

БХ УГЛЕВОДЫ.pptx
- Количество слайдов: 116
 УГЛЕВОДЫ Номенклатура и классификация Моносахариды Изомерия и стереохимия Химические свойства и реакции Олигосахариды Полисахариды Производные (С Н 2 О)n
УГЛЕВОДЫ Номенклатура и классификация Моносахариды Изомерия и стереохимия Химические свойства и реакции Олигосахариды Полисахариды Производные (С Н 2 О)n
 х 1 Полисахариды (гликаны) >10 Гликоконъюгаты: Альдозы") Номенклатура Олигосахариды х 2 -10 Моносахариды (простые) х 1 Полисахариды (гликаны) >10 Гликоконъюгаты: Альдозы Дисахариды Гомо- Гликопротеиды Кетозы Более крупные олигосахариды Гетеро- Протеогликаны ✓ Триозы глицеральдегид дигидроксиацетон ✓ Тетрозы ✓ Пентозы рибоза ✓ Гексозы глюкоза фруктоза сахароза лактоза мальтоза изомальтоза трегалоза ✓ Резервные крахмал гликоген декстран ✓ Структурные целлюлоза агароза хитин гликозаминогликаны Гликолипиды Липополисахариды
Номенклатура Олигосахариды х 2 -10 Моносахариды (простые) х 1 Полисахариды (гликаны) >10 Гликоконъюгаты: Альдозы Дисахариды Гомо- Гликопротеиды Кетозы Более крупные олигосахариды Гетеро- Протеогликаны ✓ Триозы глицеральдегид дигидроксиацетон ✓ Тетрозы ✓ Пентозы рибоза ✓ Гексозы глюкоза фруктоза сахароза лактоза мальтоза изомальтоза трегалоза ✓ Резервные крахмал гликоген декстран ✓ Структурные целлюлоза агароза хитин гликозаминогликаны Гликолипиды Липополисахариды
 Моносахариды Альдозы Ряды альдоз и кетоз Кетозы
Моносахариды Альдозы Ряды альдоз и кетоз Кетозы
 Стереоизомерия Энантиомеры – зеркальные отражения Диастереомеры – не зеркальные, но разные при ≥ 1 хиральном центре Эпимеры – разные только по 1 центру + и - означают оптическое вращение, но не имеют отношения к D и L
Стереоизомерия Энантиомеры – зеркальные отражения Диастереомеры – не зеркальные, но разные при ≥ 1 хиральном центре Эпимеры – разные только по 1 центру + и - означают оптическое вращение, но не имеют отношения к D и L
 Аномерия
Аномерия
 Изображения и конформации Haworth Fisher Кресло Ванна
Изображения и конформации Haworth Fisher Кресло Ванна
 Реакции моносахаридов
Реакции моносахаридов
-Glc (трегалоза)") Дисахариды ✓Редуцирующий и нередуцирующий концы; α и β аномерные формы Glc-(α 1 -1α)-Glc (трегалоза) – криопротектор ?
Дисахариды ✓Редуцирующий и нередуцирующий концы; α и β аномерные формы Glc-(α 1 -1α)-Glc (трегалоза) – криопротектор ?
