жидкое) состояние бислоя. Наружная и внутренняя стороны биологических мембран обращены в качественно разные гидрофильные среды. Это является причиной асимметричного строения мембран в наружном слое плазмалеммы содержится больше стеринов и гликолипидов.
 Рис. 6. Схема строения цитоплазматической мембраны: 1 — фосфолипид; 2 — холестерол; 3 — глобулярный мембранный белок; 4 — углеводные цепочки; 5 — неполярный участок мембранного белке.
Рис. 6. Схема строения цитоплазматической мембраны: 1 — фосфолипид; 2 — холестерол; 3 — глобулярный мембранный белок; 4 — углеводные цепочки; 5 — неполярный участок мембранного белке.
 Липиды, входящие в состав мембранного бислоя, не закреплены жестко, а непрерывно меняются местами. Перемещения липидных молекул бывают двух типов: 1) в пределах своего монослоя (латеральная диффузия) и 2) путем перестановки двух липидных молекул, противостоящих другу в двух монослоях ( «флип-флоп» ). При латеральной диффузии молекулы липидов претерпевают миллионы перестановок в секунду, а скорость ее составляет 5— 10 мкм/с. Перестановки молекул липидов из одного монослоя в другой происходят значительно реже, но могут ускоряться мембранными белками.
Липиды, входящие в состав мембранного бислоя, не закреплены жестко, а непрерывно меняются местами. Перемещения липидных молекул бывают двух типов: 1) в пределах своего монослоя (латеральная диффузия) и 2) путем перестановки двух липидных молекул, противостоящих другу в двух монослоях ( «флип-флоп» ). При латеральной диффузии молекулы липидов претерпевают миллионы перестановок в секунду, а скорость ее составляет 5— 10 мкм/с. Перестановки молекул липидов из одного монослоя в другой происходят значительно реже, но могут ускоряться мембранными белками.
 Строительным "кирпичиком" мембраны являются фосфолипиды. Вследствие своего строения фосфолипиды амфифильны, поскольку обладают полярной "головой", образованной заряженной группой фосфата (иногда ионизированной группировкой, присоединенной к ней, пример- фосфатидилхолин). Полярной частью молекула хорошо взаимодействует с водной средой. Кроме того, у фосфолипидов имеется довольно длинный неполярный (гидрофобный) "хвост". Хвост образован остатками карбоновых кислот, очень часто- ненасыщенных, содержащих кратную связь примерно в середине цепи. Кратная связь чаще всего находится в цис-конфигурации, поэтому "ножка" не прямая, а как бы изогнутая (см. рис. , (а). Если образец фосфолипида размешать в водной среде, образуются так называемые мицеллы (b) построенные так, что полярные головы обращены в водный слой, а неполярные хвосты - внутрь мицеллы. Мицеллы являются формой, через которую происходит усвоение липидов в организме. В определенных условиях можно получить липосомы, состоящие из бислоя липидов и очень напоминающие клетку, только в миниатюре. У липосом имеется внутреннее пространство, практически изолированное от внешней среды вследствие того, что структура липосомы чрезвычайно стабильна. Способность некоторых липидов к "самосборке" в двойные слои является очень важным свойством, имеющим решающую роль в построении клеточных мембран.
Строительным "кирпичиком" мембраны являются фосфолипиды. Вследствие своего строения фосфолипиды амфифильны, поскольку обладают полярной "головой", образованной заряженной группой фосфата (иногда ионизированной группировкой, присоединенной к ней, пример- фосфатидилхолин). Полярной частью молекула хорошо взаимодействует с водной средой. Кроме того, у фосфолипидов имеется довольно длинный неполярный (гидрофобный) "хвост". Хвост образован остатками карбоновых кислот, очень часто- ненасыщенных, содержащих кратную связь примерно в середине цепи. Кратная связь чаще всего находится в цис-конфигурации, поэтому "ножка" не прямая, а как бы изогнутая (см. рис. , (а). Если образец фосфолипида размешать в водной среде, образуются так называемые мицеллы (b) построенные так, что полярные головы обращены в водный слой, а неполярные хвосты - внутрь мицеллы. Мицеллы являются формой, через которую происходит усвоение липидов в организме. В определенных условиях можно получить липосомы, состоящие из бислоя липидов и очень напоминающие клетку, только в миниатюре. У липосом имеется внутреннее пространство, практически изолированное от внешней среды вследствие того, что структура липосомы чрезвычайно стабильна. Способность некоторых липидов к "самосборке" в двойные слои является очень важным свойством, имеющим решающую роль в построении клеточных мембран.
 В состав мембран входят разные в функциональном отношении белки: белки-ферменты, белки, выполняющие функции насосов, переносчиков, ионных каналов, белки-регуляторы и структурные белки (рис. 7). Если такие специализированные протеиновые комплексы • погружены в липидную фазу и удерживаются гидрофобными связями (липопротеины), их называют «интегральные белки» ; • удерживаются на внутренней и внешней поверхностях мембран электростатическими связями (гидрофильные белки), взаимодействуя с гидрофильными головками полярных липидов, то их называют «периферические белки» ;
В состав мембран входят разные в функциональном отношении белки: белки-ферменты, белки, выполняющие функции насосов, переносчиков, ионных каналов, белки-регуляторы и структурные белки (рис. 7). Если такие специализированные протеиновые комплексы • погружены в липидную фазу и удерживаются гидрофобными связями (липопротеины), их называют «интегральные белки» ; • удерживаются на внутренней и внешней поверхностях мембран электростатическими связями (гидрофильные белки), взаимодействуя с гидрофильными головками полярных липидов, то их называют «периферические белки» ;
 Рис. 7. Схема строения цитоплазматической мембраны и основные группы мембранных белков
Рис. 7. Схема строения цитоплазматической мембраны и основные группы мембранных белков
 Интегральные белковые глобулы располагаются в фосфолипидных слоях мембран ориентированно. Определенные участки липопротеиновых глобул (участки узнавания) служат для самосборки полиэнзимных мембранных комплексов. На положение белков в мембране оказывают влияние состав фосфолипидов (свободных и связанных), величина электростатического заряда мембраны. Функциональная активность мембран и изменения мембранного потенциала сопровождаются всплыванием или погружением субъединиц, их латеральными перемещениями.
Интегральные белковые глобулы располагаются в фосфолипидных слоях мембран ориентированно. Определенные участки липопротеиновых глобул (участки узнавания) служат для самосборки полиэнзимных мембранных комплексов. На положение белков в мембране оказывают влияние состав фосфолипидов (свободных и связанных), величина электростатического заряда мембраны. Функциональная активность мембран и изменения мембранного потенциала сопровождаются всплыванием или погружением субъединиц, их латеральными перемещениями.
 соком и окруженная мембраной") 2. 3 Вакуолярная система Вакуоль это полость, заполненная клеточным (вакуолярным) соком и окруженная мембраной (тонопластом) (рис. 8). В меристематических клетках вакуоли представлены мелкими пузырьками. Для зрелых клеток характерна большая центральная вакуоль. Рис. 8. Вакуоли растительных клеток
2. 3 Вакуолярная система Вакуоль это полость, заполненная клеточным (вакуолярным) соком и окруженная мембраной (тонопластом) (рис. 8). В меристематических клетках вакуоли представлены мелкими пузырьками. Для зрелых клеток характерна большая центральная вакуоль. Рис. 8. Вакуоли растительных клеток
 Сначала образуются провакуоли.") Пути формирования вакуолярной системы: • из расширенных цистерн ЭР (рис. 9) Сначала образуются провакуоли. Слияние провакуолей приводит к возникновению более крупных вакуолей и образованию вакуолярной мембраны — тонопласта. Тонопласт может образовывать инвагинации, что приводит к включению в вакуоль участков цитоплазмы; • путем автофагии Процесс начинается с окружения мембраной эндоплазматического ретикулюма (ЭР) участка цитоплазмы (автофагическая вакуоль). Деятельность кислых гидролаз в замкнутом мембраной пространстве приводит к разрушению полимерного содержимого и поступлению воды. Процесс вакуолизации — необходимое условие роста клеток растяжением.
