transpozoni_ta_retrotranspozoni.pptx
- Количество слайдов: 23
 ТРАНСПОЗОНИ ТА РЕТРОТРАНСПОЗОНИ Структура, механізм переміщення та роль у підтримці цілісності хромосом
ТРАНСПОЗОНИ ТА РЕТРОТРАНСПОЗОНИ Структура, механізм переміщення та роль у підтримці цілісності хромосом
 ПЛАН 1. 2. 3. 4. 5. Загальне поняття про транспозони Механізм транспозиції Еволюція геному еукаріот під впливом транспозонів Вплив транспозонів на регуляцію експресії генів Структурно-функціональна роль ретротранспозонів в геномі еукаріот. Ретротранспозони і проблема збереження кінців хромосом у ряду поколінь Ретротранспозони редагують двуланцюгові розриви ДНК
ПЛАН 1. 2. 3. 4. 5. Загальне поняття про транспозони Механізм транспозиції Еволюція геному еукаріот під впливом транспозонів Вплив транспозонів на регуляцію експресії генів Структурно-функціональна роль ретротранспозонів в геномі еукаріот. Ретротранспозони і проблема збереження кінців хромосом у ряду поколінь Ретротранспозони редагують двуланцюгові розриви ДНК
 1. ЗАГАЛЬНЕ ПОНЯТТЯ ПРО ТРАНСПОЗОНИ Транспозон – це послідовність ДНК, здатна переміщатися усередині геному в результаті процесу, що називається транспозицією. Ферменти, що каталізують транспозицію отримали назву транспозази. Вони здебільшого закодовані у ДНК самого транспозону. Довжина транспозонів приблизно від 2000 до 10000 пар нуклеотидів, присутні вони у геномі у кількості 5 -10 копій на диплоідну клітину.
1. ЗАГАЛЬНЕ ПОНЯТТЯ ПРО ТРАНСПОЗОНИ Транспозон – це послідовність ДНК, здатна переміщатися усередині геному в результаті процесу, що називається транспозицією. Ферменти, що каталізують транспозицію отримали назву транспозази. Вони здебільшого закодовані у ДНК самого транспозону. Довжина транспозонів приблизно від 2000 до 10000 пар нуклеотидів, присутні вони у геномі у кількості 5 -10 копій на диплоідну клітину.
 Транспозони - один з класів мобільних елементів геному, які, вбудовуючись в геном, можуть викликати мутації, в тому числі і такі значні як хромосомні перебудови. Вони відіграють важливу роль у процесах переносу лікарської стійкості серед мікроорганізмів, рекомбінації, та обміну генетичним матеріалом між різними видами як у природі (горизонтальний перенос генів), так і в ході генноінженерних досліджень. Також вони відіграють значну роль в процесі регуляції експресії генів.
Транспозони - один з класів мобільних елементів геному, які, вбудовуючись в геном, можуть викликати мутації, в тому числі і такі значні як хромосомні перебудови. Вони відіграють важливу роль у процесах переносу лікарської стійкості серед мікроорганізмів, рекомбінації, та обміну генетичним матеріалом між різними видами як у природі (горизонтальний перенос генів), так і в ході генноінженерних досліджень. Також вони відіграють значну роль в процесі регуляції експресії генів.