 Крахмал = Амилоза Амилопектин +") Резервные полисахариды (гликаны) Крахмал = Амилоза Амилопектин +
Резервные полисахариды (гликаны) Крахмал = Амилоза Амилопектин +
 Гликоген") Резервные полисахариды (гликаны) Гликоген
Резервные полисахариды (гликаны) Гликоген
 Структурные полисахариды Растения Насекомые Бурые водоросли
Структурные полисахариды Растения Насекомые Бурые водоросли
 Целлюлоза
Целлюлоза
 Гликозаминогликаны
Гликозаминогликаны
 стафилококков") Пептидогликаны грамм(+) стафилококков
Пептидогликаны грамм(+) стафилококков
 Гликопротеины
Гликопротеины
 Протеогликаны
Протеогликаны
 Протеогликаны Гликокаликс
Протеогликаны Гликокаликс
 бактерий") ЛПС внешней мембраны грамм(-) бактерий
ЛПС внешней мембраны грамм(-) бактерий
 ПИЩЕВАРЕНИЕ УГЛЕВОДОВ Отделы ЖКТ Ферменты Субстраты и расщепляемые связи Транспорт моносахаридов через мембраны Нарушения пищеварения углеводов
ПИЩЕВАРЕНИЕ УГЛЕВОДОВ Отделы ЖКТ Ферменты Субстраты и расщепляемые связи Транспорт моносахаридов через мембраны Нарушения пищеварения углеводов
 Переваривание углеводов ü α-амилаза слюны ü Панкреатическая α-амилаза ü Сахаразо-изомальтазный комплекс ü Гликоамилазный комплекс ü Лактаза, трегалаза
Переваривание углеводов ü α-амилаза слюны ü Панкреатическая α-амилаза ü Сахаразо-изомальтазный комплекс ü Гликоамилазный комплекс ü Лактаза, трегалаза
 Панкреатическая амилаза Гидролизует α-1, 4 -гликозидные связи, не гидролизует α-1, 6 и β-1, 4
Панкреатическая амилаза Гидролизует α-1, 4 -гликозидные связи, не гидролизует α-1, 6 и β-1, 4
 и изомальтозу (α-1,") Сахаразо-изомальтазный комплекс сахараза изомальтаза линкер трансмембранный цитоплазматический Гидролизует сахарозу (α-1, 2) и изомальтозу (α-1, 6), меньше мальтозу (α-1, 4), не гидролизует β-1, 4
Сахаразо-изомальтазный комплекс сахараза изомальтаза линкер трансмембранный цитоплазматический Гидролизует сахарозу (α-1, 2) и изомальтозу (α-1, 6), меньше мальтозу (α-1, 4), не гидролизует β-1, 4
мальтоза (α-1, 4) Лактаза лактоза (β-1, 4) Трегалаза трегалоза (α-1,") Остальные ферменты Гликоамилазный комплекс (олиго)мальтоза (α-1, 4) Лактаза лактоза (β-1, 4) Трегалаза трегалоза (α-1, 1)
Остальные ферменты Гликоамилазный комплекс (олиго)мальтоза (α-1, 4) Лактаза лактоза (β-1, 4) Трегалаза трегалоза (α-1, 1)
 Транспорт моносахаридов через кишечный эпителий
Транспорт моносахаридов через кишечный эпителий
 Транспорт глюкозы из крови в клетки ü ü ü GLUT-1 – транспорт глюкозы в мозг GLUT-2 – основной GLUT-3 – нервная ткань, больше сродство чем у GLUT-1 GLUT-4 – мышцы, адипоциты, зависит от инсулина GLUT-5 – тонкий кишечник, наименее изучен
Транспорт глюкозы из крови в клетки ü ü ü GLUT-1 – транспорт глюкозы в мозг GLUT-2 – основной GLUT-3 – нервная ткань, больше сродство чем у GLUT-1 GLUT-4 – мышцы, адипоциты, зависит от инсулина GLUT-5 – тонкий кишечник, наименее изучен
 Механизм регуляции GLUT-4 инсулином
Механизм регуляции GLUT-4 инсулином