Пути формирования вакуолярной системы: • из расширенных цистерн ЭР (рис. 9) Сначала образуются провакуоли. Слияние провакуолей приводит к возникновению более крупных вакуолей и образованию вакуолярной мембраны — тонопласта. Тонопласт может образовывать инвагинации, что приводит к включению в вакуоль участков цитоплазмы; • путем автофагии Процесс начинается с окружения мембраной эндоплазматического ретикулюма (ЭР) участка цитоплазмы (автофагическая вакуоль). Деятельность кислых гидролаз в замкнутом мембраной пространстве приводит к разрушению полимерного содержимого и поступлению воды. Процесс вакуолизации — необходимое условие роста клеток растяжением.
:") 4 3 2 5 1 Рис. 9. Онтогенез вакуолей (по Рубину Б. А. ): 1 — недавно образовавшаяся вакуоль, 2 — более развитая вакуоль с окружающим ее тонопластом, 3 — основная плазма, 4 — тяж эндоплазматической сети, 5 — митохондрия
4 3 2 5 1 Рис. 9. Онтогенез вакуолей (по Рубину Б. А. ): 1 — недавно образовавшаяся вакуоль, 2 — более развитая вакуоль с окружающим ее тонопластом, 3 — основная плазма, 4 — тяж эндоплазматической сети, 5 — митохондрия
 Вакуолярный сок Имеет р. Н 5, 0 -6, 5, но может быть равной 1, 0 (бегония) или 2, 0 (лимон). Включает органические вещества и минеральные соли. В состав вакуолярного сока входят: • соединения, которые могут быть вторично использованы в обмене веществ: органические кислоты, углеводы, аминокислоты и белки. Вакуоль может служить местом отложения запасных белков (алейроновые зерна); • вещества, выведенные из обмена веществ и изолированные от цитоплазмы в вакуоли: фенолы, таннины, алкалоиды, антоцианы; • ферменты. Большинство из них − гидролазы (расщепляют полимеры до низкомолекулярных веществ) с оптимумом активности при кислом р. Н;
Вакуолярный сок Имеет р. Н 5, 0 -6, 5, но может быть равной 1, 0 (бегония) или 2, 0 (лимон). Включает органические вещества и минеральные соли. В состав вакуолярного сока входят: • соединения, которые могут быть вторично использованы в обмене веществ: органические кислоты, углеводы, аминокислоты и белки. Вакуоль может служить местом отложения запасных белков (алейроновые зерна); • вещества, выведенные из обмена веществ и изолированные от цитоплазмы в вакуоли: фенолы, таннины, алкалоиды, антоцианы; • ферменты. Большинство из них − гидролазы (расщепляют полимеры до низкомолекулярных веществ) с оптимумом активности при кислом р. Н;
 Системы доставки веществ в вакуоль: • АТР-зависимая Н+-помпа, выносящая ионы Н+ из цитоплазмы в вакуоль (рис. 10 ). Обеспечивает поступление в вакуоль анионов органических кислот, сахаров, вход и выход ионов К+; • система переносчиков в тонопласте − обусловливает накопление в вакуолях аминокислот и других соединений. Цитоплазма Вакуоль р. Н 5, 0 р. Н 7, 4 Рис. 10. Н+-насос на основе АТР-азы Тонопласт
Системы доставки веществ в вакуоль: • АТР-зависимая Н+-помпа, выносящая ионы Н+ из цитоплазмы в вакуоль (рис. 10 ). Обеспечивает поступление в вакуоль анионов органических кислот, сахаров, вход и выход ионов К+; • система переносчиков в тонопласте − обусловливает накопление в вакуолях аминокислот и других соединений. Цитоплазма Вакуоль р. Н 5, 0 р. Н 7, 4 Рис. 10. Н+-насос на основе АТР-азы Тонопласт
 поддержание гомеостаза растительной клетки; 2) является одним из факторов, регулирующих поступление") Функции вакуоли: 1) поддержание гомеостаза растительной клетки; 2) является одним из факторов, регулирующих поступление и передвижение воды и минеральных солей в клетке, т. е. обуславливающих осмотические свойства растительной клетки; 3) является одним из основных факторов, обуславливающих рост клетки растяжением; 4) может служить местом отложения запасных питательных веществ.
Функции вакуоли: 1) поддержание гомеостаза растительной клетки; 2) является одним из факторов, регулирующих поступление и передвижение воды и минеральных солей в клетке, т. е. обуславливающих осмотические свойства растительной клетки; 3) является одним из основных факторов, обуславливающих рост клетки растяжением; 4) может служить местом отложения запасных питательных веществ.
 2. 4 Субклеточные структуры
2. 4 Субклеточные структуры
 2. 4. 1 Ядро В молодых меристематических клетках занимает от 1/4 до 1/6 объема протопласта, а в сформировавшихся клетках − от 1/20 до 1/200.
2. 4. 1 Ядро В молодых меристематических клетках занимает от 1/4 до 1/6 объема протопласта, а в сформировавшихся клетках − от 1/20 до 1/200.
 Размеры. Диаметр ~ 10 мкм. Форма. может быть сферическим, удлиненным или лопастным. Химический состав. Белок составляет 75 -80 % от сухой массы, ДНК 14 %, РНК 10 %. РНК ядра сосредоточена в основном в рибосомах, ее количество непостоянно, в связи с чем варьирует и отношение РНК к ДНК.
Размеры. Диаметр ~ 10 мкм. Форма. может быть сферическим, удлиненным или лопастным. Химический состав. Белок составляет 75 -80 % от сухой массы, ДНК 14 %, РНК 10 %. РНК ядра сосредоточена в основном в рибосомах, ее количество непостоянно, в связи с чем варьирует и отношение РНК к ДНК.
, ядрышко. • Ядерная оболочка (кариолемма)") Строение ядра. В ядре различают ядерную оболочку, кариоплазму (нуклеоплазму), ядрышко. • Ядерная оболочка (кариолемма) состоит из двух элементарных мембран — внутренней и наружной. • Ядерная оболочка пронизана порами диаметром 10 -20 нм, через которые транспортируются нуклеиновые кислоты и белки. Ядерные поры не являются статическими образованиями они могут закрываться и одновременно образовываться в других участках мембраны. • С наружной мембраной ядерной оболочки могут быть непосредственно связаны элементы эндоплазматического ретикулума (рис. 11).
Строение ядра. В ядре различают ядерную оболочку, кариоплазму (нуклеоплазму), ядрышко. • Ядерная оболочка (кариолемма) состоит из двух элементарных мембран — внутренней и наружной. • Ядерная оболочка пронизана порами диаметром 10 -20 нм, через которые транспортируются нуклеиновые кислоты и белки. Ядерные поры не являются статическими образованиями они могут закрываться и одновременно образовываться в других участках мембраны. • С наружной мембраной ядерной оболочки могут быть непосредственно связаны элементы эндоплазматического ретикулума (рис. 11).
 Рис. 11. Электронно-микроскопическая фотография ядра и участка гладкого ЭР
Рис. 11. Электронно-микроскопическая фотография ядра и участка гладкого ЭР
 • Внутреннее содержимое ядра − нуклеоплазма. • В нуклеоплазме локализованы участки хроматина, состоящего из комплекса ДНК, РНК и белков. В делящихся клетках хроматин организуется в хромосомы. • В нуклеоплазме ядра содержатся также ферменты и кофакторы, необходимые для обеспечения процессов репликации и транскрипции ДНК, молекулы различных РНК, ферменты, фосфорилирующие и ацетилирующие ядерные белки, ферменты гликолиза и др.
• Внутреннее содержимое ядра − нуклеоплазма. • В нуклеоплазме локализованы участки хроматина, состоящего из комплекса ДНК, РНК и белков. В делящихся клетках хроматин организуется в хромосомы. • В нуклеоплазме ядра содержатся также ферменты и кофакторы, необходимые для обеспечения процессов репликации и транскрипции ДНК, молекулы различных РНК, ферменты, фосфорилирующие и ацетилирующие ядерные белки, ферменты гликолиза и др.
 РНК") Через поры в ядерной оболочке из ядра в цитоплазму выходят молекулы матричной (информационной) РНК (анимация "The Inner Life of the Cell" ).
Через поры в ядерной оболочке из ядра в цитоплазму выходят молекулы матричной (информационной) РНК (анимация "The Inner Life of the Cell" ).