 Транспозони були відкриті в 1951 році Барбарою Мак-Клінток, яка в 1983 році була нагороджена за ці дослідження Нобелівською Премію. Транспозони зазвичай складаються з двох прямих або інвертований повторюваних послідовностей ДНК, між якими знаходяться гени, необхідні для транспозиції. Іноді в складі центральної частини транспозонів знаходяться гени, що забезпечують селективну перевагу для організму, що містить мобільний елемент
Транспозони були відкриті в 1951 році Барбарою Мак-Клінток, яка в 1983 році була нагороджена за ці дослідження Нобелівською Премію. Транспозони зазвичай складаються з двох прямих або інвертований повторюваних послідовностей ДНК, між якими знаходяться гени, необхідні для транспозиції. Іноді в складі центральної частини транспозонів знаходяться гени, що забезпечують селективну перевагу для організму, що містить мобільний елемент
 Три основні родини транспозонів Структура Гени, що входять до складу Спосіб переміщення у геномі Приклади Три основні родини транспозонів Кодують транспозазу Переміщуються у вигляді фрагменту ДНК або при вирізанні, реплікації Р-елемент (дрозофіла), Ac-Ds (кукуруза) IS 1 (E. coli) Кодують зворотну транскриптазу, нагадують ретровірус Переміщуються шляхом утворення проміжної форми РНК, що синтезується з промотора що локалізований у LTR Copla TY THE-1 bsl Кодують зворотню транскриптазу Переміщується шляхом утворення поміжної форми РНК , що синтезується з сусіднього промотора Р-елемент (дрозофіла), L 1 (людина) Короткі інвертовані Прямі довгі кінцеві повтори LTR Poly-A на 3´ кінці кожного РНК транскрипту
Три основні родини транспозонів Структура Гени, що входять до складу Спосіб переміщення у геномі Приклади Три основні родини транспозонів Кодують транспозазу Переміщуються у вигляді фрагменту ДНК або при вирізанні, реплікації Р-елемент (дрозофіла), Ac-Ds (кукуруза) IS 1 (E. coli) Кодують зворотну транскриптазу, нагадують ретровірус Переміщуються шляхом утворення проміжної форми РНК, що синтезується з промотора що локалізований у LTR Copla TY THE-1 bsl Кодують зворотню транскриптазу Переміщується шляхом утворення поміжної форми РНК , що синтезується з сусіднього промотора Р-елемент (дрозофіла), L 1 (людина) Короткі інвертовані Прямі довгі кінцеві повтори LTR Poly-A на 3´ кінці кожного РНК транскрипту
 2. МЕХАНІЗМ ТРАНСПОЗИЦІЇ Механізм транспозиції різноманітній У однієї групи транспозонів він практично повністю співпадає з частиною життєвого циклу ретровіруса. Такі елементи отримали незву ретротранспозони. Одним з найбільш вичених ретротранспозонів є знайдений у дріджів елемент Ту1
2. МЕХАНІЗМ ТРАНСПОЗИЦІЇ Механізм транспозиції різноманітній У однієї групи транспозонів він практично повністю співпадає з частиною життєвого циклу ретровіруса. Такі елементи отримали незву ретротранспозони. Одним з найбільш вичених ретротранспозонів є знайдений у дріджів елемент Ту1

 Багато трансозонів відрізняються від ретротранспозонів тим, що вони не існують поза хромосомою клітини хазяїна. Транспозази, що їх переміщують діють на ДНК транспозону у той час коли він перебуває в інтегрованому стані в клітині хазяїна. Транспозаза повязується з інвертованими короткими повторами на обох кінцях транпозону, і обмежує ці уінці для подальшої рекомбінації.
Багато трансозонів відрізняються від ретротранспозонів тим, що вони не існують поза хромосомою клітини хазяїна. Транспозази, що їх переміщують діють на ДНК транспозону у той час коли він перебуває в інтегрованому стані в клітині хазяїна. Транспозаза повязується з інвертованими короткими повторами на обох кінцях транпозону, і обмежує ці уінці для подальшої рекомбінації.
