2bfdcbc56d3720c0bfec630349eb995d.ppt
- Количество слайдов: 176
 Transkripsiyon Prof. Dr. Selma Yılmazer İ. Ü Cerrahpaşa Tıp Fakültesi Tıbbi Biyoloji A. D
Transkripsiyon Prof. Dr. Selma Yılmazer İ. Ü Cerrahpaşa Tıp Fakültesi Tıbbi Biyoloji A. D
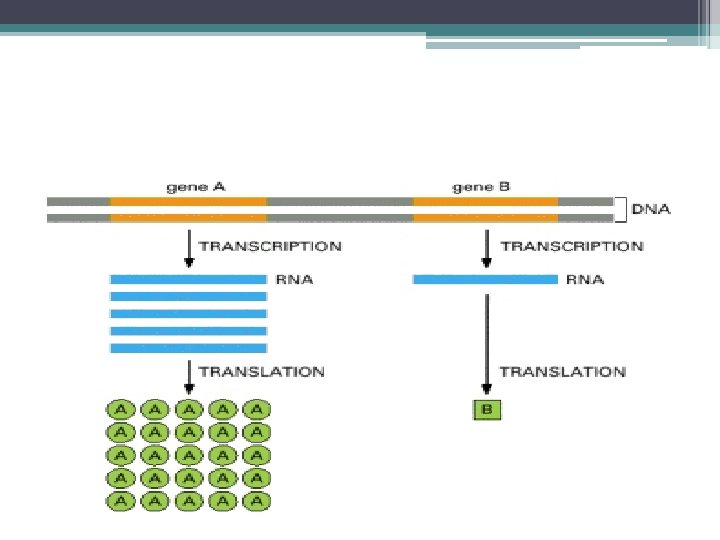
 • DNA molekülünün taşıdığı bilginin protein moleküllerine çevrilmesine Gen ekspresyonu") Gen ekspresyonu (gen anlatımı) • DNA molekülünün taşıdığı bilginin protein moleküllerine çevrilmesine Gen ekspresyonu denir. • Gen ekspresyonu 2 aşamada gerçekleşir: 1. Transkripsiyon (Yazılım): DNA nın taşıdığı bilgi, RNA Moleküllerine aktarılır. 1. Translasyon (Çeviri): Kalıtsal bilgi, m. RNA aracılığı ile proteine çevrilir. DNA, ▫ protein sentezi için doğrudan kalıp görevi yapmaz. ▫ Aracı moleküllere yani m. RNA lara ihtiyaç vardır.
Gen ekspresyonu (gen anlatımı) • DNA molekülünün taşıdığı bilginin protein moleküllerine çevrilmesine Gen ekspresyonu denir. • Gen ekspresyonu 2 aşamada gerçekleşir: 1. Transkripsiyon (Yazılım): DNA nın taşıdığı bilgi, RNA Moleküllerine aktarılır. 1. Translasyon (Çeviri): Kalıtsal bilgi, m. RNA aracılığı ile proteine çevrilir. DNA, ▫ protein sentezi için doğrudan kalıp görevi yapmaz. ▫ Aracı moleküllere yani m. RNA lara ihtiyaç vardır.
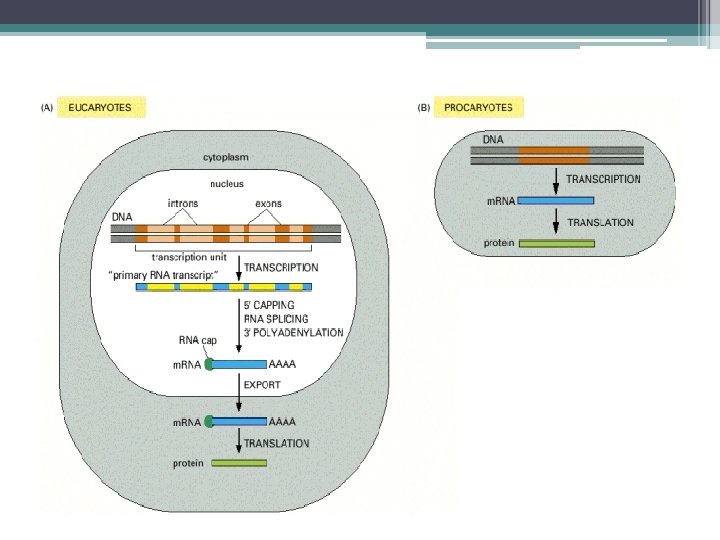
 Gen ekspresyonunun (gen anlatımı) • ilk basamağı RNA sentezidir. (Transkripsiyon)") Gen ekspresyonu (gen anlatımı) Gen ekspresyonunun (gen anlatımı) • ilk basamağı RNA sentezidir. (Transkripsiyon) Ökaryot hücrelerde Transkripsiyon; • RNA sentezlenmesi (ilk transkriptin oluşması), • Sentezlenen RNA nın işlevsel forma yani olgun m. RNA ya dönüştürülmesi; RNA işlenmesi ile gerçekleşir. (Örn. İntronların kesilip çıkartılması, uç modifikasyonları)
Gen ekspresyonu (gen anlatımı) Gen ekspresyonunun (gen anlatımı) • ilk basamağı RNA sentezidir. (Transkripsiyon) Ökaryot hücrelerde Transkripsiyon; • RNA sentezlenmesi (ilk transkriptin oluşması), • Sentezlenen RNA nın işlevsel forma yani olgun m. RNA ya dönüştürülmesi; RNA işlenmesi ile gerçekleşir. (Örn. İntronların kesilip çıkartılması, uç modifikasyonları)