 Основная функция ядра − хранение генетической информации клетки. 2) В ядре") Функции ядра. 1) Основная функция ядра − хранение генетической информации клетки. 2) В ядре происходит − репликация ДНК, − процесс транскрипции ДНК в РНК различных типов. 3) В тесном взаимодействии с цитоплазмой ядро участвует в обеспечении экспрессии генетической информации и контролирует процессы жизнедеятельности клетки. 4) Огромную роль ядро играет в процессе деления клеток.
Функции ядра. 1) Основная функция ядра − хранение генетической информации клетки. 2) В ядре происходит − репликация ДНК, − процесс транскрипции ДНК в РНК различных типов. 3) В тесном взаимодействии с цитоплазмой ядро участвует в обеспечении экспрессии генетической информации и контролирует процессы жизнедеятельности клетки. 4) Огромную роль ядро играет в процессе деления клеток.
 2. 4. 2 Ядрышко В интерфазном ядре имеется одно или несколько ядрышек. Ядрышко формируется на определенных участках ДНК, называемых ядрышковым организатором. В хроматине ядрышка находятся участки ДНК, ответственные за синтез рибосомальных РНК (р. РНК). Основная функция ядрышка − в нем синтезируются ядерные белки.
2. 4. 2 Ядрышко В интерфазном ядре имеется одно или несколько ядрышек. Ядрышко формируется на определенных участках ДНК, называемых ядрышковым организатором. В хроматине ядрышка находятся участки ДНК, ответственные за синтез рибосомальных РНК (р. РНК). Основная функция ядрышка − в нем синтезируются ядерные белки.
 2. 4. 3 Пластиды — это ограниченные двойной мембраной округлые или овальные органоиды, содержащие внутреннюю систему мембран, т. е. имеют ламеллярное строение. Растительные клетки содержат пластиды несколько типов пластид: а) окрашенные пластиды • хлоропласты − имеют зеленую окраску, • хромопласты − имеют желтую, красную или коричневую окраску в зависимости от природы содержащегося в них пигмента; б) бесцветные пластиды • лейкопласты − не содержат пигментов, поэтому лишены окраски, • этиопласты − бесцветные, формируются при выращивании растений в темноте, • протопластиды − присутствуют в меристемах, женских половых клетках, в клетках зародыша.
2. 4. 3 Пластиды — это ограниченные двойной мембраной округлые или овальные органоиды, содержащие внутреннюю систему мембран, т. е. имеют ламеллярное строение. Растительные клетки содержат пластиды несколько типов пластид: а) окрашенные пластиды • хлоропласты − имеют зеленую окраску, • хромопласты − имеют желтую, красную или коричневую окраску в зависимости от природы содержащегося в них пигмента; б) бесцветные пластиды • лейкопласты − не содержат пигментов, поэтому лишены окраски, • этиопласты − бесцветные, формируются при выращивании растений в темноте, • протопластиды − присутствуют в меристемах, женских половых клетках, в клетках зародыша.
 Пластиды всех типов связаны единством происхождения, они образуются из одного и того же предшественника − протопластид (рис. 12). Пластиды различных типов могут взаимно переходить друг в друга. Рис. 12. Биогенез хлоропластов и взаимосвязь компонентов пластидной системы: 1— крахмальные зерна, 2—липидные капли, содержащие каротиноиды, 3— кристаллы пигментов Протопластида Хлоропласт Лейкопласт Хромопласт
Пластиды всех типов связаны единством происхождения, они образуются из одного и того же предшественника − протопластид (рис. 12). Пластиды различных типов могут взаимно переходить друг в друга. Рис. 12. Биогенез хлоропластов и взаимосвязь компонентов пластидной системы: 1— крахмальные зерна, 2—липидные капли, содержащие каротиноиды, 3— кристаллы пигментов Протопластида Хлоропласт Лейкопласт Хромопласт
 Пропластиды При нормальном освещении пропластиды превращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов. Несколько иначе развитие пластид происходит в темноте. У этиолированных проростков происходит в начале увеличение объема пластид, этиопластов, но система внутренних мембран не строит ламеллярные структуры, а образует массу мелких пузырьков, которые скапливаютсяя в отдельные зоны и даже могут формировать сложные решетчатые структуры (проламеллярные тела). В мембранах этиопластов содержится протохлорофилл, предшественник хлорофилла желтого цвета. Под действие света из этиопластов образуются хлоропласты, протохлорофилл превращается в хлорофилл, происходит синтез новых мембран, фотосинтетических ферментов и компонентов цепи переноса электронов. При освещении клеток мембранные пузырьки и трубочки быстро реорганизуются, из них развивается полная система ламелл и тилакоидов, характерная для нормального хлоропласта.
Пропластиды При нормальном освещении пропластиды превращаются в хлоропласты. Сначала они растут, при этом происходит образование продольно расположенных мембранных складок от внутренней мембраны. Одни из них простираются по всей длине пластиды и формируют ламеллы стромы; другие образуют ламеллы тилакоидов, которые выстраиваются в виде стопки и образуют граны зрелых хлоропластов. Несколько иначе развитие пластид происходит в темноте. У этиолированных проростков происходит в начале увеличение объема пластид, этиопластов, но система внутренних мембран не строит ламеллярные структуры, а образует массу мелких пузырьков, которые скапливаютсяя в отдельные зоны и даже могут формировать сложные решетчатые структуры (проламеллярные тела). В мембранах этиопластов содержится протохлорофилл, предшественник хлорофилла желтого цвета. Под действие света из этиопластов образуются хлоропласты, протохлорофилл превращается в хлорофилл, происходит синтез новых мембран, фотосинтетических ферментов и компонентов цепи переноса электронов. При освещении клеток мембранные пузырьки и трубочки быстро реорганизуются, из них развивается полная система ламелл и тилакоидов, характерная для нормального хлоропласта.
 Хлоропласты в клетках растения
Хлоропласты в клетках растения
 Хлоропласты в клетках Декабриста
Хлоропласты в клетках Декабриста
 Хлоропласты. Форма − овальные тельца. Размеры − длина 5 -10 мкм, диаметр 2 -3 мкм. Количество в клетке − 15 -20 в одной клетке листа, у некоторых водорослей − 1 -2 гигантских хлоропласта различной формы. Окраска − зеленая, обусловлена содержанием в хлоропластах хлорофилла. Химический состав. • Структурную основу составляют белки (30 -45 % от сухого веса). Большая часть белков хлоропластов принадлежит к липопротеидам, доля водорастворимых белков незначительна. • Содержание воды − около 75 %. • Важной составной частью являются липиды (20 -40 % от сухого веса), которые находятся как в связанном, так и в свободном состоянии. По мере старения организма повышается содержание свободных липидов. • Общее содержание минеральных веществ в хлоропластах в 2 -3 раза ниже их среднего содержания в листьях в целом. Вместе с тем в них наблюдается исключительно высокое содержание железа, меди, цинка и магния. В хлоропластах сосредоточено до 80 % всего железа, содержащегося в листьях, до 60 -70 % всего цинка и около 50 % меди.
Хлоропласты. Форма − овальные тельца. Размеры − длина 5 -10 мкм, диаметр 2 -3 мкм. Количество в клетке − 15 -20 в одной клетке листа, у некоторых водорослей − 1 -2 гигантских хлоропласта различной формы. Окраска − зеленая, обусловлена содержанием в хлоропластах хлорофилла. Химический состав. • Структурную основу составляют белки (30 -45 % от сухого веса). Большая часть белков хлоропластов принадлежит к липопротеидам, доля водорастворимых белков незначительна. • Содержание воды − около 75 %. • Важной составной частью являются липиды (20 -40 % от сухого веса), которые находятся как в связанном, так и в свободном состоянии. По мере старения организма повышается содержание свободных липидов. • Общее содержание минеральных веществ в хлоропластах в 2 -3 раза ниже их среднего содержания в листьях в целом. Вместе с тем в них наблюдается исключительно высокое содержание железа, меди, цинка и магния. В хлоропластах сосредоточено до 80 % всего железа, содержащегося в листьях, до 60 -70 % всего цинка и около 50 % меди.