 Транспозон ДНК, що кодує транспозазу Короткі інвертовані повтори Транспозаза Хромосома
Транспозон ДНК, що кодує транспозазу Короткі інвертовані повтори Транспозаза Хромосома
 У інших транспозонів механізм транспозиції зводиться для того щоб розірвати та поновити ДНК. Існують транспозони, що при переміщенні реплікуються. транспозон Відокремлення білка
У інших транспозонів механізм транспозиції зводиться для того щоб розірвати та поновити ДНК. Існують транспозони, що при переміщенні реплікуються. транспозон Відокремлення білка
 3. ЕВОЛЮЦІЯ ГЕНОМУ ЕУКАРІОТ ПІД ВПЛИВОМ ТРАНСПОЗОНІВ Геноми зазвичай містять багато різноманітних транспозуючих елементів, або транспозонів На частку транспозонів доводиться принаймні 10% ДНК геному вищих еукаріот. Більшість цих елементів переміщається лише зрідка, але оскільки їх кількість у клітині велика, транспозиція значно впливає на різноманітність видів Наприклад, більше половини спонтанних мутацій, вивчених у дрозофіли, викликана вбудовуванням транспозона всередину мутованого гена або поблизу нього. Мутації можуть виникати або коли елемент вбудовується в ген, або коли він починає переміщатися в будь-яке інше місце. При вирізанні транспозону з хромосоми зазвичай він залишає на місці свого перебування одну з копій, що. Таким чином, переміщення транспозону супроводжується вставками і поділами у нуклеотидній послідовності
3. ЕВОЛЮЦІЯ ГЕНОМУ ЕУКАРІОТ ПІД ВПЛИВОМ ТРАНСПОЗОНІВ Геноми зазвичай містять багато різноманітних транспозуючих елементів, або транспозонів На частку транспозонів доводиться принаймні 10% ДНК геному вищих еукаріот. Більшість цих елементів переміщається лише зрідка, але оскільки їх кількість у клітині велика, транспозиція значно впливає на різноманітність видів Наприклад, більше половини спонтанних мутацій, вивчених у дрозофіли, викликана вбудовуванням транспозона всередину мутованого гена або поблизу нього. Мутації можуть виникати або коли елемент вбудовується в ген, або коли він починає переміщатися в будь-яке інше місце. При вирізанні транспозону з хромосоми зазвичай він залишає на місці свого перебування одну з копій, що. Таким чином, переміщення транспозону супроводжується вставками і поділами у нуклеотидній послідовності
 Транспозони вносять свій внесок у варіабельність геному і іншими засобами. Якщо два транспозона, які пізнаються одним і тим же сайт-специфічним ферментом рекомбінації (транспозазою), вбудовуються в сусідні сайти хромосоми, ДНК між ними може стати субстратом для транспозиції, здійснюваної за допомогою транспозази. Так що це дуже ефективний шлях переміщення екзонів, справедливо твердження, що транспозони можуть сприяти утворенню нових генів
Транспозони вносять свій внесок у варіабельність геному і іншими засобами. Якщо два транспозона, які пізнаються одним і тим же сайт-специфічним ферментом рекомбінації (транспозазою), вбудовуються в сусідні сайти хромосоми, ДНК між ними може стати субстратом для транспозиції, здійснюваної за допомогою транспозази. Так що це дуже ефективний шлях переміщення екзонів, справедливо твердження, що транспозони можуть сприяти утворенню нових генів
 Мутантний білок з додатковим доменом
Мутантний білок з додатковим доменом
 Іншою унікальною особливістю, що відрізняє транспозони від звичайних мутагенів, є їх здатність довгий час перебувати в стані спокою в хромосомі. Час від часу у частини популяції бурхливо активуються рух транспозонів і, відповідно, їх мутагенна активність. При таких катастрофічних змінах в геномах, названих транспозиційні вибухами, відбуваються майже одночасне переміщення транспозонів декількох типів. Вперше транспозиційні вибухи були виявлені в розвиваючихся рослинах кукурудзи. Якщо такі вибухи відбуваються в клітинах зародкової лінії, то вони викликають множинні зміни в геномі потомства окремої організму. Змінюючи властивості організму, транспозиційні вибухи підвищують ймовірність того, що дві нових ознаки, кожна з яких сам по собі не має селективної перевагами, виявлються вигідними, проявляючись разом у єдиній особини в популяції. Є дані про те, що у певних рослин транспозиційні вибухи можуть активуватися сильними стресами, викликаними зовнішніми умовами. Це призводить до появи різноманітних, випадковим чином модифікованих нащадків, частина з яких може виявитися краще пристосованої до виживання в нових умовах. Таким чином, транспозони не можна уявляти тільки як паразитів, а в деяких випадках вони здатні діяти як корисні симбіонти, сприяючи виживання тих видів, у геномі яких вони містяться.