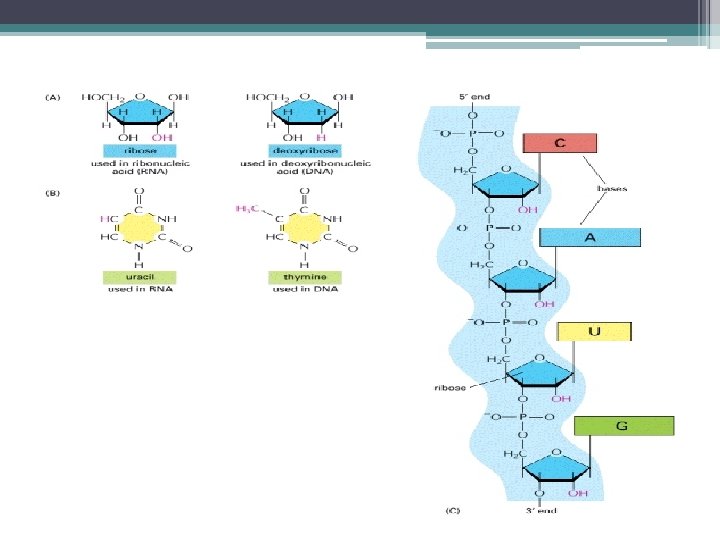
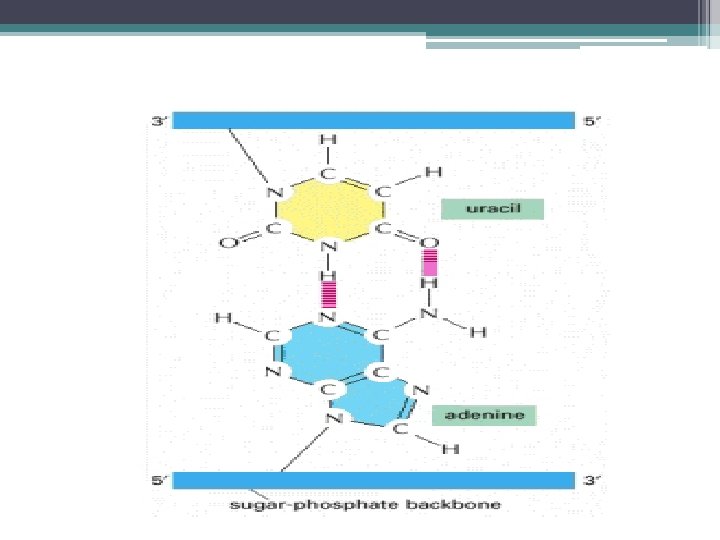
 Transkripsiyon; • DNA nın gen adı verilen bölgelerinin • m. RNA, r. RNA ve t. RNA vermek üzere kopyalanmasıdır. • Transkripsiyonu gerçekleştiren enzim RNA polimerazdır. • 1960’ da J. Hurwitz ve S. Weissman tarafından bulunmuştur • m. RNA, DNA nın dizisine komplementer (tamamlayıcı) bir baz dizisi içerir.
Transkripsiyon; • DNA nın gen adı verilen bölgelerinin • m. RNA, r. RNA ve t. RNA vermek üzere kopyalanmasıdır. • Transkripsiyonu gerçekleştiren enzim RNA polimerazdır. • 1960’ da J. Hurwitz ve S. Weissman tarafından bulunmuştur • m. RNA, DNA nın dizisine komplementer (tamamlayıcı) bir baz dizisi içerir.
: Protein Sentezi için kalıp görevi") RNA Çeşitleri • Haberci RNA’ lar (m. RNA lar): Protein Sentezi için kalıp görevi yapar. • Ribozomal RNA’ lar (r. RNA lar) ve • Taşıyıcı RNA’ lar (t. RNA lar) m. RNA nın proteine çevrilmesinde görev yapar. • Küçük RNA’ lar – m. RNA kırpılması – r. RNA işlenmesi v. b. – Gen ekspresyonu düzenlenmesi
RNA Çeşitleri • Haberci RNA’ lar (m. RNA lar): Protein Sentezi için kalıp görevi yapar. • Ribozomal RNA’ lar (r. RNA lar) ve • Taşıyıcı RNA’ lar (t. RNA lar) m. RNA nın proteine çevrilmesinde görev yapar. • Küçük RNA’ lar – m. RNA kırpılması – r. RNA işlenmesi v. b. – Gen ekspresyonu düzenlenmesi
 Transkripsiyon • Transkripsiyon için – DNA Kalıbına – RNA polimeraza – Ribonükleotid trifosfatlara (ATP, GTP, CTP, UTP) – Magnezyum İyonuna ihtiyaç vardır.
Transkripsiyon • Transkripsiyon için – DNA Kalıbına – RNA polimeraza – Ribonükleotid trifosfatlara (ATP, GTP, CTP, UTP) – Magnezyum İyonuna ihtiyaç vardır.
 RNA Sentezinin Aşamaları • • RNA Polimerazın DNA Kalıbına bağlanması. Sentezin başlaması Zincirin uzaması Sentezin sonlanması
RNA Sentezinin Aşamaları • • RNA Polimerazın DNA Kalıbına bağlanması. Sentezin başlaması Zincirin uzaması Sentezin sonlanması
 Prokaryotlarda Transkripsiyon • E. Coli, ▫ ▫ Transkripsiyon çalışmalarında bir model. m. RNA, E. Coli’ de bulundu. RNA Polimeraz, E. Coli’ den saflaştırıldı. Gen ekspresyonu regülasyonunu sağlayan mekanizmalar E. Coli’ de keşfedildi.