. • Это двухмембранные органоиды. • Внутреннее содержимое хлоропластов −") Структура хлоропластов (рис. 13, 14). • Это двухмембранные органоиды. • Внутреннее содержимое хлоропластов − строма (матрикс) представляет собой гомогенную среду. • Хлоропласты, как и другие органоиды, например, митохондрии, имеют ламеллярное строение, т. е. внутренние структуры органоида образованы инвагинациями внутренней мембраны ламеллами. • В хлоропластах внутренняя мембрана образует уплощенные инвагинации — тилакоиды, которые могут иметь форму дисков и в этом случае называются тилакоидами гран. Несколько лежащих друг над другом тилакоидов образуют стопку — грану. Другие тилакоиды, связывающие между собой граны или не контактирующие с ними, называются тилакоидами стромы. При таком строении значительно увеличивается фотоактивная поверхность хлоропластов. и обеспечивается максимальное использование световой энергии. • В мембранах тилакоидов локализованы зеленые (хлорофиллы), желтые и красные (каротиноиды) пигменты, компоненты редоксцепей и запасания энергии, участвующие в поглощении и использовании энергии света.
Структура хлоропластов (рис. 13, 14). • Это двухмембранные органоиды. • Внутреннее содержимое хлоропластов − строма (матрикс) представляет собой гомогенную среду. • Хлоропласты, как и другие органоиды, например, митохондрии, имеют ламеллярное строение, т. е. внутренние структуры органоида образованы инвагинациями внутренней мембраны ламеллами. • В хлоропластах внутренняя мембрана образует уплощенные инвагинации — тилакоиды, которые могут иметь форму дисков и в этом случае называются тилакоидами гран. Несколько лежащих друг над другом тилакоидов образуют стопку — грану. Другие тилакоиды, связывающие между собой граны или не контактирующие с ними, называются тилакоидами стромы. При таком строении значительно увеличивается фотоактивная поверхность хлоропластов. и обеспечивается максимальное использование световой энергии. • В мембранах тилакоидов локализованы зеленые (хлорофиллы), желтые и красные (каротиноиды) пигменты, компоненты редоксцепей и запасания энергии, участвующие в поглощении и использовании энергии света.
. 1 — липидная капля;") Рис. 13. Схема строения хлоропласта (по Полевому В. В. ). 1 — липидная капля; 2 — зерна крахмала; 3 — наружная мембрана; 4 — грана; 5 — ДНК; 6 — рибосома; 7 — строма; 8 — тилакоид граны; 9 — тилакоид стромы; 10 — внутренняя мембрана.
Рис. 13. Схема строения хлоропласта (по Полевому В. В. ). 1 — липидная капля; 2 — зерна крахмала; 3 — наружная мембрана; 4 — грана; 5 — ДНК; 6 — рибосома; 7 — строма; 8 — тилакоид граны; 9 — тилакоид стромы; 10 — внутренняя мембрана.
 Внутренняя среда хлоропласта Грана Зерна крахмала Рис. 14. Электронно-микроскопическая фотография хлоропласта
Внутренняя среда хлоропласта Грана Зерна крахмала Рис. 14. Электронно-микроскопическая фотография хлоропласта
, в которых") • Граны часто имеют форму цилиндров размером 2 мкм (рис. 15), в которых пигментно-липидные слои чередуются с белковыми слоями. Грана Рис. 15. Трехмерная модель группы гран (по Рубину Б. А. ). Тилакоиды
• Граны часто имеют форму цилиндров размером 2 мкм (рис. 15), в которых пигментно-липидные слои чередуются с белковыми слоями. Грана Рис. 15. Трехмерная модель группы гран (по Рубину Б. А. ). Тилакоиды
. На внутренней поверхности их мембран имеются") • Морфологически тилакоиды не гомогенны (рис. 16). На внутренней поверхности их мембран имеются специфические структурные образования, названные Парком квантосомами. Рис. 16. Схема строения тилакоидов хлоропласта (метод криоскопии) (по Рубину Б. А. ): Частицы (квантосомы) Б и В расположены на разных сторонах поверхности тилакоида стромы, частицы А и Г проникли через поверхность мембраны
• Морфологически тилакоиды не гомогенны (рис. 16). На внутренней поверхности их мембран имеются специфические структурные образования, названные Парком квантосомами. Рис. 16. Схема строения тилакоидов хлоропласта (метод криоскопии) (по Рубину Б. А. ): Частицы (квантосомы) Б и В расположены на разных сторонах поверхности тилакоида стромы, частицы А и Г проникли через поверхность мембраны
 • Хлоропласты − это системы, способные к автономному синтезу белков. В них присутствуют низко- и высокомолекулярная РНК, специфическая кольцевая ДНК и ферменты, активирующие аминокислоты. Хлоропласты обладают собственными рибосомами. • При росте клетки количество хлоропластов увеличивается путем деления. Иногда наблюдается почкование хлоропластов. Затем размер дочерних хлоропластов увеличивается. Деление хлоропластов происходит через 6 — 20 ч и не обязательно совпадает с делением ядра. Оно может регулироваться красным светом (660 нм) и устраняется облучением дальним красным светом (730 нм). Деление останавливается также низкой температурой.
• Хлоропласты − это системы, способные к автономному синтезу белков. В них присутствуют низко- и высокомолекулярная РНК, специфическая кольцевая ДНК и ферменты, активирующие аминокислоты. Хлоропласты обладают собственными рибосомами. • При росте клетки количество хлоропластов увеличивается путем деления. Иногда наблюдается почкование хлоропластов. Затем размер дочерних хлоропластов увеличивается. Деление хлоропластов происходит через 6 — 20 ч и не обязательно совпадает с делением ядра. Оно может регулироваться красным светом (660 нм) и устраняется облучением дальним красным светом (730 нм). Деление останавливается также низкой температурой.
 Функции хлоропластов. Основная функция − осуществление фотосинтеза. Кроме того: • В хлоропластах происходит вся сложная цепь процессов превращений первичных продуктов фотосинтеза (наращивание углеродной цепи, образование и распад полимерных форм углеводов* и др. ). • В хлоропластах представлен весь набор биохимических систем, участвующих в синтезе АТФ. * Биохимические системы синтеза и превращения углеводов функционируют в строме хлоропластов. В ней же может откладываться крахмал.
Функции хлоропластов. Основная функция − осуществление фотосинтеза. Кроме того: • В хлоропластах происходит вся сложная цепь процессов превращений первичных продуктов фотосинтеза (наращивание углеродной цепи, образование и распад полимерных форм углеводов* и др. ). • В хлоропластах представлен весь набор биохимических систем, участвующих в синтезе АТФ. * Биохимические системы синтеза и превращения углеводов функционируют в строме хлоропластов. В ней же может откладываться крахмал.
 Хромопласты. Встречаются в клетках как надземных, так и подземных органов растений. Их присутствием объясняется окраска плодов томатов, рябины (рис. 17, А) и др. Это обусловлено тем, что хромопласты содержат в везикулах (пузырьках) стромы каротиноиды. • Хромопласты образуются из хлоропластов и значительно реже их лейкопластов (например, в корне моркови). Процесс обесцвечивания и изменения хлоропластов легко наблюдать при развитии лепестков или при созревании плодов. При этом в пластидах могут накапливаться окрашенные в желтый цвет капельки (глобулы) или в них появляются тела в форме кристаллов. Эти процессы сопряжены с постепенным уменьшением числа мембран в пластиде, с исчезновением хлорофилла и крахмала. Процесс образования окрашенных глобул объясняется тем, что при разрушении ламелл хлоропластов выделяются липидные капли, в которых хорошо растворяются различные пигменты (например, каротиноиды). Таким образом, хромопласты представляют собой дегенерирующие формы пластид, подвернутые липофанерозу – распаду липопротедных комплексов.
Хромопласты. Встречаются в клетках как надземных, так и подземных органов растений. Их присутствием объясняется окраска плодов томатов, рябины (рис. 17, А) и др. Это обусловлено тем, что хромопласты содержат в везикулах (пузырьках) стромы каротиноиды. • Хромопласты образуются из хлоропластов и значительно реже их лейкопластов (например, в корне моркови). Процесс обесцвечивания и изменения хлоропластов легко наблюдать при развитии лепестков или при созревании плодов. При этом в пластидах могут накапливаться окрашенные в желтый цвет капельки (глобулы) или в них появляются тела в форме кристаллов. Эти процессы сопряжены с постепенным уменьшением числа мембран в пластиде, с исчезновением хлорофилла и крахмала. Процесс образования окрашенных глобул объясняется тем, что при разрушении ламелл хлоропластов выделяются липидные капли, в которых хорошо растворяются различные пигменты (например, каротиноиды). Таким образом, хромопласты представляют собой дегенерирующие формы пластид, подвернутые липофанерозу – распаду липопротедных комплексов.