Іншою унікальною особливістю, що відрізняє транспозони від звичайних мутагенів, є їх здатність довгий час перебувати в стані спокою в хромосомі. Час від часу у частини популяції бурхливо активуються рух транспозонів і, відповідно, їх мутагенна активність. При таких катастрофічних змінах в геномах, названих транспозиційні вибухами, відбуваються майже одночасне переміщення транспозонів декількох типів. Вперше транспозиційні вибухи були виявлені в розвиваючихся рослинах кукурудзи. Якщо такі вибухи відбуваються в клітинах зародкової лінії, то вони викликають множинні зміни в геномі потомства окремої організму. Змінюючи властивості організму, транспозиційні вибухи підвищують ймовірність того, що дві нових ознаки, кожна з яких сам по собі не має селективної перевагами, виявлються вигідними, проявляючись разом у єдиній особини в популяції. Є дані про те, що у певних рослин транспозиційні вибухи можуть активуватися сильними стресами, викликаними зовнішніми умовами. Це призводить до появи різноманітних, випадковим чином модифікованих нащадків, частина з яких може виявитися краще пристосованої до виживання в нових умовах. Таким чином, транспозони не можна уявляти тільки як паразитів, а в деяких випадках вони здатні діяти як корисні симбіонти, сприяючи виживання тих видів, у геномі яких вони містяться.
 4. ВПЛИВ ТРАНСПОЗОНІВ НА РЕГУЛЯЦІЮ ЕКСПРЕСІЇ ГЕНІВ Транспозони можуть впливати на регуляцію генів. Перебудови послідовностей ДНК, викликані транспозонами, часто змінюють експресію прилеглих генів, що може призвести до різних порушень у розвитку тварин чи рослин, наприклад їх пігментації. Велика частина таких змін щодо регуляції генів, як правило, виявляється шкідливою для організму, але деякі - можуть виявитися і корисними. Властивості мутацій, що викликаються транспозонами. незвичайні і дозволяють відрізнити їх від мутацій виникли внаслідок помилок у реплікації або репарації ДНК. Одна важлива відмінність полягає в тому, що при переміщенні транспозону поблизу гена часто виявляються нові послідовності, які діють як ділянки впізнавання для сайт-специфічних ДНКзв'язуючих білків, включаючи транспозазу і білки, що регулюють транскрипцію ДНК, яка входить до складу транспозона. Таким чином, ці послідовності можуть діяти як енхансери і посилювати транскрипцію генів, розташованих від них на відстані тисяч нуклеотидних пар.
4. ВПЛИВ ТРАНСПОЗОНІВ НА РЕГУЛЯЦІЮ ЕКСПРЕСІЇ ГЕНІВ Транспозони можуть впливати на регуляцію генів. Перебудови послідовностей ДНК, викликані транспозонами, часто змінюють експресію прилеглих генів, що може призвести до різних порушень у розвитку тварин чи рослин, наприклад їх пігментації. Велика частина таких змін щодо регуляції генів, як правило, виявляється шкідливою для організму, але деякі - можуть виявитися і корисними. Властивості мутацій, що викликаються транспозонами. незвичайні і дозволяють відрізнити їх від мутацій виникли внаслідок помилок у реплікації або репарації ДНК. Одна важлива відмінність полягає в тому, що при переміщенні транспозону поблизу гена часто виявляються нові послідовності, які діють як ділянки впізнавання для сайт-специфічних ДНКзв'язуючих білків, включаючи транспозазу і білки, що регулюють транскрипцію ДНК, яка входить до складу транспозона. Таким чином, ці послідовності можуть діяти як енхансери і посилювати транскрипцію генів, розташованих від них на відстані тисяч нуклеотидних пар.
 А. Дрозофіла Б. Левиний зів В. Кукурудза
А. Дрозофіла Б. Левиний зів В. Кукурудза
 Геноми вищих еукаріот, де довгі послідовності некодирующей ДНК перемежовуються відносно короткими кодуючими ділянками, це «родюче підґрунтя» для інтеграції і виключення мобільних елементів. У зв'язку з тим, що на транскрипцію генів впливають і віддалені від них на десятки тисяч нуклеотидних пар ділянки, можна очікувати, що багато змін, що виникли при транспозиції геному, вплинуть і на експресію генів. І навпаки, мабуть, лише деякі перебудови приведуть до руйнування коротких екзонів, що містять кодують послідовності. Транспозони, переміщаючи енхансери по геному, можуть сприяти оптимізації регуляції генів з метою виживання організму в ряді поколінь.