Prokaryotlarda Transkripsiyon • E. Coli, ▫ ▫ Transkripsiyon çalışmalarında bir model. m. RNA, E. Coli’ de bulundu. RNA Polimeraz, E. Coli’ den saflaştırıldı. Gen ekspresyonu regülasyonunu sağlayan mekanizmalar E. Coli’ de keşfedildi.
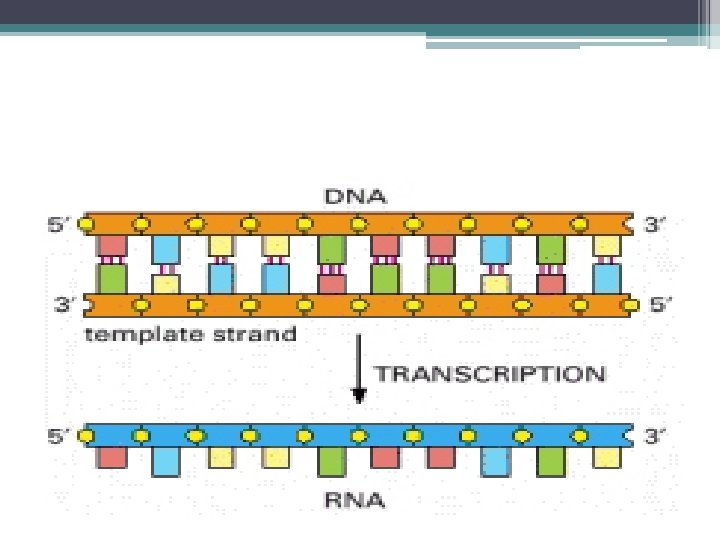
 RNA Polimeraz ve Transkripsiyon • RNA sentezinden sorumlu olan enzim; RNA Polimerazdır. • DNA’ yı kalıp olarak kullanarak • Ona komplementer olan bazları bir araya getirir. • Ribonukleozid 5’-trifosfatların (NTP ler) polimerleşmesini katalizler.
RNA Polimeraz ve Transkripsiyon • RNA sentezinden sorumlu olan enzim; RNA Polimerazdır. • DNA’ yı kalıp olarak kullanarak • Ona komplementer olan bazları bir araya getirir. • Ribonukleozid 5’-trifosfatların (NTP ler) polimerleşmesini katalizler.
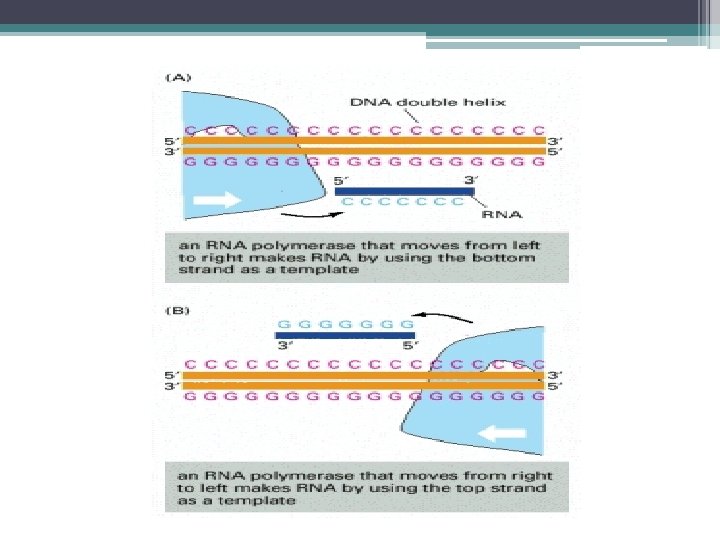
 •") RNA Polimeraz • RNA zincirinin uzamasını daima 5’->3’ Yönünde katalizler. (DNA Polimeraz gibi) • Primerlere ihtiyaç duymaz (DNA Polimerazdan farklı olarak) • Sentezlenen RNA zinciri ile kalıp DNA ipliği birbirine antiparaleldir. • Prokaryotlarda tek bir RNA Polimeraz, m. RNA, t. RNA ve r. RNA sentezini gerçekleştirir.
RNA Polimeraz • RNA zincirinin uzamasını daima 5’->3’ Yönünde katalizler. (DNA Polimeraz gibi) • Primerlere ihtiyaç duymaz (DNA Polimerazdan farklı olarak) • Sentezlenen RNA zinciri ile kalıp DNA ipliği birbirine antiparaleldir. • Prokaryotlarda tek bir RNA Polimeraz, m. RNA, t. RNA ve r. RNA sentezini gerçekleştirir.
 RNA Polimeraz • RNA Polimeraz; Çok sayıda polipeptidden oluşmuş kompleks bir enzimdir. • Bakteri RNA Polimerazı, α, β, β’ ω ve σ alt birimlerinden oluşur. • σ(sigma) alt birimi, komplekse daha zayıf bağlanır. • Yeni sentezlenen RNA birkaç nükleotid uzunluğa eriştiğinde σ, diğer alt birimlerden ayrılır.