 Нарушения переваривания углеводов
Нарушения переваривания углеводов
 СУДЬБА ГЛЮКОЗЫ В КЛЕТКЕ Пути использования Фосфорилирование глюкозы Гексокиназа и глюкокиназа
СУДЬБА ГЛЮКОЗЫ В КЛЕТКЕ Пути использования Фосфорилирование глюкозы Гексокиназа и глюкокиназа
 структура (гликозилирование)") Основные пути утилизации глюкозы в клетке запасание (гликоген, . . . жиры) структура (гликозилирование) энергия (гликолиз) анаболизм (пентозный путь)
Основные пути утилизации глюкозы в клетке запасание (гликоген, . . . жиры) структура (гликозилирование) энергия (гликолиз) анаболизм (пентозный путь)
 Фосфорилирование глюкозы
Фосфорилирование глюкозы
 Гексокиназа и глюкокиназа Ткани : Гексокиназа Печень : глюкоза Глюкокиназа + Гексокиназа + Глюкозо-6 фосфатаза
Гексокиназа и глюкокиназа Ткани : Гексокиназа Печень : глюкоза Глюкокиназа + Гексокиназа + Глюкозо-6 фосфатаза
 ОБМЕН ГЛИКОГЕНА Биологический смысл Реакции и ферменты Энергетический баланс Регуляция
ОБМЕН ГЛИКОГЕНА Биологический смысл Реакции и ферменты Энергетический баланс Регуляция
 Биомедицинское значение Поддержание уровня глюкозы в крови Абсорбтивный период Постабсорбтивный период Локализация в организме Печень 40 % Мышцы 60 % Ключевые участники Глюкозо-6 -фосфат Ветвящие ферменты Гексокиназа Глюкокиназа Гликогенсинтаза Гликогенфосфорилаза
Биомедицинское значение Поддержание уровня глюкозы в крови Абсорбтивный период Постабсорбтивный период Локализация в организме Печень 40 % Мышцы 60 % Ключевые участники Глюкозо-6 -фосфат Ветвящие ферменты Гексокиназа Глюкокиназа Гликогенсинтаза Гликогенфосфорилаза
 Строение гликогена Значение разветвленной структуры
Строение гликогена Значение разветвленной структуры
 Синтез гликогена гексокиназа глюкокиназа фосфоглюкомутаза УДФ-глюкопирофосфорилаза гликогенсинтаза ветвление
Синтез гликогена гексокиназа глюкокиназа фосфоглюкомутаза УДФ-глюкопирофосфорилаза гликогенсинтаза ветвление
 Распад гликогена DG 0’ = +3, 1 к. Дж/моль 100 Фi : 1 глюкозо-1 -Ф DG = -6 к. Дж/моль (Глюкоза)n + H 3 PO 4 (Глюкоза)n-1 + Глю-6 -Ф
Распад гликогена DG 0’ = +3, 1 к. Дж/моль 100 Фi : 1 глюкозо-1 -Ф DG = -6 к. Дж/моль (Глюкоза)n + H 3 PO 4 (Глюкоза)n-1 + Глю-6 -Ф
 Особенности ветвления и деветвления Для дебранчинга нужны две ферментативных активности
Особенности ветвления и деветвления Для дебранчинга нужны две ферментативных активности
") Нарушения обмена гликогена (гликогенозы)
Нарушения обмена гликогена (гликогенозы)
 Регуляция обмена гликогена: физиологическая роль
Регуляция обмена гликогена: физиологическая роль
 Активность гликогенсинтазы и фосфорилазы регулируется фосфорилированием синтез распад
Активность гликогенсинтазы и фосфорилазы регулируется фосфорилированием синтез распад
 Печень - первичная мишень действия инсулина Из поджелудочной железы инсулин сначала попадает в печень через портальную вену, и лишь затем в системную циркуляцию