Геноми вищих еукаріот, де довгі послідовності некодирующей ДНК перемежовуються відносно короткими кодуючими ділянками, це «родюче підґрунтя» для інтеграції і виключення мобільних елементів. У зв'язку з тим, що на транскрипцію генів впливають і віддалені від них на десятки тисяч нуклеотидних пар ділянки, можна очікувати, що багато змін, що виникли при транспозиції геному, вплинуть і на експресію генів. І навпаки, мабуть, лише деякі перебудови приведуть до руйнування коротких екзонів, що містять кодують послідовності. Транспозони, переміщаючи енхансери по геному, можуть сприяти оптимізації регуляції генів з метою виживання організму в ряді поколінь.
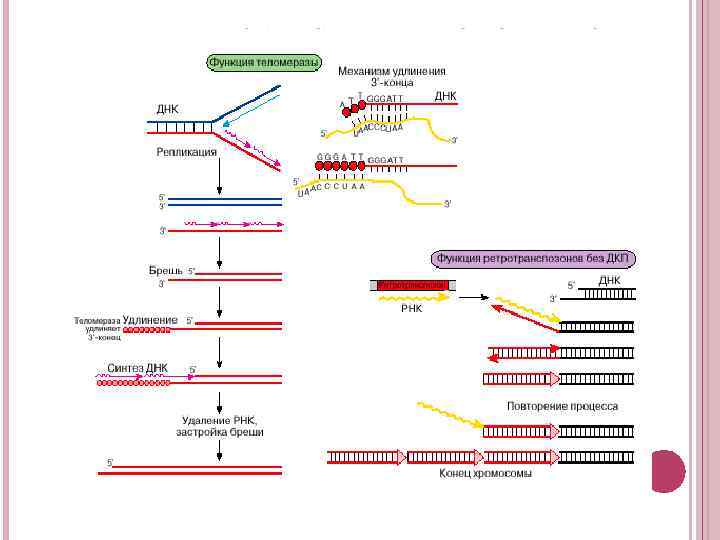
 5. СТРУКТУРНО-ФУНКЦІОНАЛЬНА РОЛЬ РЕТРОТРАНСПОЗОНІВ У ГЕНОМІ ЕУКАРІОТ При відтворенні ДНК перед клітинним поділом синтез ДНК починається з утворенняи затравки РНК, оскільки фермент ДНКполімераза здатний тільки додавати дезоксірібонуклеотідниі ланки до 3'-кінця полинуклеотидного ланцюга, але нездатний починати синтез ланцюга ДНК. Затравака потім видаляється, і прориви забудовуються. Однак на одному з кінців реплицируємої молекул залишиться прорив, який не вдається забудувати за допомогою ДНК-полімерази, що працює в 5'-3' напрямі. Виникає небезпека, що самотньо виступаючий однониткових кінець ДНК буде знищений яким-небудь ферментом, в результаті чого молекула вкорочувається з кінця. Якщо не вжити відповідних заходів, то при кожному акті реплікації ДНК хромосома буде зменшуватися з кінців. У кінцевому підсумку можуть бути втрачені важливі гени і клітина загине. Зазвичай для збереження кінця ДНК використовується фермент теломераза, що складається з двох компонентів: білка і РНК-матриці, за допомогою якої подовжується кінець ДНК.