RNA Polimeraz • RNA Polimeraz; Çok sayıda polipeptidden oluşmuş kompleks bir enzimdir. • Bakteri RNA Polimerazı, α, β, β’ ω ve σ alt birimlerinden oluşur. • σ(sigma) alt birimi, komplekse daha zayıf bağlanır. • Yeni sentezlenen RNA birkaç nükleotid uzunluğa eriştiğinde σ, diğer alt birimlerden ayrılır.
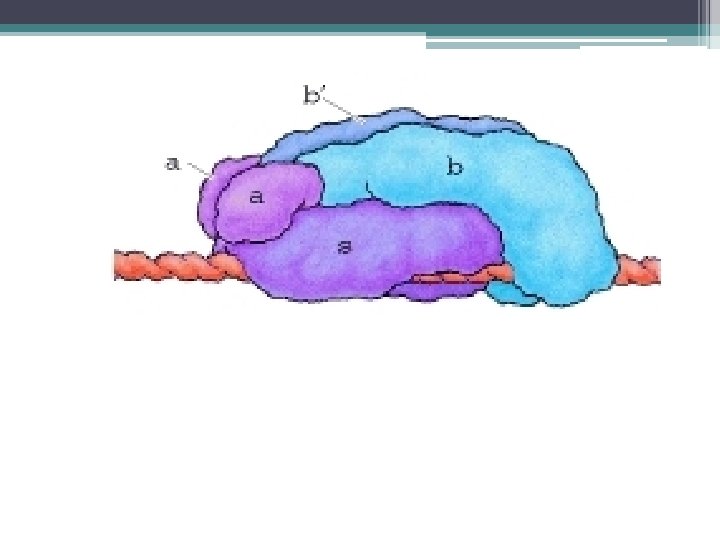
 alt biriminden oluşan") RNA Polimeraz • İki α, bir β’ ve bir ω (omega) alt biriminden oluşan çekirdek RNA polimeraz, RNA nın uzama aşamasını gerçekleştirir. • Çekirdek RNA Polimeraz NTP lerin RNA ya polimerleşmesini katalizler. • Bu işlem için σ gerekmez.
RNA Polimeraz • İki α, bir β’ ve bir ω (omega) alt biriminden oluşan çekirdek RNA polimeraz, RNA nın uzama aşamasını gerçekleştirir. • Çekirdek RNA Polimeraz NTP lerin RNA ya polimerleşmesini katalizler. • Bu işlem için σ gerekmez.
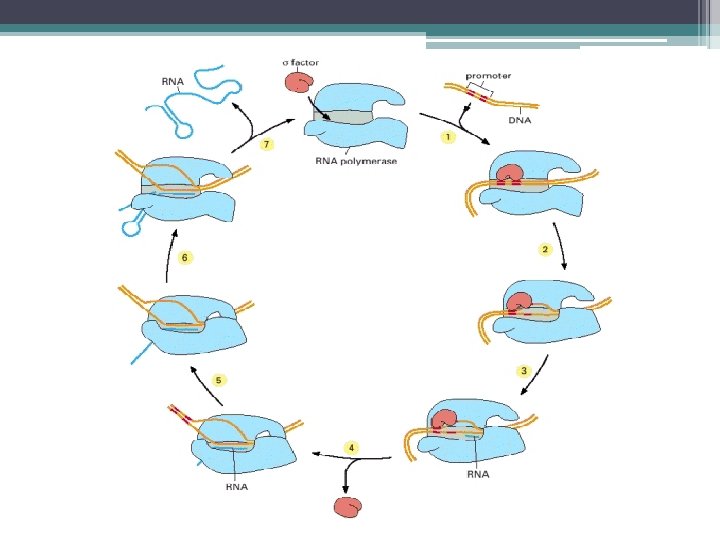
 RNA Polimeraz • σ , kompleksin transkripsiyonun başlangıç yeri olan DNA dizisine bağlanması için gereklidir. • İş görebilen bir RNAnın oluşması için sentez, gendeki başlangıç noktasından başlamak zorundadır. • RNA Polimerazın bağlandığı bu özgün DNA dizisine, Promotör denir. • Transkripsiyon, genlerin başlangıcındaki bu özgün yerlerde başlar.
RNA Polimeraz • σ , kompleksin transkripsiyonun başlangıç yeri olan DNA dizisine bağlanması için gereklidir. • İş görebilen bir RNAnın oluşması için sentez, gendeki başlangıç noktasından başlamak zorundadır. • RNA Polimerazın bağlandığı bu özgün DNA dizisine, Promotör denir. • Transkripsiyon, genlerin başlangıcındaki bu özgün yerlerde başlar.
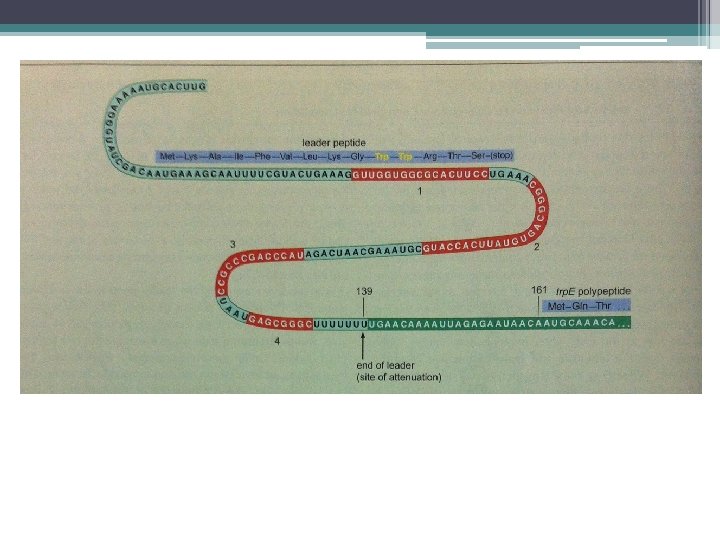
 Promotör Dizisi • E. Coli’ de keşfedildi. • Değişik genlerde transkripsiyon başlangıç noktasının yukarı yönünde birbirine benzeyen 2 farklı dizi bulundu. • Bu ortak diziler 6 nukleotid içerir. • Transkripsiyon başlangıç yerinin (+1) • -10 ve • -35 baz çifti öncesinde bulunur.