 Рис. 17. Пластиды. А – хромопласты в клетках плодов рябины; Б – лейкопласты в клетках кожицы листа традесканции: 1 – ядро; 2 – лейкоплаты.
Рис. 17. Пластиды. А – хромопласты в клетках плодов рябины; Б – лейкопласты в клетках кожицы листа традесканции: 1 – ядро; 2 – лейкоплаты.
 • Если структура пропластид сохраняется у органоидов зрелых клеток, их называют лейкопластами. В лейкопластах откладываются запасные вещества и названия они получают в зависимости от этих соединений: если запасается крахмал — амилопласты, жиры — элайопласты, белки — протеинопласты и т. д. • Этиопласты формируются при выращивании зеленых растений в темноте. При освещении они превращаются в хлоропласты, при этом внутренняя система мембран подвергается сложным перестройкам.
• Если структура пропластид сохраняется у органоидов зрелых клеток, их называют лейкопластами. В лейкопластах откладываются запасные вещества и названия они получают в зависимости от этих соединений: если запасается крахмал — амилопласты, жиры — элайопласты, белки — протеинопласты и т. д. • Этиопласты формируются при выращивании зеленых растений в темноте. При освещении они превращаются в хлоропласты, при этом внутренняя система мембран подвергается сложным перестройкам.
 Лейкопласты. Если структура пропластид сохраняется у органоидов зрелых клеток, их называют лейкопластами (рис. 17, Б). В лейкопластах откладываются запасные вещества и названия они получают в зависимости от этих соединений: если запасается • крахмал — амилопласты, • жиры — элайопласты, • белки — протеинопласты и т. д.
Лейкопласты. Если структура пропластид сохраняется у органоидов зрелых клеток, их называют лейкопластами (рис. 17, Б). В лейкопластах откладываются запасные вещества и названия они получают в зависимости от этих соединений: если запасается • крахмал — амилопласты, • жиры — элайопласты, • белки — протеинопласты и т. д.
 Лейкопласты отличаются от хлоропластов отсутствием развитой ламеллярной системы. Встречаются они в клетках запасающих тканей. Из-за их неопределенной морфологии лейкопласты трудно отличить от пропластид, а иногда и от митохондрий. Они, как и пропластиды, бедны ламеллами, но тем не менее способны к образованию под влиянием света нормальных тилакоидных структур и к приобретению зеленой окраски. В темноте лейкопласты могут накапливать в проламеллярных телах различные запасные вещества, а в строме лейкопластов откладываются зерна вторичного крахмала. Если в хлоропластах происходит отложение так называемого транзиторного крахмала, который присутствует здесь лишь во время ассимиляции СО 2, то в лейкопластах может происходить истинное запасание крахмала. В некоторых тканях (эндосперм злаков, корневища и клубни) накопление крахмала в лейкопластах приводит к образованию амилопластов, сплошь заполненных гранулами запасного крахмала, расположенных в строме пластиды.
Лейкопласты отличаются от хлоропластов отсутствием развитой ламеллярной системы. Встречаются они в клетках запасающих тканей. Из-за их неопределенной морфологии лейкопласты трудно отличить от пропластид, а иногда и от митохондрий. Они, как и пропластиды, бедны ламеллами, но тем не менее способны к образованию под влиянием света нормальных тилакоидных структур и к приобретению зеленой окраски. В темноте лейкопласты могут накапливать в проламеллярных телах различные запасные вещества, а в строме лейкопластов откладываются зерна вторичного крахмала. Если в хлоропластах происходит отложение так называемого транзиторного крахмала, который присутствует здесь лишь во время ассимиляции СО 2, то в лейкопластах может происходить истинное запасание крахмала. В некоторых тканях (эндосперм злаков, корневища и клубни) накопление крахмала в лейкопластах приводит к образованию амилопластов, сплошь заполненных гранулами запасного крахмала, расположенных в строме пластиды.
 2. 4. 4 Митохондрии
2. 4. 4 Митохондрии
 Митохондрии растительной клетки. Форма − округлые или гантелевидные тельца. Размеры − длина 1 -5 мкм, диаметром 0, 4 -0, 5 мкм. Количество в клетке − от десятков до 5 000. Структура (рис. 18). Митохондрии состоят в основном из белка (60 -65 % сухой массы) и липидов (30 %). • Это двухмембранные органоиды. Толщина наружной и внутренней мембран − 5 -6 нм каждая. • Перимитохондриальное пространство (промежуток между мемранами) заполнено жидкостью типа сыворотки. • Внутренняя мембрана образует различной формы складки − кристы (рис. ). На внутренней поверхности внутренней мембраны расположены грибовидные частицы − оксисомы, содержащие окислительные ферменты. • Внутреннее содержимое митохондрий − матрикс. В матриксе содержатся рибосомы и митохондриальная ДНК (0, 5 %), которая имеет кольцевое строение и отвечает за синтез белков митохондрий. • Митохондрии имеют все типы РНК (1 %), делятся независимо от деления ядра, в клетке образуются от предсуществующих митохондрий путем деления или почкования. Полупериод жизни митохондрий − 5− 10 дней.
Митохондрии растительной клетки. Форма − округлые или гантелевидные тельца. Размеры − длина 1 -5 мкм, диаметром 0, 4 -0, 5 мкм. Количество в клетке − от десятков до 5 000. Структура (рис. 18). Митохондрии состоят в основном из белка (60 -65 % сухой массы) и липидов (30 %). • Это двухмембранные органоиды. Толщина наружной и внутренней мембран − 5 -6 нм каждая. • Перимитохондриальное пространство (промежуток между мемранами) заполнено жидкостью типа сыворотки. • Внутренняя мембрана образует различной формы складки − кристы (рис. ). На внутренней поверхности внутренней мембраны расположены грибовидные частицы − оксисомы, содержащие окислительные ферменты. • Внутреннее содержимое митохондрий − матрикс. В матриксе содержатся рибосомы и митохондриальная ДНК (0, 5 %), которая имеет кольцевое строение и отвечает за синтез белков митохондрий. • Митохондрии имеют все типы РНК (1 %), делятся независимо от деления ядра, в клетке образуются от предсуществующих митохондрий путем деления или почкования. Полупериод жизни митохондрий − 5− 10 дней.
 Рис. 18. Схема строения митохондрии: а – трехмерная структура; б – продольный разрез; в – часть кристы; 1– внутренняя мембрана; 2 – матрикс; 3 – межмембранное (перимитохондриальное) пространство; 4 – наружная мембрана; 5– АТФ-сомы (оксисомы); 6 – липидная капля; 7 – ДНК; 8 – рибосомы
Рис. 18. Схема строения митохондрии: а – трехмерная структура; б – продольный разрез; в – часть кристы; 1– внутренняя мембрана; 2 – матрикс; 3 – межмембранное (перимитохондриальное) пространство; 4 – наружная мембрана; 5– АТФ-сомы (оксисомы); 6 – липидная капля; 7 – ДНК; 8 – рибосомы
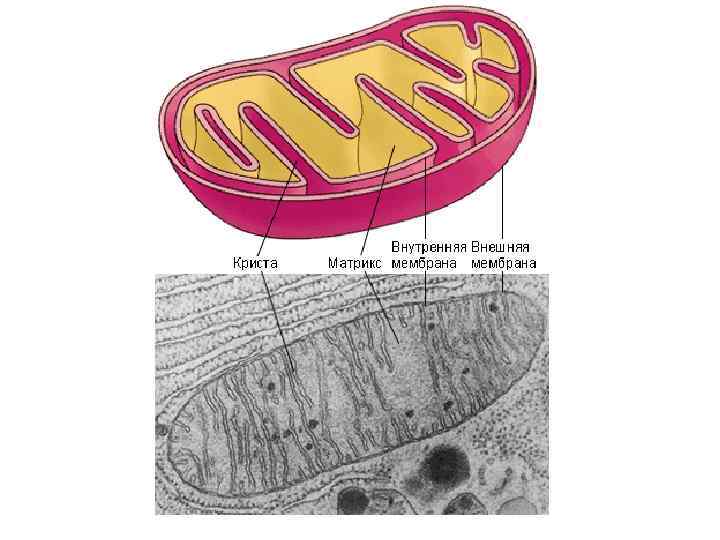
 Функции. Митохондрии являются центрами энергетической активности клеток − в них функционируют системы аэробного дыхания и окислительного фосфорилирования: • Во внутренней мембране митохондрий локализованы компоненты электронтранспортной цепи и АТФ-синтетазные комплексы, осуществляющие транспорт электронов и протонов и синтез АТФ. • В матриксе располагаются системы окисления ди- и трикарбоновых кислот, ряд систем синтеза липидов, аминокислот и др. Митохондрии способны передвигаться к местам усиленного потребления энергии. Они могут ассоциировать друг с другом путем тесного сближения или при помощи тяжей. При анаэробном дыхании митохондрии исчезают.