Печень - первичная мишень действия инсулина Из поджелудочной железы инсулин сначала попадает в печень через портальную вену, и лишь затем в системную циркуляцию
 Механизм действия инсулина на общий метаболизм
Механизм действия инсулина на общий метаболизм
 Глобальная регуляция метаболизма Insulin Growth factors Ins. R RTK PI 3 K NOX Akt IRS 1/2 H 2 O 2 TSC 2 S 6 K 1 m. TORC 1 Autophagy m. TORC 2 Feedback enables: - Insulin resistance - Cell migration - Proliferation Glucose PPARγ SREBP Adipo/Lipido genesis 4 E-BP Proteins (growth)
Глобальная регуляция метаболизма Insulin Growth factors Ins. R RTK PI 3 K NOX Akt IRS 1/2 H 2 O 2 TSC 2 S 6 K 1 m. TORC 1 Autophagy m. TORC 2 Feedback enables: - Insulin resistance - Cell migration - Proliferation Glucose PPARγ SREBP Adipo/Lipido genesis 4 E-BP Proteins (growth)
 Механизм регуляции синтеза гликогена инсулином синтез распад
Механизм регуляции синтеза гликогена инсулином синтез распад
 Регуляция распада гликогена через b 2 -рецепторы синтез распад
Регуляция распада гликогена через b 2 -рецепторы синтез распад
 Действие адреналина через a 1 -рецепторы синтез распад
Действие адреналина через a 1 -рецепторы синтез распад
 Связывание Са 2+ имитирует фосфорилирование синтез гликогена
Связывание Са 2+ имитирует фосфорилирование синтез гликогена
 ГЛИКОЛИЗ Биологический смысл Реакции первой и второй фаз Сопряженность реакций Анаэробный и аэробный Другие углеводные субстраты Энергетический баланс Регуляция
ГЛИКОЛИЗ Биологический смысл Реакции первой и второй фаз Сопряженность реакций Анаэробный и аэробный Другие углеводные субстраты Энергетический баланс Регуляция
 Центральное место в метаболизме
Центральное место в метаболизме
 Общий вид и 2 фазы
Общий вид и 2 фазы
") Первая фаза гликолиза ( ΔG’ ~ 0 )
Первая фаза гликолиза ( ΔG’ ~ 0 )
 Вторая фаза гликолиза
Вторая фаза гликолиза
 ГАФД : кислотно-основный и ковалентный катализ
ГАФД : кислотно-основный и ковалентный катализ
") Энергетика гликолиза и сопряженные реакции Δ=-120 k. J/mol (~4 АТФ)
Энергетика гликолиза и сопряженные реакции Δ=-120 k. J/mol (~4 АТФ)
 Реальная цена гликолиза – КПД ≥ 60%
Реальная цена гликолиза – КПД ≥ 60%
 Аэробный и анаэробный гликолиз
Аэробный и анаэробный гликолиз
 Молочнокислое и спиртовое брожение
Молочнокислое и спиртовое брожение
 Malt
Malt
 Эритроциты и 2, 3 -бисфосфоглицератный цикл
Эритроциты и 2, 3 -бисфосфоглицератный цикл
 3. Ковалентная") Регуляция гликолиза 1. За счет концентраций субстрата/продукта 2. Метаболитами (АТФ-АДФ, НАДН-НАД+) 3. Ковалентная модификация (гормональная) 4. Аллостерически (2, 6 -БФГ) 5. Изоформный состав (гексокиназа, ЛДГ)
Регуляция гликолиза 1. За счет концентраций субстрата/продукта 2. Метаболитами (АТФ-АДФ, НАДН-НАД+) 3. Ковалентная модификация (гормональная) 4. Аллостерически (2, 6 -БФГ) 5. Изоформный состав (гексокиназа, ЛДГ)
 ГЛЮКОНЕОГЕНЕЗ Биологический смысл Последовательность превращений Энергетический баланс Регуляция
ГЛЮКОНЕОГЕНЕЗ Биологический смысл Последовательность превращений Энергетический баланс Регуляция