Promotör Dizisi • E. Coli’ de keşfedildi. • Değişik genlerde transkripsiyon başlangıç noktasının yukarı yönünde birbirine benzeyen 2 farklı dizi bulundu. • Bu ortak diziler 6 nukleotid içerir. • Transkripsiyon başlangıç yerinin (+1) • -10 ve • -35 baz çifti öncesinde bulunur.
 E. coli’nin Promotör Dizileri • -10 ve -35 elemanları olarak adlandırılırlar. • Bu diziler, farklı genlerde birbirlerine benzer. • Transkripsiyon başlangıç yeri, +1 olarak tanımlanır. • -10 ve -35 ortak dizilerdeki mutasyonlar, promotör işlevi üzerinde güçlü etki yaratır. • σ alt birimi, -35 ve -10 promotör bölgelerinin ikisine de bağlanır.
E. coli’nin Promotör Dizileri • -10 ve -35 elemanları olarak adlandırılırlar. • Bu diziler, farklı genlerde birbirlerine benzer. • Transkripsiyon başlangıç yeri, +1 olarak tanımlanır. • -10 ve -35 ortak dizilerdeki mutasyonlar, promotör işlevi üzerinde güçlü etki yaratır. • σ alt birimi, -35 ve -10 promotör bölgelerinin ikisine de bağlanır.
 E. coli’nin promotör dizileri
E. coli’nin promotör dizileri
 • DNA ayak izi yöntemi ile RNA polimerazın Promotöre bağlandığı yerler tanımlanmıştır
• DNA ayak izi yöntemi ile RNA polimerazın Promotöre bağlandığı yerler tanımlanmıştır
 DNA sarmalının lokal olarak 14") Transkripsiyonun başlaması • RNA polimerazın bağlanması, (Kapalı promotör kompleksi) DNA sarmalının lokal olarak 14 bazı açarak (-12 den +2 ye kadar) çözülmesine neden olur. Böylece tek iplikli DNA oluşur. (Açık promotör kompleksi) • RNA polimerazın Transkripsiyonu başlatması DNA kalıbını kullanarak buna tamamlayıcı olan ilk iki serbest NTP yi fosfodiester bağı ile birleştirerek başlatır. • RNA polimeraz senteze devam eder. • Sentezlenen zincir, 10 nükleotid uzunluğa erişince, σ ayrılır. .
Transkripsiyonun başlaması • RNA polimerazın bağlanması, (Kapalı promotör kompleksi) DNA sarmalının lokal olarak 14 bazı açarak (-12 den +2 ye kadar) çözülmesine neden olur. Böylece tek iplikli DNA oluşur. (Açık promotör kompleksi) • RNA polimerazın Transkripsiyonu başlatması DNA kalıbını kullanarak buna tamamlayıcı olan ilk iki serbest NTP yi fosfodiester bağı ile birleştirerek başlatır. • RNA polimeraz senteze devam eder. • Sentezlenen zincir, 10 nükleotid uzunluğa erişince, σ ayrılır. .
 E. Coli RNA polimerazı ile transkripsiyon
E. Coli RNA polimerazı ile transkripsiyon
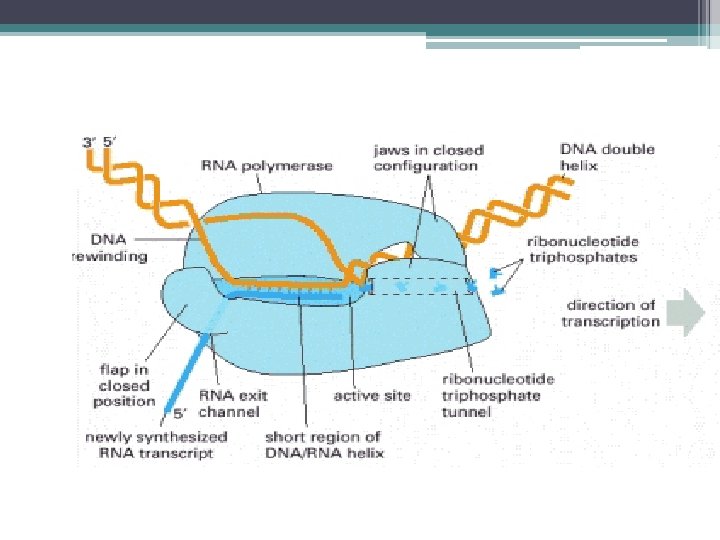
 Zincirin Uzaması • RNA polimeraz ileri doğru hareket eder, sentez devam eder. • RNA zinciri uzar • Uzama sırasında polimeraz, kalıp DNA ile birleşik kalır. • Uzama sırasında polimeraz, (m. RNA sentezinde) ▫ Transkripsiyon bölgesinde ~15 bazlık açık bölge kalacak şekilde öndeki DNA yı açar, arkadaki DNA yı geri sarar. ▫ β ve β’ alt birimler arasındaki kanal, 20 bç lik DNA barındırır ve polimerazın aktif bölgesini oluşturur.
