лекция 5 скел мышца.ppt
- Количество слайдов: 43


 01 октября 2014 г. МЫШЕЧНЫЕ")
УО БГМУ КАФЕДРА НОРМАЛЬНОЙ ФИЗИОЛОГИИ Лекция № 3 (5) 01 октября 2014 г. МЫШЕЧНЫЕ ТКАНИ. Лектор : Переверзев Владимир Алексеевич
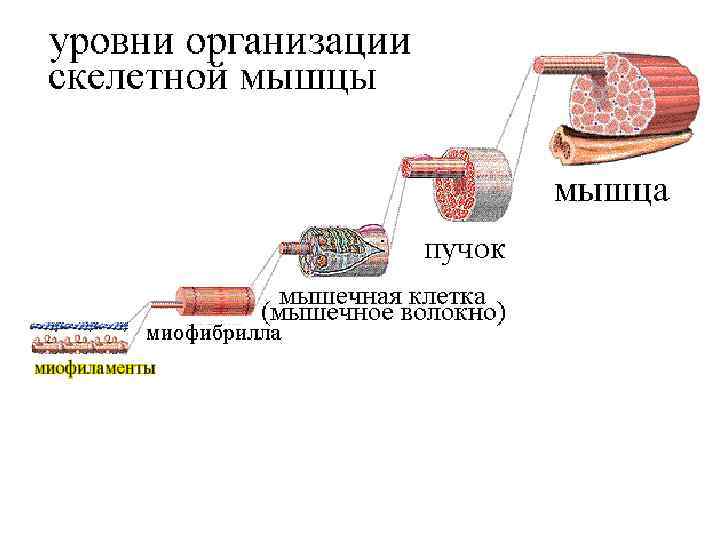

• Скелетные мышцы – активная часть опорнодвигательного аппарата, включающего кости, связки, сухожилия и их сочленения, а с функциональной точки зрения и мотонейроны, вызывающие возбуждение мышечных волокон. • Мотонейрон вместе с иннервируемыми им мышечными волокнами называют нейромоторной (или двигательной) единицей. • Например, в глазных мышцах одна ДЕ содержит 13 -20 мышечных волокон, в мышцах туловища –сотни волокон.

Функции скелетных мышц • Поддержание позы и положения тела в простр-ве • Передвижение тела и его частей в пространстве • Перемещение частей тела относительно друга • Осуществление дыхательных движений • «Периферические сердца» движение крови по сосудам • Участие в выработке тепла • Депонирование питательных веществ
 – обладают малой силой, но")
Типы двигательных единиц - Медленные, неутомляемые (имеют красный цвет) – обладают малой силой, но могут быть длительно в состоянии тонического сокращения без признаков утомления – Тип 1. - Быстрые, легкоутомляемые (имеют белый цвет) – их волокна обладают большой силой сокращения – Тип 2 Б - Быстрые, устойчивые к утомлению – тип 2 Б – имеют относительно большую силу сокращения, в них медленно развивается утомление ! Утомление – временное снижение работоспособности

Свойства скелетных мышц - Возбудимость Проводимость Сократимость Эластичность

Особенности возбудимости - ПП -90 м. В - Амплитуда 130 м. В - Длительность ПД 3 -5 мсек

Особенности проводимости - Скорость проведения возбуждения 3 -5 м в секунду

Особенности сократимости - Изменение длины - Изменение напряжения
")
Режимы сокращений - Изотонический –мышца укорачивается при отсутствии возрастания ее напряжения (только изолированная мышца) - Изометрический – напряжение мышцы возрастает, а длина практически не уменьшается (при попытке поднять непосильный груз) - Ауксотонический - мышца укорачивается и возрастает ее напряжение (трудовая деятельность человека)