Функции. Митохондрии являются центрами энергетической активности клеток − в них функционируют системы аэробного дыхания и окислительного фосфорилирования: • Во внутренней мембране митохондрий локализованы компоненты электронтранспортной цепи и АТФ-синтетазные комплексы, осуществляющие транспорт электронов и протонов и синтез АТФ. • В матриксе располагаются системы окисления ди- и трикарбоновых кислот, ряд систем синтеза липидов, аминокислот и др. Митохондрии способны передвигаться к местам усиленного потребления энергии. Они могут ассоциировать друг с другом путем тесного сближения или при помощи тяжей. При анаэробном дыхании митохондрии исчезают.
 Делящиеся митохондрии
Делящиеся митохондрии
 Пероксисома клетки листа. В центре её кристаллическое") 2. 4. 5 Пероксисомы и глиоксисомы (микротела) Пероксисома клетки листа. В центре её кристаллическое белковое ядро.
2. 4. 5 Пероксисомы и глиоксисомы (микротела) Пероксисома клетки листа. В центре её кристаллическое белковое ядро.
 Форма. Округлые органоиды. Размеры. Диаметр − 0, 2 -1, 5 мкм. Количество в клетке. Близко к числу митохондрий. Структура. Ограничены элементарной мембраной, содержат гранулярный матрикс. В некоторых микротелах обнаруживается белковый кристаллоид, состоящий из трубочек диаметром около 6 нм. В клетках растений обнаружены два типа микротел, функционально различающихся : 1) Пероксисомы Многочисленны в клетках листьев. Они тесно связаны с хлоропластами. Функции: а) в пероксисомах окисляется синтезируемая в хлоропластах при фотосинтеза гликолевая кислота и образуется аминокислота глицин; б) в листьях высших растений пероксисомы участвуют в фотодыхании. 2) Глиоксисомы Появляются при прорастании семян, в которых запасаются жиры. Функции: содержат ферменты, необходимые для превращения жирных кислот в сахара: системы β-окисления жирных кислот и глиоксилатный цикл.
Форма. Округлые органоиды. Размеры. Диаметр − 0, 2 -1, 5 мкм. Количество в клетке. Близко к числу митохондрий. Структура. Ограничены элементарной мембраной, содержат гранулярный матрикс. В некоторых микротелах обнаруживается белковый кристаллоид, состоящий из трубочек диаметром около 6 нм. В клетках растений обнаружены два типа микротел, функционально различающихся : 1) Пероксисомы Многочисленны в клетках листьев. Они тесно связаны с хлоропластами. Функции: а) в пероксисомах окисляется синтезируемая в хлоропластах при фотосинтеза гликолевая кислота и образуется аминокислота глицин; б) в листьях высших растений пероксисомы участвуют в фотодыхании. 2) Глиоксисомы Появляются при прорастании семян, в которых запасаются жиры. Функции: содержат ферменты, необходимые для превращения жирных кислот в сахара: системы β-окисления жирных кислот и глиоксилатный цикл.
 Форма. Сферические, сильно преломляющие свет образования") 2. 4. 6 Сферосомы (липидные капли, или олеосомы) Форма. Сферические, сильно преломляющие свет образования Размеры. Диаметр − 0, 5 мкм. Химический состав и строение. Содержат липиды и такие ферменты, как липаза и эстераза. Функции. В них хранятся запасы липидов клетки. При прорастании семян, запасающих жиры, сферосомы функционируют в комплексе с глиоксисомами в процессах глюконеогенеза.
2. 4. 6 Сферосомы (липидные капли, или олеосомы) Форма. Сферические, сильно преломляющие свет образования Размеры. Диаметр − 0, 5 мкм. Химический состав и строение. Содержат липиды и такие ферменты, как липаза и эстераза. Функции. В них хранятся запасы липидов клетки. При прорастании семян, запасающих жиры, сферосомы функционируют в комплексе с глиоксисомами в процессах глюконеогенеза.
, или эндоплазматическая сеть (ЭС) Схематическое представление клеточного ядра,") 2. 4. 7 Эндоплазматический ретикулюм (ЭР), или эндоплазматическая сеть (ЭС) Схематическое представление клеточного ядра, эндоплазматического ретикулума и комплекса Гольджи. (1) Ядро клетки. (2) Поры ядерной мембраны. (3) Гранулярный эндоплазматический ретикулум. (4) Агранулярный эндоплазматический ретикулум. (5) Рибосомы на поверхности гранулярного эндоплазматического ретикулума. (6) Транспортируемые белки. (7) Транспортные везикулы. (8) Комплекс Гольджи
2. 4. 7 Эндоплазматический ретикулюм (ЭР), или эндоплазматическая сеть (ЭС) Схематическое представление клеточного ядра, эндоплазматического ретикулума и комплекса Гольджи. (1) Ядро клетки. (2) Поры ядерной мембраны. (3) Гранулярный эндоплазматический ретикулум. (4) Агранулярный эндоплазматический ретикулум. (5) Рибосомы на поверхности гранулярного эндоплазматического ретикулума. (6) Транспортируемые белки. (7) Транспортные везикулы. (8) Комплекс Гольджи
 Размеры. Объем полостей ЭР может достигать 16% объема клетки. Строение. Представляет собой систему каналов, пузырьков и цистерн, ограниченную мембраной толщиной 5 -6 нм. ЭР может содержать на своей поверхности рибосомы (гранулярный, или шероховатый, ЭР) или не содержать их (агранулярный, или гладкий, ЭР) (рис. 19, 20 ). Эндоплазматическая сеть — очень лабильная структура. Неблагоприятные внешние воздействия (недостаток кислорода и др. ) вызывают концентрические закручивания мембран ретикулума.
Размеры. Объем полостей ЭР может достигать 16% объема клетки. Строение. Представляет собой систему каналов, пузырьков и цистерн, ограниченную мембраной толщиной 5 -6 нм. ЭР может содержать на своей поверхности рибосомы (гранулярный, или шероховатый, ЭР) или не содержать их (агранулярный, или гладкий, ЭР) (рис. 19, 20 ). Эндоплазматическая сеть — очень лабильная структура. Неблагоприятные внешние воздействия (недостаток кислорода и др. ) вызывают концентрические закручивания мембран ретикулума.
 Рис. 19. Эндоплазматический ретикулум : а – гладкий; б – шероховатый; 1– рибосомы, 2 – ядерная оболочка; 3 – ядро; 4 – клеточная тенка
Рис. 19. Эндоплазматический ретикулум : а – гладкий; б – шероховатый; 1– рибосомы, 2 – ядерная оболочка; 3 – ядро; 4 – клеточная тенка
 Рис. 20. Электронно-микроскопическая фотография шероховатого эндоплазматического ретикулума с рибосомами на поверхности мемебраны.
Рис. 20. Электронно-микроскопическая фотография шероховатого эндоплазматического ретикулума с рибосомами на поверхности мемебраны.
 секреторную функцию − в гладком ЭР образуются") Функции ЭР. В клетке ЭР выполняет 1) секреторную функцию − в гладком ЭР образуются углеводы, липиды, терпеноиды. − в гранулярном ретикулуме синтезируются мембранные белки, ферменты, необходимые для синтеза полисахаридов клеточных стенок, структурный белок и ферменты клеточных стенок, другие секретируемые белки (рис. 21); 2) транспортную функцию. − по системе ЭР переносятся вещества внутри клетки; 3) коммуникативную функцию − участвует в межклеточных взаимодействиях у растений через плазмодесмы. 4) в мембранах ЭР локализованы редоксцепи двух типов, с участием которых происходят детоксикация вредных для клетки соединений (NADPH-редуктаза и цитохром Р-450) и превращение насыщенных жирных кислот в ненасыщенные (NADH-редуктаза и цитохром b 5); 5) связующая (регуляторная).