Zincirin Uzaması • RNA polimeraz ileri doğru hareket eder, sentez devam eder. • RNA zinciri uzar • Uzama sırasında polimeraz, kalıp DNA ile birleşik kalır. • Uzama sırasında polimeraz, (m. RNA sentezinde) ▫ Transkripsiyon bölgesinde ~15 bazlık açık bölge kalacak şekilde öndeki DNA yı açar, arkadaki DNA yı geri sarar. ▫ β ve β’ alt birimler arasındaki kanal, 20 bç lik DNA barındırır ve polimerazın aktif bölgesini oluşturur.
 Sonlanma: – RNA sentezi RNA polimeraz, sonlandırma sinyali ile karşılaşana kadar devam eder. – Burada transkripsiyon durur. – Sentezlenen RNA, polimerazdan ayrılır. – Polimeraz da DNA dan ayrılır. – E. Coli’ de en basit bir sonlandırma sinyali vardır. – GC zengin ters tekrarlar, baz eşleşmesi ile sap-ilmik yapısı oluşturur. – Bu yapı, RNA nın DNA ile ilişkisini bozar. – Transkripsiyon sonlanır.
Sonlanma: – RNA sentezi RNA polimeraz, sonlandırma sinyali ile karşılaşana kadar devam eder. – Burada transkripsiyon durur. – Sentezlenen RNA, polimerazdan ayrılır. – Polimeraz da DNA dan ayrılır. – E. Coli’ de en basit bir sonlandırma sinyali vardır. – GC zengin ters tekrarlar, baz eşleşmesi ile sap-ilmik yapısı oluşturur. – Bu yapı, RNA nın DNA ile ilişkisini bozar. – Transkripsiyon sonlanır.
 Transkripsiyonun sonlanması 4 adenin nukleotidi tarafından izlenen GC den zengin ters tekrarlar ile belirlenir. Ters tekrarlar RNA da kararlı bir sap -ilmik yapısı oluşturur. Sonuçta RNA Kalıp DNA dan ayrılır.
Transkripsiyonun sonlanması 4 adenin nukleotidi tarafından izlenen GC den zengin ters tekrarlar ile belirlenir. Ters tekrarlar RNA da kararlı bir sap -ilmik yapısı oluşturur. Sonuçta RNA Kalıp DNA dan ayrılır.
 Ökaryotlarda Transkripsiyon • Prokaryot ve Ökaryot Hücreler Arasındaki Farklar: 1 - Bakteride tek bir RNA polimeraz (tüm genlerin transkripsiyonundan sorumlu) ▫ Ökaryotik hücrelerde, birden fazla RNA polimerazlar bulunur. 2 - Ökaryotik RNA polimerazlar promotör diziye, ek proteinler ile bağlanır.
Ökaryotlarda Transkripsiyon • Prokaryot ve Ökaryot Hücreler Arasındaki Farklar: 1 - Bakteride tek bir RNA polimeraz (tüm genlerin transkripsiyonundan sorumlu) ▫ Ökaryotik hücrelerde, birden fazla RNA polimerazlar bulunur. 2 - Ökaryotik RNA polimerazlar promotör diziye, ek proteinler ile bağlanır.
 Ökaryotik RNA Polimerazları ▫ 3 farklı Nükleer RNA Polimeraz bulunur ▫ RNA Polimeraz III 1 -m. RNA’ lar ; RNA Polimeraz II ile 2 -r. RNA’ lar ve t. RNA’ lar ; RNA Polimeraz I ve III ile (Pol I 28 s, 18 s, 5. 8 s r. RNA ların; Pol III 5 s r. RNA ların sentezi) 3 -Küçük RNA lar; sn. RNA (küçük nükleer RNA ; kesip ekleme ) , sc. RNA [küçük sitoplazmik RNA; protein taşınması) RNA Pol III ve RNA Pol II tarafından sentezlenir.
Ökaryotik RNA Polimerazları ▫ 3 farklı Nükleer RNA Polimeraz bulunur ▫ RNA Polimeraz III 1 -m. RNA’ lar ; RNA Polimeraz II ile 2 -r. RNA’ lar ve t. RNA’ lar ; RNA Polimeraz I ve III ile (Pol I 28 s, 18 s, 5. 8 s r. RNA ların; Pol III 5 s r. RNA ların sentezi) 3 -Küçük RNA lar; sn. RNA (küçük nükleer RNA ; kesip ekleme ) , sc. RNA [küçük sitoplazmik RNA; protein taşınması) RNA Pol III ve RNA Pol II tarafından sentezlenir.
 Ökaryotik RNA Polimerazlar • Kloroplast ve mitokondrilerde farklı RNA polimerazlar vardır. • Nükleer RNA Polimerazların üçü de • 12 -17 alt birimden oluşan kompleks enzimlerdir. • Üçü de 9 adet benzer alt birim içerir. • Bunların 5 tanesi bakteri RNA polimerazın α, β, β’ ve ω alt birimlerine benzer.
Ökaryotik RNA Polimerazlar • Kloroplast ve mitokondrilerde farklı RNA polimerazlar vardır. • Nükleer RNA Polimerazların üçü de • 12 -17 alt birimden oluşan kompleks enzimlerdir. • Üçü de 9 adet benzer alt birim içerir. • Bunların 5 tanesi bakteri RNA polimerazın α, β, β’ ve ω alt birimlerine benzer.