5. СТРУКТУРНО-ФУНКЦІОНАЛЬНА РОЛЬ РЕТРОТРАНСПОЗОНІВ У ГЕНОМІ ЕУКАРІОТ При відтворенні ДНК перед клітинним поділом синтез ДНК починається з утворенняи затравки РНК, оскільки фермент ДНКполімераза здатний тільки додавати дезоксірібонуклеотідниі ланки до 3'-кінця полинуклеотидного ланцюга, але нездатний починати синтез ланцюга ДНК. Затравака потім видаляється, і прориви забудовуються. Однак на одному з кінців реплицируємої молекул залишиться прорив, який не вдається забудувати за допомогою ДНК-полімерази, що працює в 5'-3' напрямі. Виникає небезпека, що самотньо виступаючий однониткових кінець ДНК буде знищений яким-небудь ферментом, в результаті чого молекула вкорочувається з кінця. Якщо не вжити відповідних заходів, то при кожному акті реплікації ДНК хромосома буде зменшуватися з кінців. У кінцевому підсумку можуть бути втрачені важливі гени і клітина загине. Зазвичай для збереження кінця ДНК використовується фермент теломераза, що складається з двох компонентів: білка і РНК-матриці, за допомогою якої подовжується кінець ДНК.
 Одна з причин старіння бачиться в тому, що за відсутності теломерази у деяких тканинах відбувається укорочення хромосоми із втратою важливих генів. Навпаки, безсмертя ряду клітин у культурі поза організмом, властиве, як правило, клітинам з пухлин, пояснюється реактивацією теломерази. У плодової мушки дрозофіли відсутня теломераза. але кінці ДНК подовжуються за рахунок переміщень ретротранспозонів. На цьому прикладі вперше показана важлива структурна і функціональна роль ретротранспозонів. Вони виступають як компоненти генома, які рятують хромосому від укорочування. В якості рятувальників виступають ретротранспозони, пов'язані з домами, без довгих кінцевих повторів. Ретротранспозони переміщуються, утворюючи повторювану структуру, в якій елементи з'єднані один з одним за типом "голова до хвоста“. Спочатку на РНК-транскрипті як на матриці за допомогою ревертази будується комплементарна нитка ДНК, а потім після видалення РНКматриці добудовується інша. Таким чином, якщо ці ретротранспозони і існували колись як елементи-паразити, то згодом геном господаря пристосував їх для виконання такої важливої функції, як збереження кінцевих ділянок хромосом. Ці ретротранспозони стали безцінними помічниками, що рятують хромосому від втрати генів.
Одна з причин старіння бачиться в тому, що за відсутності теломерази у деяких тканинах відбувається укорочення хромосоми із втратою важливих генів. Навпаки, безсмертя ряду клітин у культурі поза організмом, властиве, як правило, клітинам з пухлин, пояснюється реактивацією теломерази. У плодової мушки дрозофіли відсутня теломераза. але кінці ДНК подовжуються за рахунок переміщень ретротранспозонів. На цьому прикладі вперше показана важлива структурна і функціональна роль ретротранспозонів. Вони виступають як компоненти генома, які рятують хромосому від укорочування. В якості рятувальників виступають ретротранспозони, пов'язані з домами, без довгих кінцевих повторів. Ретротранспозони переміщуються, утворюючи повторювану структуру, в якій елементи з'єднані один з одним за типом "голова до хвоста“. Спочатку на РНК-транскрипті як на матриці за допомогою ревертази будується комплементарна нитка ДНК, а потім після видалення РНКматриці добудовується інша. Таким чином, якщо ці ретротранспозони і існували колись як елементи-паразити, то згодом геном господаря пристосував їх для виконання такої важливої функції, як збереження кінцевих ділянок хромосом. Ці ретротранспозони стали безцінними помічниками, що рятують хромосому від втрати генів.