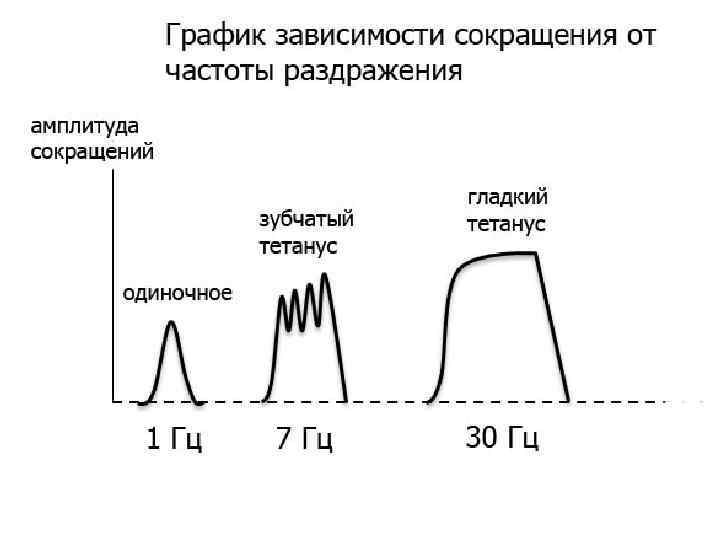
Виды сокращения • Одиночное • Тетаническое - зубчатый - гладкий • Одиночное – проявляется в результате развития одиночной волны возбуждения в мышечных волокнах • Тетаническое – сокращение мышцы, появляющееся в результате возникновения в моторных единицах многочисленных волн возбуждения, сократительный эффект от которых суммируется по амплитуде и времени.
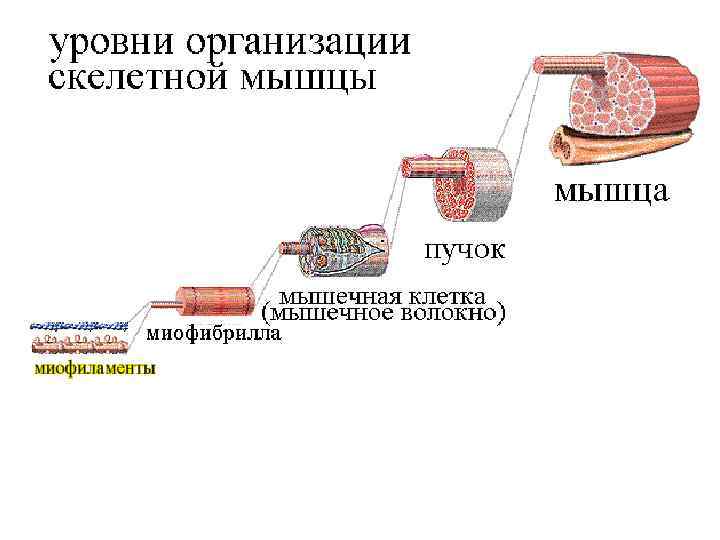

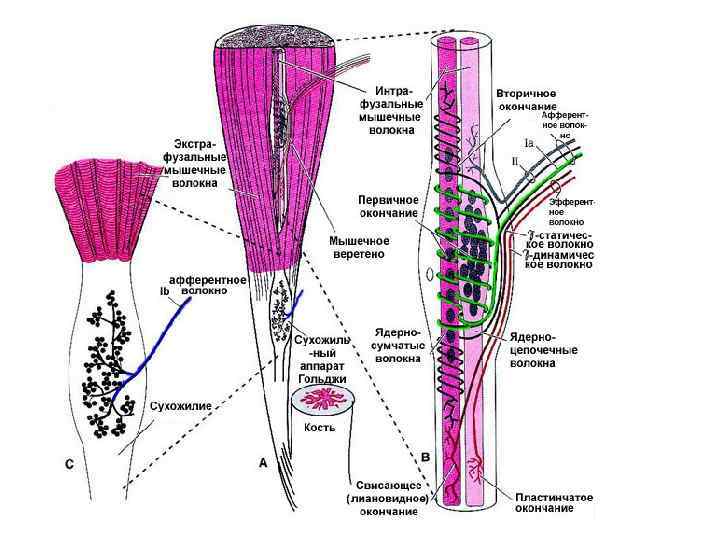
Структура мышечного волокна - Одно мышечное волокно диаметр 0, 1 мм - Длина до 12 см - Длина саркомера 2 мкм - Длина сокращ. саркомера 1 мкм - Мышечные волокна обильно кровоснабжаются - Мышечное волокно бывают ончание (интрафузальное мышечное волокно) и моторную бляшку, через которую передается импульс к сокращению мышцы (экстафузальное мышечное волокно)
 •")
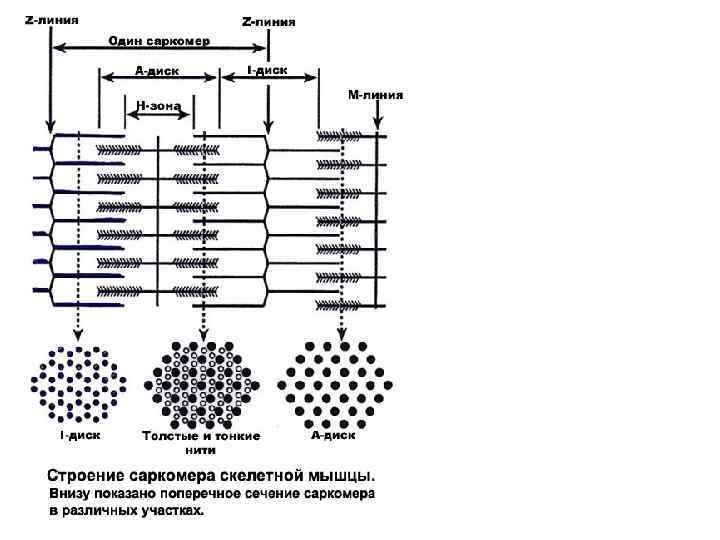
Саркомер • Под световым микроскопом видно чередование светлых и темных полос (поперечная исчерченность) • Темные диски обладают двойным лучепреломлением – анизотропные диски – А • Светлые диски этим свойством не обладают – изотропные диски – I • Часть мышечного волокна от середины одного I диска до середины другого называется саркомером