 Биомедицинское значение Поддержание уровня глюкозы в крови Абсорбтивный период Постабсорбтивный период Голодание Локализация в организме Печень 90 % Почки 10 % Субстраты глюконеогенеза Пируват Глицерин Интермедиаты ЦТК Ацетил-Ко. А Лактат Аминокислоты аланин лизин лейцин Жиры
Биомедицинское значение Поддержание уровня глюкозы в крови Абсорбтивный период Постабсорбтивный период Голодание Локализация в организме Печень 90 % Почки 10 % Субстраты глюконеогенеза Пируват Глицерин Интермедиаты ЦТК Ацетил-Ко. А Лактат Аминокислоты аланин лизин лейцин Жиры
 Цикл Кори
Цикл Кори
 Глюкозо-аланиновый цикл
Глюкозо-аланиновый цикл
 Пагубные последствия пребывания в Mac. Donald’s Синтез жиров из углеводов Синтез глюкозы из ацетил-Ко. А возможен только теоретически через цикл Кребса, но этого в клетках не происходит
Пагубные последствия пребывания в Mac. Donald’s Синтез жиров из углеводов Синтез глюкозы из ацетил-Ко. А возможен только теоретически через цикл Кребса, но этого в клетках не происходит
 Субстраты глюконеогенеза
Субстраты глюконеогенеза
 Синтез глюкозы из глицерина жировая ткань из триглицеридов только в печени и почках глюкоза
Синтез глюкозы из глицерина жировая ткань из триглицеридов только в печени и почках глюкоза
") Глюконеогенез - частичное обращение гликолиза субстратные циклы (метаболический контроль)
Глюконеогенез - частичное обращение гликолиза субстратные циклы (метаболический контроль)
 Уникальные реакции глюконеогенеза - III цикл 2 макроэргических эквивалента + внутриклеточная локализация и необходимость транспорта Ац-Ко. А
Уникальные реакции глюконеогенеза - III цикл 2 макроэргических эквивалента + внутриклеточная локализация и необходимость транспорта Ац-Ко. А
 Альтернативные пути образования ФЕП
Альтернативные пути образования ФЕП
 Уникальные реакции глюконеогенеза - I и II циклы
Уникальные реакции глюконеогенеза - I и II циклы
 Энергетический баланс глюконеогенеза Сопряжение с гидролизом АТФ/ГТФ “тянет” глюконеогенез: DG 0’ = -37, 7 к. Дж/моль (DG = -15, 6) Простое обращение гликолиза энергетически невозможно: DG 0’ = 74, 0 к. Дж/моль
Энергетический баланс глюконеогенеза Сопряжение с гидролизом АТФ/ГТФ “тянет” глюконеогенез: DG 0’ = -37, 7 к. Дж/моль (DG = -15, 6) Простое обращение гликолиза энергетически невозможно: DG 0’ = 74, 0 к. Дж/моль
 Регуляция глюконеогенеза субстратная регуляция аллостерическая регуляция I субстратная регуляция II аллостерическая регуляция III аллостерическая регуляция
Регуляция глюконеогенеза субстратная регуляция аллостерическая регуляция I субстратная регуляция II аллостерическая регуляция III аллостерическая регуляция
 Регуляция пируваткиназы в III цикле
Регуляция пируваткиназы в III цикле
 цикла PFK-1 FBPase-1") Регуляция II cубстратного (“футильного”) цикла PFK-1 FBPase-1
Регуляция II cубстратного (“футильного”) цикла PFK-1 FBPase-1