 Ökaryotlarda Transkripsiyon • Prokaryotlarda olduğu gibi, 4 aşamada gerçekleşir ▫ ▫ RNA Polimerazın DNA ya bağlanması RNA sentezinin başlaması RNA nın uzaması RNA sentezinin sonlanması
Ökaryotlarda Transkripsiyon • Prokaryotlarda olduğu gibi, 4 aşamada gerçekleşir ▫ ▫ RNA Polimerazın DNA ya bağlanması RNA sentezinin başlaması RNA nın uzaması RNA sentezinin sonlanması
 önünde • -80 konumunda") Ökaryotlarda promotör bölge • Ökaryotlarda promotör bölge; başlangıç noktasının (+1) önünde • -80 konumunda CAAT dizisi, • -30 konumunda TATA dizisi • Bazı genlerde GC elemanı • Ökaryotik RNA polimerazların transkripsiyonu başlatmak için transkripsiyon faktörleri denen özel proteinlere ihtiyacı vardır.
Ökaryotlarda promotör bölge • Ökaryotlarda promotör bölge; başlangıç noktasının (+1) önünde • -80 konumunda CAAT dizisi, • -30 konumunda TATA dizisi • Bazı genlerde GC elemanı • Ökaryotik RNA polimerazların transkripsiyonu başlatmak için transkripsiyon faktörleri denen özel proteinlere ihtiyacı vardır.
 Ökaryotik Promotör diziler • TATA kutusunun önünde verimli transkripsiyon için gerekli olan üç dizi elemanı içerir; bir CCAAT kutusu ve iki GC kutusu (konsensus dizisi GGGCGG)
Ökaryotik Promotör diziler • TATA kutusunun önünde verimli transkripsiyon için gerekli olan üç dizi elemanı içerir; bir CCAAT kutusu ve iki GC kutusu (konsensus dizisi GGGCGG)
 ( (mutasyonu Hemofili A ya neden olur) Dokuya özgün") ( (mutasyonu hemoglobinopatilere neden olur) ( (mutasyonu Hemofili A ya neden olur) Dokuya özgün genler Hypoxanthine phosphoribosyltransferase (House keeping gen) Tipik bir insan geni yapısı
( (mutasyonu hemoglobinopatilere neden olur) ( (mutasyonu Hemofili A ya neden olur) Dokuya özgün genler Hypoxanthine phosphoribosyltransferase (House keeping gen) Tipik bir insan geni yapısı
 Transkripsiyon faktörleri • Transkripsiyon faktörleri, 2 gruba ayrılır: • 1 - Genel Transkripsiyon Faktörleri • 2 - Gene Özgü transkripsiyon faktörleri, Özel genlerin ekspresyonunu denetler. • Genomdaki genlerin %5’ i, transkripsiyon faktörü kodlar.
Transkripsiyon faktörleri • Transkripsiyon faktörleri, 2 gruba ayrılır: • 1 - Genel Transkripsiyon Faktörleri • 2 - Gene Özgü transkripsiyon faktörleri, Özel genlerin ekspresyonunu denetler. • Genomdaki genlerin %5’ i, transkripsiyon faktörü kodlar.
 Transkripsiyon faktörleri • Promotör Diziler RNA polimeraz ve Transkripsiyon faktörleri tarafından tanınır • Bu faktörler, birlikte çalıştığı RNA polimeraz enzimine göre adlandırılırlar. • TF-I (RNA Polimeraz I) • TF-II (RNA Polimeraz II) • TF-III (RNA Polimeraz III)
Transkripsiyon faktörleri • Promotör Diziler RNA polimeraz ve Transkripsiyon faktörleri tarafından tanınır • Bu faktörler, birlikte çalıştığı RNA polimeraz enzimine göre adlandırılırlar. • TF-I (RNA Polimeraz I) • TF-II (RNA Polimeraz II) • TF-III (RNA Polimeraz III)
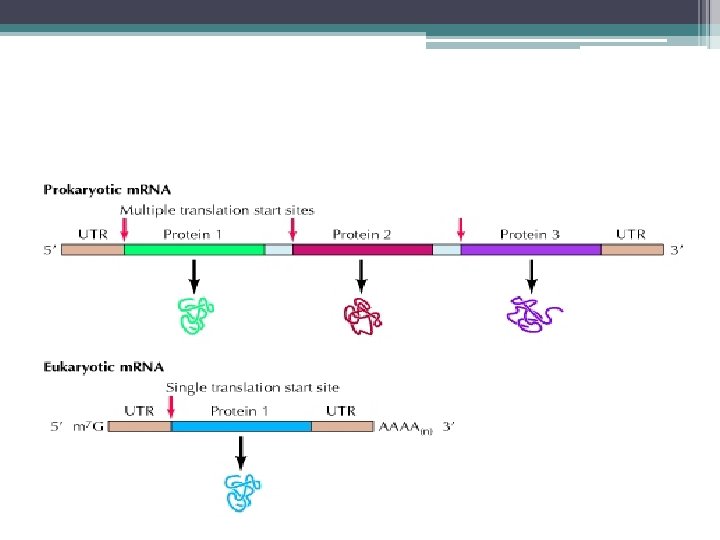
 m. RNA sentezi • Ökaryotik genler; ▫ ▫ ▫ Düzenleyici bölge Promotör Bölge Başlangıç Sinyali Eksonlar İntronlar Sonlanma Sinyalleri’ nden Oluşur.
m. RNA sentezi • Ökaryotik genler; ▫ ▫ ▫ Düzenleyici bölge Promotör Bölge Başlangıç Sinyali Eksonlar İntronlar Sonlanma Sinyalleri’ nden Oluşur.