Функции ЭР. В клетке ЭР выполняет 1) секреторную функцию − в гладком ЭР образуются углеводы, липиды, терпеноиды. − в гранулярном ретикулуме синтезируются мембранные белки, ферменты, необходимые для синтеза полисахаридов клеточных стенок, структурный белок и ферменты клеточных стенок, другие секретируемые белки (рис. 21); 2) транспортную функцию. − по системе ЭР переносятся вещества внутри клетки; 3) коммуникативную функцию − участвует в межклеточных взаимодействиях у растений через плазмодесмы. 4) в мембранах ЭР локализованы редоксцепи двух типов, с участием которых происходят детоксикация вредных для клетки соединений (NADPH-редуктаза и цитохром Р-450) и превращение насыщенных жирных кислот в ненасыщенные (NADH-редуктаза и цитохром b 5); 5) связующая (регуляторная).
 Рис. 21. Изоляция синтезируемого полипептида в цистернах шероховатого эндоплазматического ретикулума: 1 – мембрана ЭР; 2 – рибосома; 3 – и. РНК; 4 – полипептид.
Рис. 21. Изоляция синтезируемого полипептида в цистернах шероховатого эндоплазматического ретикулума: 1 – мембрана ЭР; 2 – рибосома; 3 – и. РНК; 4 – полипептид.
") 2. 4. 8 Аппарат Гольджи (АГ)
2. 4. 8 Аппарат Гольджи (АГ)
 Количество в клетке от нескольких до сотен АГ. Структура. В растительных клетках АГ представлен диктиосомами, везикулами и межцистерными образованиями (рис. 22). Уплощенные цистерны — диктиосомы расположены пачками по несколько штук. Они ограничены мембраной толщиной 7 — 8 нм. На регенерационном полюсе АГ происходит новообразование диктиосом из мембран гладкого ЭР. На секреторном полюсе формируются секреторные пузырьки (везикулы), содержащие предназначенные для секреции вещества.
Количество в клетке от нескольких до сотен АГ. Структура. В растительных клетках АГ представлен диктиосомами, везикулами и межцистерными образованиями (рис. 22). Уплощенные цистерны — диктиосомы расположены пачками по несколько штук. Они ограничены мембраной толщиной 7 — 8 нм. На регенерационном полюсе АГ происходит новообразование диктиосом из мембран гладкого ЭР. На секреторном полюсе формируются секреторные пузырьки (везикулы), содержащие предназначенные для секреции вещества.
 Везикулы Диктиосомы Рис. 22. Схема строения комплекса Гольджи
Везикулы Диктиосомы Рис. 22. Схема строения комплекса Гольджи
 секреторная − в диктиосомах АГ образуются гликопротеины и гликолипиды; 2) накопительная") Функции АГ: 1) секреторная − в диктиосомах АГ образуются гликопротеины и гликолипиды; 2) накопительная − в диктиосомах АГ осуществляется накопление и мембранная «упаковка» соединений, необходимых для синтеза полимеров клеточной стенки и различных растительных слизей; 3) транспортная − с помощью везикул АГ углеводные компоненты доставляются к плазмалемме. 4) структурная − мембрана пузырьков встраивается в плазмалемму, способствуя ее росту и обновлению, а секретируемые вещества оказываются в клеточной стенке; 5) связующая (регуляторная) − мембраны АГ являются связующим звеном между мембранами ЭР и плазмалеммой.
Функции АГ: 1) секреторная − в диктиосомах АГ образуются гликопротеины и гликолипиды; 2) накопительная − в диктиосомах АГ осуществляется накопление и мембранная «упаковка» соединений, необходимых для синтеза полимеров клеточной стенки и различных растительных слизей; 3) транспортная − с помощью везикул АГ углеводные компоненты доставляются к плазмалемме. 4) структурная − мембрана пузырьков встраивается в плазмалемму, способствуя ее росту и обновлению, а секретируемые вещества оказываются в клеточной стенке; 5) связующая (регуляторная) − мембраны АГ являются связующим звеном между мембранами ЭР и плазмалеммой.
 Комплекс Гольджи − это стопка мембранных пузырьков, одни прилепляются снизу, другие при этом отделяются сверху стопки. Пока содержимое пузырька путешествует снизу вверх, оно подвергается различным химическим модификациям (анимация "The Inner Life of the Cell" ).
Комплекс Гольджи − это стопка мембранных пузырьков, одни прилепляются снизу, другие при этом отделяются сверху стопки. Пока содержимое пузырька путешествует снизу вверх, оно подвергается различным химическим модификациям (анимация "The Inner Life of the Cell" ).
 Отделившаяся от комплекса Гольджи вакуоль изнутри подтаскивается к поверхности клетки (анимация "The Inner Life of the Cell" ).
Отделившаяся от комплекса Гольджи вакуоль изнутри подтаскивается к поверхности клетки (анимация "The Inner Life of the Cell" ).
, и то, что было внутри нее, «выплевывается» наружу.") Вакуоль сливается с мембраной (не показано), и то, что было внутри нее, «выплевывается» наружу. Часть молекул могут свободно дрейфовать по своим делам, но некоторые, оказываются закрепленными внутри вакуолии Теперь они оказались прикрепленными к мембране клетки снаружи (анимация "The Inner Life of the Cell" ).
Вакуоль сливается с мембраной (не показано), и то, что было внутри нее, «выплевывается» наружу. Часть молекул могут свободно дрейфовать по своим делам, но некоторые, оказываются закрепленными внутри вакуолии Теперь они оказались прикрепленными к мембране клетки снаружи (анимация "The Inner Life of the Cell" ).
 То, что нависает сверху - это другая клетка (анимация ("The Inner Life of the Cell" ).
То, что нависает сверху - это другая клетка (анимация ("The Inner Life of the Cell" ).
 2. 4. 9 Рибосомы Форма − округлые частицы. Размеры. Диаметр − 20 -30 нм. Локализация в клетке. Рибосомы в цитоплазме могут быть − свободными, − прикрепленными к мембранам ЭР, к наружной мембране ядерной оболочки либо образуют полирибосомные (полисомные) комплексы. Полирибосомы разрушаются при воздействии на растения неблагоприятных факторов внешней среды (например, засухи, недостатка кислорода).
2. 4. 9 Рибосомы Форма − округлые частицы. Размеры. Диаметр − 20 -30 нм. Локализация в клетке. Рибосомы в цитоплазме могут быть − свободными, − прикрепленными к мембранам ЭР, к наружной мембране ядерной оболочки либо образуют полирибосомные (полисомные) комплексы. Полирибосомы разрушаются при воздействии на растения неблагоприятных факторов внешней среды (например, засухи, недостатка кислорода).
: − в цитоплазме") Строение рибосом. Каждая рибосома состоит из двух нуклеопротеиновых субъединиц (рис. 23): − в цитоплазме растительных клеток находятся 80 S рибосомы, состоящие из 40 и 60 S субъединиц, − в хлоропластах — 70 S рибосомы, − в митохондриях — 78 — 80 S рибосомы. Субъединицы рибосом, образованные в ядрышке, поступают в цитоплазму, где происходит сборка рибосом на молекуле м. РНК. Функции рибосом. Рибосомы осуществляют синтез белков трансляцию матричной, или информационной, РНК (м. РНК).
Строение рибосом. Каждая рибосома состоит из двух нуклеопротеиновых субъединиц (рис. 23): − в цитоплазме растительных клеток находятся 80 S рибосомы, состоящие из 40 и 60 S субъединиц, − в хлоропластах — 70 S рибосомы, − в митохондриях — 78 — 80 S рибосомы. Субъединицы рибосом, образованные в ядрышке, поступают в цитоплазму, где происходит сборка рибосом на молекуле м. РНК. Функции рибосом. Рибосомы осуществляют синтез белков трансляцию матричной, или информационной, РНК (м. РНК).
 Рис. 23. Строение рибосом: 1 − малая субъединица, 2 − большая субъединица, 3 − функционально активная рибосома, состоящая из двух субъединц.
Рис. 23. Строение рибосом: 1 − малая субъединица, 2 − большая субъединица, 3 − функционально активная рибосома, состоящая из двух субъединц.