 ПЕНТОЗНЫЙ ПУТЬ Биологический смысл Последовательность превращений Энергетический баланс Регуляция
ПЕНТОЗНЫЙ ПУТЬ Биологический смысл Последовательность превращений Энергетический баланс Регуляция
 Суммарная схема ПФП
Суммарная схема ПФП
 Биологическое значение и энергетический баланс Образование восстановительных эквивалентов НАДФН+Н+ Образование рибозы для синтеза пуриновых и перимидиновых оснований Синтез нуклеотидов Метаболизм других моносахаридов Структурные компоненты клетки 6 Глюкозо-6 -фосфат + 12 НАДФ + 12 H 2 O 5 Глюкозо-6 -фосфат + 12 НАДФH + 12 H+ + 6 CO 2
Биологическое значение и энергетический баланс Образование восстановительных эквивалентов НАДФН+Н+ Образование рибозы для синтеза пуриновых и перимидиновых оснований Синтез нуклеотидов Метаболизм других моносахаридов Структурные компоненты клетки 6 Глюкозо-6 -фосфат + 12 НАДФ + 12 H 2 O 5 Глюкозо-6 -фосфат + 12 НАДФH + 12 H+ + 6 CO 2
 Метаболические превращения
Метаболические превращения
 Окислительный этап
Окислительный этап
") Неокислительный этап (пентозный цикл)
Неокислительный этап (пентозный цикл)
 С 7 С 6 С 6 С 5 С 3") Неокислителный этап (пентозный цикл) С 7 С 6 С 6 С 5 С 3 С 4 С 5 С 3
Неокислителный этап (пентозный цикл) С 7 С 6 С 6 С 5 С 3 С 4 С 5 С 3
 ОБЩИЙ ПУТЬ КАТАБОЛИЗМА l 3 стадии катаболизма l Биологический смысл ОПК l ОПК: ЦТК + ДЦ + ОФ
ОБЩИЙ ПУТЬ КАТАБОЛИЗМА l 3 стадии катаболизма l Биологический смысл ОПК l ОПК: ЦТК + ДЦ + ОФ
 3 стадии и место ОПК в общем катаболизме Стадия 1. Общие блоки Стадия 2. Общий продукт Ацетил-Ко. А Стадия 3. Общий Путь Катаболизма (ОПК) Конечные продукты АТФ НАДН
3 стадии и место ОПК в общем катаболизме Стадия 1. Общие блоки Стадия 2. Общий продукт Ацетил-Ко. А Стадия 3. Общий Путь Катаболизма (ОПК) Конечные продукты АТФ НАДН
 ЦТК есть первая стадия ОПК ЦТК ДЦ ОФ
ЦТК есть первая стадия ОПК ЦТК ДЦ ОФ
 ОБРАЗОВАНИЕ Ацетил-Ко. А ИЗ ПИРУВАТА Транспорт пирувата l Устройство и механизм ПДК l Коферменты l Сопряженные реакции l Стехиометрия l Регуляция l
ОБРАЗОВАНИЕ Ацетил-Ко. А ИЗ ПИРУВАТА Транспорт пирувата l Устройство и механизм ПДК l Коферменты l Сопряженные реакции l Стехиометрия l Регуляция l
 Перенос пирувата в митохондрии За счет электрохимического градиента
Перенос пирувата в митохондрии За счет электрохимического градиента
 Механизм окислительного декарбоксилирования пирувата
Механизм окислительного декарбоксилирования пирувата
 Механизм действия тиаминового кофермента Тиазоловое кольцо
Механизм действия тиаминового кофермента Тиазоловое кольцо
") Липоевая кислота и кофермент А Пантотеновая к-та Фосфоаденозин-дифосфат β-меркаптоэтиламин Липоамид Ацетил-Кофермент-А (Ац-Ко. А)
Липоевая кислота и кофермент А Пантотеновая к-та Фосфоаденозин-дифосфат β-меркаптоэтиламин Липоамид Ацетил-Кофермент-А (Ац-Ко. А)
 Тунелирование субстрата Интермедиаты «передаются с рук на руки» и никогда не покидают поверхности ферментов
Тунелирование субстрата Интермедиаты «передаются с рук на руки» и никогда не покидают поверхности ферментов