Сарколемма • Мембрана мышечного волокна образована типичной плазматической мембраной, укрепленной соединительнотканными волокнами. Эти волокна, объединяясь у концов мышечных волокон, образуют сухожилия, с помощью которых мышца крепится к костям.

Саркоплазма • СПР депо ионов Са • СПР – это сеть цистерн и трубочек, ограниченных двухслойными белково-липидными мембранами • Триада • Миоглобин
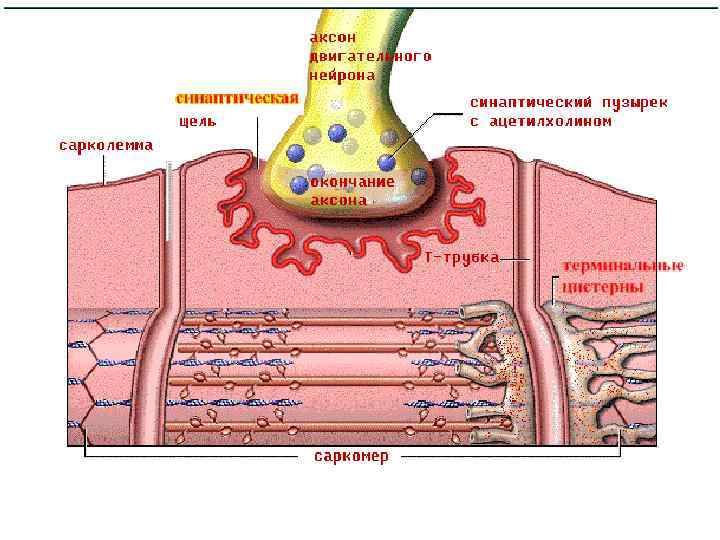
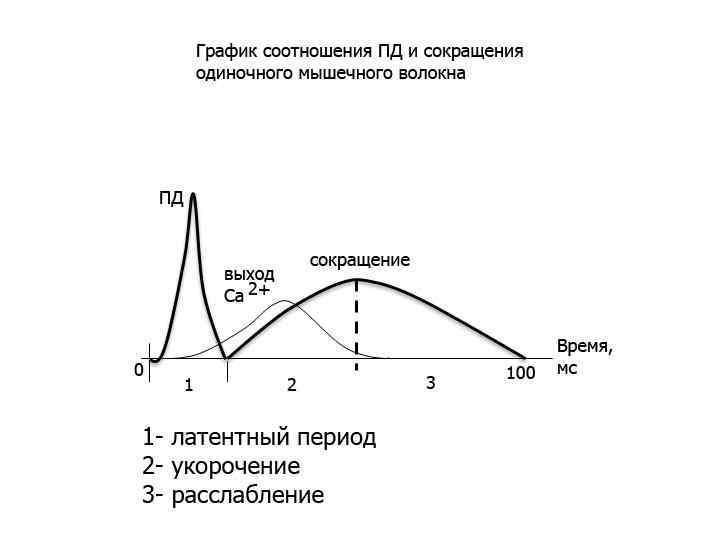
 (2) (3) Вход Na+ внутрь клетки – деполяризация – ПКП, ПКПпотенциал")
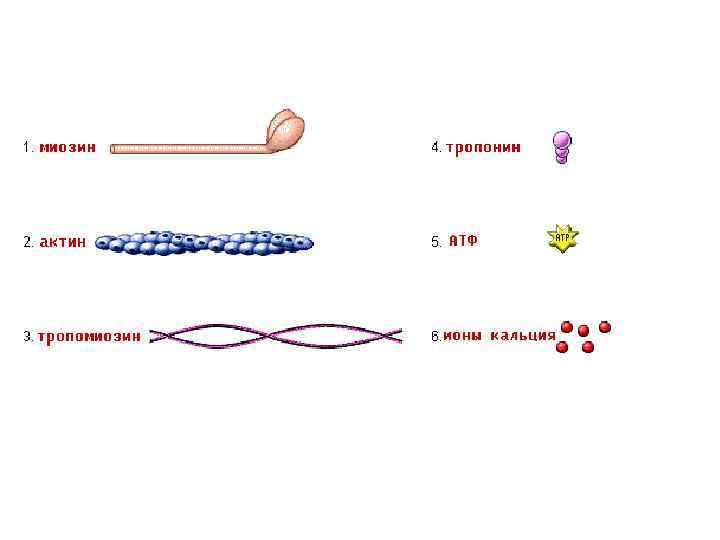
Электромеханическое сопряжение (1) (2) (3) Вход Na+ внутрь клетки – деполяризация – ПКП, ПКПпотенциал концевой пластинки, вид локального потенциала, подчиняется закону силы, по амплитуде может перезаряжать мембрану до -10 мв, суммируется. Электрогенно, за счет локальных токов, происходит формирование ПД на мембране рядом с постсинаптической (электровозбудимая часть мембраны), затем ПД распространяется по система Т-трубочек и достигает СПР В СПР имеются потенциалзависимые каналы (рианодиновые рецепторы) для Са 2+, Са 2+ выходит в цитоплазму по градиенту концентрации (в цитоплазме 10 -8), (в СПР 10 -4) и связывается там с Ссубъединицей тропонина.