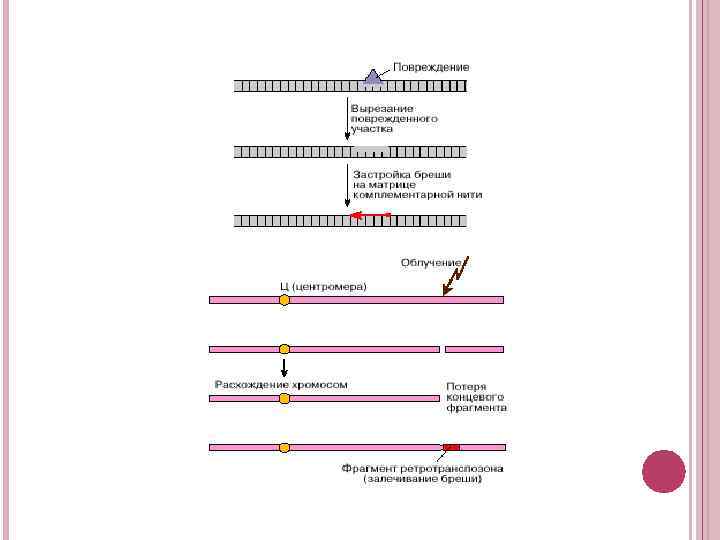
 Пошкодження однієї з комплементарних ниток ДНК можуть бути усунені за рахунок видалення цієї ділянки і його ресинтезу з використанням непошкодженою комплементарної нитки ДНК. Складніше виправляється двуланцюговий розрив, що призводить до утворення двох окремих фрагментів подвійної спіралі ДНК. Хромосоми регулярно розходяться по дочірнім клітинам, якщо вони не втратили центромери, до якої прикріплюються нитки веретена поділу і розтягають хромосоми. Однак фрагмент хромосоми, що лежить від центромери далі, за розривом, буде втрачено. Зазвичай двухнитевой розрив заліковується за допомогою гомологічною молекули ДНК, наприклад сестринської, тільки що реплікованої нитки. Цей процес здійснюється шляхом ресинтезу копії втраченої ДНК на місці дірки, що утворилася. Однак якщо клітина позбавлена звичайної системи заліковування двуланцюгового розриву, то як заплатки може бути використана рухома ДНК. Виявилося, що в ролі такої рухомий ДНК може виступати ДНК ретротранспозонів. У цьому випадку рятувальну функцію здійснює клас ретротранспозонів, що містять ДКП. Заплатка дозволить хромосомі зберегти цілісність і не втратити кінцевого фрагмента. Правда, пролом в двуланцюгової спіралі ДНК буде заліплені латкою з ретротранспозону, тобто вихідна нуклеотидна послідовність не буде відновлена. Однак якщо район розриву не містив істотного гена, то клітина, а можливо, і організм збережуть життєздатність. Можливість участі ретротранспозонів, що містять довгі кінцеві повтори, в процесі загоєння двуланцюгових розривів була виявлена в клітинах дріжджів недавно, тому молекулярні механізми виявленого явища залишаються поки нез'ясованими.
Пошкодження однієї з комплементарних ниток ДНК можуть бути усунені за рахунок видалення цієї ділянки і його ресинтезу з використанням непошкодженою комплементарної нитки ДНК. Складніше виправляється двуланцюговий розрив, що призводить до утворення двох окремих фрагментів подвійної спіралі ДНК. Хромосоми регулярно розходяться по дочірнім клітинам, якщо вони не втратили центромери, до якої прикріплюються нитки веретена поділу і розтягають хромосоми. Однак фрагмент хромосоми, що лежить від центромери далі, за розривом, буде втрачено. Зазвичай двухнитевой розрив заліковується за допомогою гомологічною молекули ДНК, наприклад сестринської, тільки що реплікованої нитки. Цей процес здійснюється шляхом ресинтезу копії втраченої ДНК на місці дірки, що утворилася. Однак якщо клітина позбавлена звичайної системи заліковування двуланцюгового розриву, то як заплатки може бути використана рухома ДНК. Виявилося, що в ролі такої рухомий ДНК може виступати ДНК ретротранспозонів. У цьому випадку рятувальну функцію здійснює клас ретротранспозонів, що містять ДКП. Заплатка дозволить хромосомі зберегти цілісність і не втратити кінцевого фрагмента. Правда, пролом в двуланцюгової спіралі ДНК буде заліплені латкою з ретротранспозону, тобто вихідна нуклеотидна послідовність не буде відновлена. Однак якщо район розриву не містив істотного гена, то клітина, а можливо, і організм збережуть життєздатність. Можливість участі ретротранспозонів, що містять довгі кінцеві повтори, в процесі загоєння двуланцюгових розривів була виявлена в клітинах дріжджів недавно, тому молекулярні механізми виявленого явища залишаються поки нез'ясованими.