 и малой (слева). Обе они состоят") Рибосома состоит из двух субчастиц — большой (справа) и малой (слева). Обе они состоят из длинных цепочек РНК (зеленые) и белков (красные). В процессе синтеза новых белков две субчастицы рибосомы соединяются, охватывая при этом цепочку матричной РНК. Перемещая эту РНК на три нуклеотида за шаг, рибосома осуществляет синтез белка. Соединение аминокислот друг с другом протекает при непосредственном участии рибосомальных РНК (р. РНК), которые являются катализаторами реакции. Рисунок с сайта "Molecule of the Month"
Рибосома состоит из двух субчастиц — большой (справа) и малой (слева). Обе они состоят из длинных цепочек РНК (зеленые) и белков (красные). В процессе синтеза новых белков две субчастицы рибосомы соединяются, охватывая при этом цепочку матричной РНК. Перемещая эту РНК на три нуклеотида за шаг, рибосома осуществляет синтез белка. Соединение аминокислот друг с другом протекает при непосредственном участии рибосомальных РНК (р. РНК), которые являются катализаторами реакции. Рисунок с сайта "Molecule of the Month"
 Макромолекулы в цитоплазме клетки: Синим цветом показаны РНК; красным - белки; зеленым - рибосомы. Относительные размеры компонентов РНК, рибосом и белков соблюдены.
Макромолекулы в цитоплазме клетки: Синим цветом показаны РНК; красным - белки; зеленым - рибосомы. Относительные размеры компонентов РНК, рибосом и белков соблюдены.
 Микротрубочки Локализованы в наружном кортикальном слое цитоплазмы неделящихся") 2. 5 Цитоскелет (микротрубочки и микрофиламенты) Микротрубочки Локализованы в наружном кортикальном слое цитоплазмы неделящихся растительных клеток. Размеры: наружный диаметр 30 нм, внутренний — около 14 нм. Они ориентированы параллельно другу и перпендикулярно продольной оси клетки (рис. 24). Строение. Состоят из глобулярного кислого белка тубулина, субъединицей которого является димер, состоящий из двух глобулярных мономеров. В микротрубочке димеры белка располагаются по спирали. Трубка образована 13 субъединицами тубулина. Цитоплазматические микротрубочки легко диссоциируют на субъединицы (разбираются) и собираются вновь. Функции. При делении клетки микротрубочки составляют основу структуры веретена, пучки трубочек прикрепляются также к кинетохорам хромосом. У монадных водорослей микротрубочки входят в состав жгутиков, обеспечивая их подвижность.
2. 5 Цитоскелет (микротрубочки и микрофиламенты) Микротрубочки Локализованы в наружном кортикальном слое цитоплазмы неделящихся растительных клеток. Размеры: наружный диаметр 30 нм, внутренний — около 14 нм. Они ориентированы параллельно другу и перпендикулярно продольной оси клетки (рис. 24). Строение. Состоят из глобулярного кислого белка тубулина, субъединицей которого является димер, состоящий из двух глобулярных мономеров. В микротрубочке димеры белка располагаются по спирали. Трубка образована 13 субъединицами тубулина. Цитоплазматические микротрубочки легко диссоциируют на субъединицы (разбираются) и собираются вновь. Функции. При делении клетки микротрубочки составляют основу структуры веретена, пучки трубочек прикрепляются также к кинетохорам хромосом. У монадных водорослей микротрубочки входят в состав жгутиков, обеспечивая их подвижность.
 Рис. 24. Схема организации цитоскелета: 1 – микрофиламенты; 3 – микротрубочки; 4 – эндоплазматический ретикулум; 5 – цитоплазматическая мембрана; митохондрия; 7 – микротрабекулы
Рис. 24. Схема организации цитоскелета: 1 – микрофиламенты; 3 – микротрубочки; 4 – эндоплазматический ретикулум; 5 – цитоплазматическая мембрана; митохондрия; 7 – микротрабекулы
 Микрофиламенты. Состоят из немышечного актина − сократительного белка, близкого по молекулярной массе аминокислотному составу с актином мышц. Он может находиться в мономерной (глобулярный, Г-актин) или в полимерной форме двойной спирали (фибриллярный, Ф-актин). Функции. Микрофиламенты актина взаимодействуют с микротрубочками кортикального слоя и плазмалеммой. Они участвуют в пространственной организации метаболических процессов, протекающих в растворимой фазе цитоплазмы, и служат основой ее двигательной активности.
Микрофиламенты. Состоят из немышечного актина − сократительного белка, близкого по молекулярной массе аминокислотному составу с актином мышц. Он может находиться в мономерной (глобулярный, Г-актин) или в полимерной форме двойной спирали (фибриллярный, Ф-актин). Функции. Микрофиламенты актина взаимодействуют с микротрубочками кортикального слоя и плазмалеммой. Они участвуют в пространственной организации метаболических процессов, протекающих в растворимой фазе цитоплазмы, и служат основой ее двигательной активности.
 Рис. 25. Строение микротрубочек
Рис. 25. Строение микротрубочек
 Показаны разные элементы цитоскелета, образующие внутреннюю архитектуру клетки (анимация "The Inner Life of the Cell" ). Микротрубочки цитоскелета (анимация "The Inner Life of the Cell" )
Показаны разные элементы цитоскелета, образующие внутреннюю архитектуру клетки (анимация "The Inner Life of the Cell" ). Микротрубочки цитоскелета (анимация "The Inner Life of the Cell" )
 3. Мембранный принцип организации поверхности протоплазмы и органоидов растительной клетки. Биохимическая и функциональная разнокачественность мембран. Основные функции мембран.
3. Мембранный принцип организации поверхности протоплазмы и органоидов растительной клетки. Биохимическая и функциональная разнокачественность мембран. Основные функции мембран.
 Мембранные структурные компоненты клеток одномембранные • плазмалемма; • эндоплазматический ретикулум; • комплекс Гольджи; • лизосомы; • вакуоль двумембранные • ядро; • митохондрии; • пластиды
Мембранные структурные компоненты клеток одномембранные • плазмалемма; • эндоплазматический ретикулум; • комплекс Гольджи; • лизосомы; • вакуоль двумембранные • ядро; • митохондрии; • пластиды
 3. 3 Функции биологических мембран Первичным назначением клеточной мембраны было отделение внутренней среды от внешней. В процессе эволюции в связи с возникновением большого количества специализированных внутриклеточных отсеков (компартментов), различающихся физико-химической средой и характером метаболических процессов, мембраны стали выполнять ряд специфических функций, позволивших осуществлять на разных сторонах мембраны разнообразные, иногда противоположно направленные биохимические реакции.
3. 3 Функции биологических мембран Первичным назначением клеточной мембраны было отделение внутренней среды от внешней. В процессе эволюции в связи с возникновением большого количества специализированных внутриклеточных отсеков (компартментов), различающихся физико-химической средой и характером метаболических процессов, мембраны стали выполнять ряд специфических функций, позволивших осуществлять на разных сторонах мембраны разнообразные, иногда противоположно направленные биохимические реакции.
 барьерные, 2) транспортные, 3) осмотические, 4) электрические, 5) структурные,") Основные функции биологических мембран: 1) барьерные, 2) транспортные, 3) осмотические, 4) электрические, 5) структурные, 6) энергетические, 7) биосинтетические, 8) секреторные, 9) пищеварительные, 10) рецепторно-регуляторные и другие. Далее самостоятельно описать функции мембран
Основные функции биологических мембран: 1) барьерные, 2) транспортные, 3) осмотические, 4) электрические, 5) структурные, 6) энергетические, 7) биосинтетические, 8) секреторные, 9) пищеварительные, 10) рецепторно-регуляторные и другие. Далее самостоятельно описать функции мембран
 Взаимодействие мембранных систем клетки: 1 – ядро; 2 – ядерная пора; 3 – рибосома; 4 – экспортный белок; 5 – белок, используемый на нужды клетки; 6 – продукты секреции (белки, полисахариды и др); 7 – секреторные пузырьки; 8 – комплекс Гольджи; 9 – гладкий ЭР; 10 – шероховатый ЭР.
Взаимодействие мембранных систем клетки: 1 – ядро; 2 – ядерная пора; 3 – рибосома; 4 – экспортный белок; 5 – белок, используемый на нужды клетки; 6 – продукты секреции (белки, полисахариды и др); 7 – секреторные пузырьки; 8 – комплекс Гольджи; 9 – гладкий ЭР; 10 – шероховатый ЭР.