Теория скользящих нитей • При повороте головки миозина тянут за собой актиновые нити, продвигая их между миозиновыми. За одно грибковое движение головка может продвинуть актиновую нить на 1% от длины саркомера. Для максимального сокращения нужны повторные грибковые движения головок. Это имеет место при достаточной концентрации АТФ и ионов Са 2+ в саркоплазме.

Расслабление мышечного волокна • Концентрация Са 2+ стала 10 -7 • Работа кальциевого насоса (переход кальция из саркоплазмы в СПР) • Разрыв мостиков между актином и миозином (АТФ и ее связью с молекулами миозина) • После отсоединения эластические силы растягивают саркомер и перемещают нити актина в исходное состояние. • Эластические силы формируются за счет: 1) Эластической тяги спиралевидных клеточных белков, входящих в структуру саркомера 2) Эластических свойств мембран СПР и сарколеммы 3) Эластичности соединительной ткани мышцы, сухожилий и действия сил гравитации

Гладкая мышца

Строение ГМК
 висцеральные (унитарные) гладкие мультиунитарные")
Морфо-функциональная классификация гладких мышц (по плотности иннервации, природе их активности) висцеральные (унитарные) гладкие мультиунитарные гладкие мышцы: во всех внутренних органах (ресничная мышца и мышцы радужки не (ЖКТ, коже, бронхах, сосудах); обладают спонтанной активностью, их спонтанной активностью (автоматией), активность часто имеет нейрогенную миогенная природа мышечного тонуса; природу; мышечные клетки хорошо двигательные нервные окончания на иннервированы вегетативными нервами. небольшом количестве мышечных клеток.