 Электронная микроскопия ПДК Электронная микрофотография 3 D-image reconstruction
Электронная микроскопия ПДК Электронная микрофотография 3 D-image reconstruction
 Неравная стехиометрия и состав ПДК
Неравная стехиометрия и состав ПДК
, продукты ингибируют (Ац-Ко. А, НАДН).") Регуляция пируватдегидрогеназы Регуляция метаболитами: субстраты активируют (пируват, НАД+, АТФ), продукты ингибируют (Ац-Ко. А, НАДН). Регуляция идет через киназу ПДК
Регуляция пируватдегидрогеназы Регуляция метаболитами: субстраты активируют (пируват, НАД+, АТФ), продукты ингибируют (Ац-Ко. А, НАДН). Регуляция идет через киназу ПДК
 l История открытия l Последовательность событий l Энергетика l Термодинамика l") ЦИКЛ КРЕБСА (ЦТК) l История открытия l Последовательность событий l Энергетика l Термодинамика l Ключевые механизмы l Регуляция
ЦИКЛ КРЕБСА (ЦТК) l История открытия l Последовательность событий l Энергетика l Термодинамика l Ключевые механизмы l Регуляция
 История открытия 1932. Hans Krebs. 1935. Albert Szent-Gyorgyi. Только некоторые малые органические кислоты окисляются экстрактами почек и печени. Добавление любой из трех дикарбоновых кислот к экстрактам мышц повышало потребление кислорода существенно больше, чем требовалось только для их окисления. Оксалоацетат образуется из пирувата. сукцинат малат фумарат цитрат ацетат Цитрат > изоцитрат > aкетоглутарат Было известно: a -КГ > сукцинат Лимитирующие вещества. Окисление сукцината скорость-лимитирующая стадия (ингибирование малонатом) оксалоацетат пируват Karl Martius Franz Knoop. сукцинат фумарат малат Последовательно сть реакций определилась
История открытия 1932. Hans Krebs. 1935. Albert Szent-Gyorgyi. Только некоторые малые органические кислоты окисляются экстрактами почек и печени. Добавление любой из трех дикарбоновых кислот к экстрактам мышц повышало потребление кислорода существенно больше, чем требовалось только для их окисления. Оксалоацетат образуется из пирувата. сукцинат малат фумарат цитрат ацетат Цитрат > изоцитрат > aкетоглутарат Было известно: a -КГ > сукцинат Лимитирующие вещества. Окисление сукцината скорость-лимитирующая стадия (ингибирование малонатом) оксалоацетат пируват Karl Martius Franz Knoop. сукцинат фумарат малат Последовательно сть реакций определилась
 История открытия 1937. Hans Krebs. Экстракты мышц образуют цитрат при добавлении оксалоацетата и либо пирувата, либо ацетата. Кребс уже знал о метаболических циклах - в 1932 он открыл цикл мочевины Круг замкнулся
История открытия 1937. Hans Krebs. Экстракты мышц образуют цитрат при добавлении оксалоацетата и либо пирувата, либо ацетата. Кребс уже знал о метаболических циклах - в 1932 он открыл цикл мочевины Круг замкнулся
 Последовательность реакций и энергетика
Последовательность реакций и энергетика
 Еще раз про энергетический баланс
Еще раз про энергетический баланс
 Асимметрия цикла Вводимая карбоксильная группа удаляется на втором цикле