Гладкие мышцы Свойства гладких мышц: - возбудимость - проводимость - сократимость - пластичность (способность длительное время сохранять приданную им за счет растяжения длину без увеличения напряжения. )

Передача информации с нервного волокна на гладкую мышцу. • Передача информации с нервного волокна на гладкую мышцу осуществляется через нейроэффекторное соединение с помощью нейромедиаторов, основными из которых являются ацетилхолин и норадреналин. Они действуют на гладкие миоциты через семисегментные рецепторы, которые стимулируют образование или выделение в цитоплазму вторичных посредников. Иннервированы гладкие мышцы автономной нервной системой.
 - Автоматия - Длительность")
Особенности возбудимости - ПП (Е 0= /-30/ -- /-70/м. В) - Автоматия - Длительность ПД десятки миллисекунд - ПД развивается преимущественно за счет входа Са 2+
")
Особенности проведения возбуждения - Скорость проведения возбуждения 2 -10 см/c - Электрические синапсы (коннексоны) - Свойства функционального синцития
 - Длительность одиночного сокращения до")
Особенности сократимости - Латентный период (0, 251, 00 с) - Длительность одиночного сокращения до 1 мин - Малая сила сокращения - Способность находиться в тоническом сокращении без развития утомления - Расход энергии на сокращение в 100 -500 раз меньше, чем скелетной м. - Могут находиться в состоянии тонического сокращения всю жизнь • Нити актина и миозина располагаются менее упорядоченно чем в скелетной • Нет белка тропонина. Его функцию выполняет белок – кальдесмон, активируемый комплексом 4 Са 2+-кальмодулин • Необходимо расщепления АТФ и перенос фосфата на головки миозина
 -Автоматия -Нервные импульсы (ПД) -БАВ (биологические активные вещества)")
Условия сокращения Многочисленные раздражители: (высокая реактивность) -Автоматия -Нервные импульсы (ПД) -БАВ (биологические активные вещества) -Гормоны -Нейромедиаторы -Простагландины -Метаболиты -Физические факторы (растяжение)
")
Ключевой момент сокращения - Фосфорилирование головок миозина за счет киназы легких цепей миозина (КЛЦМ) - Дефосфорилирование за счет фосфатазы легких цепей миозина (ФЛЦМ)
 Межклеточное пространство 2) СПР (саркоплазматичес кий ретикулум) 3) кавеолы")
Источники Са 2+ 1) Межклеточное пространство 2) СПР (саркоплазматичес кий ретикулум) 3) кавеолы

Последовательность процессов, приводящих к сокращению ГМК • - вход Са 2+ в саркоплазму • Активация кальмодулина • Образование комплекса 4 Са + кальмодулин • Активация кальдесмона и КЛЦМ • Освобождение активных центров на актине и фосфорилирование головок миозина • Связывание головок миозина с актином и поворот головок, при котором нити актина втягиваются между нитями миозина
 Са 2+ - Распад комплекса")
Условия для расслабления - Снижение (до 10 -7 ) Са 2+ - Распад комплекса 4 Са 2+ кальмодулин - Силы упругости восстанавливают исходную длину ГМК
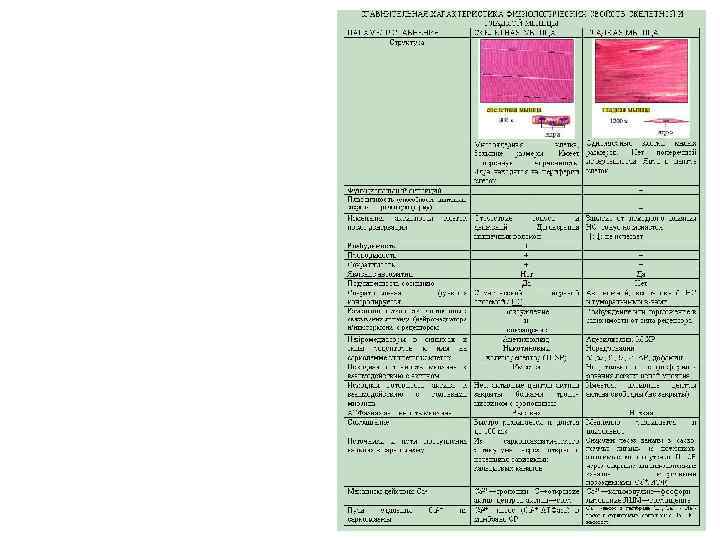

Отличия нейроэффекторных соединений гладких мышц от нейромышечных синапсов скелетных мышц ПАРАМЕТР СРАВНЕНИЯ СКЕЛЕТНАЯ МЫШЦА ГЛАДКАЯ МЫШЦА Иннервация Соматической Автономной нервной системой Нейромедиаторы в синапсах и типы Ацетилхолин; М-ХР рецепторов к ним на сарколемме Никотиновый холинорецептор Норадреналин мышечных клеток (Н-ХР) α 1, α 2, β 1, β 2, β 3–АР; дофамин Наличие варикозных расширений на аксоне и выделение из них медиатора Наличие пресинаптической терминали на аксоне и выделение нейромедиатора из нее Ширина синаптической щели Диффузия медиатора из синаптической щели Изменение активности клеток после связывания лиганда (нейромедиатора и/или гормона) с рецептором нет Имеются, многочисленные есть нет Малая (30 нм) большая Практически отсутствует Достаточно выражена Возбуждение и сокращение Возбуждение или торможение (отсутствием возбуждения) в зависимости от типа рецептора

Понятие о миоэпителиальных клетках и их функциях • Название этих клеток связано с тем, что они, с одной стороны, имеют эпителиальное происхождение, а с другой стороны, обладают сократительной активностью. Миоэпителиальные клетки находятся в экзокринных железах. Они окружают клетки секреторных отделов (экзокриноциты) и выводных протоков (эпителиоциты) слюнных, слезных, потовых, молочных желез, прикрепляясь при помощи полудесмосом к базальной мембране. • Миоэпителиальные клетки сильно уплощены, имеют звездчатую форму и своими отростками охватывают экзокриноциты и эпителиоциты. Сократительный аппарат миоэпителиальных клеток представлен (как и в гладких мышцах), готовыми к взаимодействию с миозином актиновыми миофиламентами, прикрепленными через αактинин к плотным тельцам, и миозиновыми толстыми нитями, структура которых зависит от сокращения и расслабления.

• • • Механизм сокращения и расслабления миоэпителиоцитов напоминает таковой у гладких миоцитов. Нейромедиатор (ацетилхолин, норадреналин) или гормон (окситоцин, адреналин), связавшись со специфическим рецептором, через свой рецептор активирует диссоциацию G– белка, α-субъединица которого далее активирует фосфолипазу С. Фосфолипаза С катализирует образование инозитолтрифосфата (ИТФ 3) и диацилглицерола из фосфоинозитолдифосфата мембраны клетки. ИТФ 3 диффундирует к эндоплазматическому ретикулуму и после взаимодействия со своими рецепторами вызывает открытие кальциевых каналов и высвобождение Са 2+ из депо в цитоплазму. Увеличение содержания Са 2+ в цитоплазме является ключевым событием для инициации сокращения миоэпителиоцита. Са 2+ связывается с кальмодулином, полученный комплекс активирует фермент - киназу легких цепей миозина, необходимую для фосфорилирования легких цепей и сборки толстой нити из молекул миозина. Далее происходит прикрепление к актину фосфорилированных головок миозина и их поворот, при котором нити актина втягиваются между нитями миозина – совершается одно гребковое движение. В результате гидролиза АТФ актин-миозиновые связи разрушаются, головки миозина восстанавливают свою конформацию и приобретают готовность к новому гребковому движению. Продолжающаяся стимуляция рецепторов миоэпителиоцита гормоном или нейромедиатором и высокая концентрация Са 2+ в цитоплазме поддерживают дальнейшее сокращение клетки. Таким образом, сила и продолжительность сокращения миоэпителиальных клеток (как и гладких миоцитов и мышечных волокон скелетных мышц) определяется концентрацией свободного Са 2+, окружающего миофиламенты. Расслабление миоэпителиальных клеток наступает при снижении содержания Са 2+ в цитоплазме менее 10 -7 М/л, что приводит к распаду комплекса Са 2+ кальмодулин, дефосфорилированию головок миозина, разрыву связей между актином и миозином. Сокращение миоэпителиальных клеток облегчает выделение секрета из концевых отделов и его продвижение по выводным протокам экзокринных желез. Ацетилхолин стимулирует сокращение миоэпителиальных клеток слезных и потовых желез, норадреналин - слюнных желез, окситоцин - молочных желез при лактации.
лекция 5 скел мышца.ppt