Асимметрия цикла Вводимая карбоксильная группа удаляется на втором цикле
 Термодинамика цикла -31, 4 +6, 3 +29, 7 -8, 4 сопряжение Величины DG 0’, к. Дж/моль самая медленная +3, 8 -30, 1 0 -3, 3
Термодинамика цикла -31, 4 +6, 3 +29, 7 -8, 4 сопряжение Величины DG 0’, к. Дж/моль самая медленная +3, 8 -30, 1 0 -3, 3
 1 2") Механизм цитратсинтазной реакции Последовательная упорядоченная двусубстратная реакция (ordered single displacement) 1 2
Механизм цитратсинтазной реакции Последовательная упорядоченная двусубстратная реакция (ordered single displacement) 1 2
 Аконитазная реакция ΔG' +6, 3 Аконитат гидратаза цистеин Fe. S кластер изоцитрат -8, 4
Аконитазная реакция ΔG' +6, 3 Аконитат гидратаза цистеин Fe. S кластер изоцитрат -8, 4
 Первое окисление: изоцитрат-ДГ Окислительное декарбоксилирование Две формы фермента: НАД+-зависимый – в матриксе митохондрий и НАДФ+-зависимый – в матриксе и цитозоле
Первое окисление: изоцитрат-ДГ Окислительное декарбоксилирование Две формы фермента: НАД+-зависимый – в матриксе митохондрий и НАДФ+-зависимый – в матриксе и цитозоле
 Второе окисление: α-кетоглутарат-ДГ ΔG’= -30, 1 Самая «движущая» реакция
Второе окисление: α-кетоглутарат-ДГ ΔG’= -30, 1 Самая «движущая» реакция
 α-кетоглутарат-ДГ – аналог ПДК Механизм практически идентичен ПДК: комплекс из 3 ферментов и тех же 5 коферментов НО: первичная структура белков отлична
α-кетоглутарат-ДГ – аналог ПДК Механизм практически идентичен ПДК: комплекс из 3 ферментов и тех же 5 коферментов НО: первичная структура белков отлична
 Механизм сукцинаттиокиназной реакции
Механизм сукцинаттиокиназной реакции
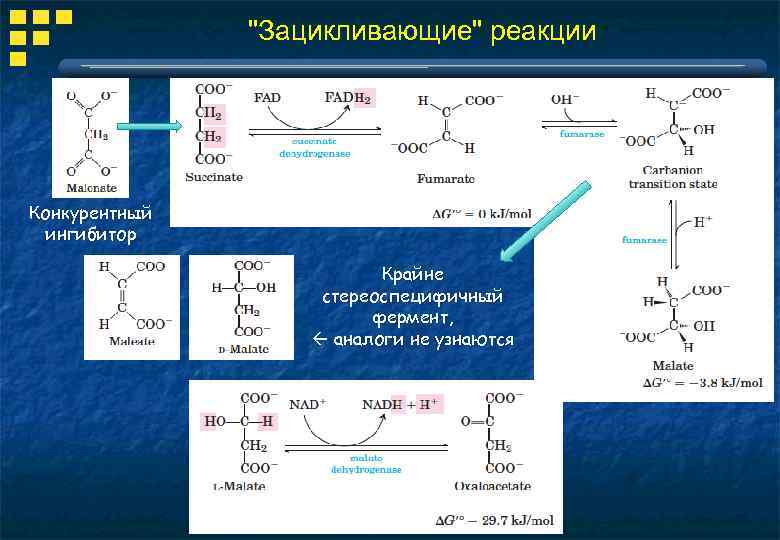 "Зацикливающие" реакции Конкурентный ингибитор Крайне стереоспецифичный фермент, аналоги не узнаются
"Зацикливающие" реакции Конкурентный ингибитор Крайне стереоспецифичный фермент, аналоги не узнаются
 Регуляция ЦТК Регуляция метаболитами: преимущественно за счет АТФ и НАДН - продуктов ЦТК
Регуляция ЦТК Регуляция метаболитами: преимущественно за счет АТФ и НАДН - продуктов ЦТК
 Другие функции ЦТК l Анаболические функции l Анаплеротические реакции
Другие функции ЦТК l Анаболические функции l Анаплеротические реакции
 Анаболическая функция ЦТК
Анаболическая функция ЦТК
 Биосинтез у анаэробов У них нет α-КГ-ДГ
Биосинтез у анаэробов У них нет α-КГ-ДГ
 Анаплеротические реакции Восполнение компонентов цикла и компенсация оксалоацетата
Анаплеротические реакции Восполнение компонентов цикла и компенсация оксалоацетата
 Пируваткарбоксилаза, биотин и оксалоацетат
Пируваткарбоксилаза, биотин и оксалоацетат
 Общая стратегия для решения сходных задач Ацетат СО 2 Ацил
Общая стратегия для решения сходных задач Ацетат СО 2 Ацил
 Челночный механизм транспорта ацетил-Ко. А Экспорт цитрата из митохондрий и возврат малата и пирувата
Челночный механизм транспорта ацетил-Ко. А Экспорт цитрата из митохондрий и возврат малата и пирувата