 Transkripsiyonun başlaması • RNA Polimeraz II ve transkripsiyon faktörleri tarafından promotör bölgelerin tanınması ile başlar. • 5 genel transkripsiyon faktörüne ihtiyaç vardır. ▫ ▫ ▫ TF II A, TF II B, TF II D, TF II E, TF II F, TF II H
Transkripsiyonun başlaması • RNA Polimeraz II ve transkripsiyon faktörleri tarafından promotör bölgelerin tanınması ile başlar. • 5 genel transkripsiyon faktörüne ihtiyaç vardır. ▫ ▫ ▫ TF II A, TF II B, TF II D, TF II E, TF II F, TF II H
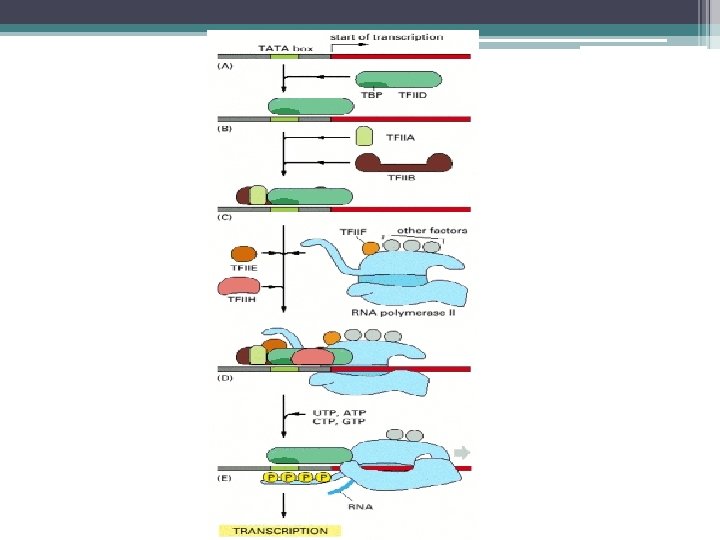
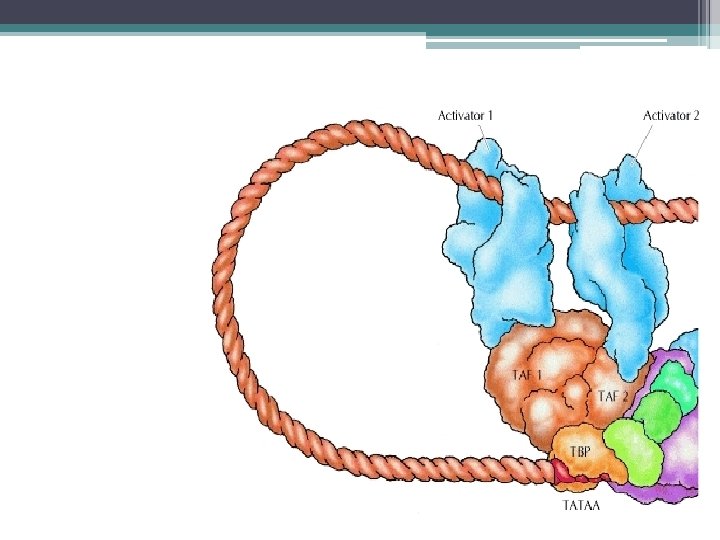
; TATA dizisine bağlanır.") Transkripsiyonun başlaması 1 - Önce TF II D (genel transkripsiyon faktörü); TATA dizisine bağlanır. (TF II D ; TBP (TATA ya bağlanan protein) ve TAF lardan (TBP e bağlanan faktörler) oluşan 10 polipeptid içeren bir kompleksdir. ) 2 - Sonra, TF IIB, TBP ye bağlanır. 3 - TF IIF ile birlikte polimeraz bağlanır 4 - En son olarak TF IIE ve TF IIH komplekse eklenir.
Transkripsiyonun başlaması 1 - Önce TF II D (genel transkripsiyon faktörü); TATA dizisine bağlanır. (TF II D ; TBP (TATA ya bağlanan protein) ve TAF lardan (TBP e bağlanan faktörler) oluşan 10 polipeptid içeren bir kompleksdir. ) 2 - Sonra, TF IIB, TBP ye bağlanır. 3 - TF IIF ile birlikte polimeraz bağlanır 4 - En son olarak TF IIE ve TF IIH komplekse eklenir.
 Transkripsiyonun başlaması • TFII H, çok alt birimli bir faktördür. • TFII H nin 2 alt birimi, başlama yeri yakınında DNA yı çözen helikazlardır. • TFII H nin diğer alt birimi, Polimeraz II nin C ucu (CTD) bölgesindeki tekrarlayan dizileri fosforilleyen bir protein kinazdır.
Transkripsiyonun başlaması • TFII H, çok alt birimli bir faktördür. • TFII H nin 2 alt birimi, başlama yeri yakınında DNA yı çözen helikazlardır. • TFII H nin diğer alt birimi, Polimeraz II nin C ucu (CTD) bölgesindeki tekrarlayan dizileri fosforilleyen bir protein kinazdır.
 • CTD; 7 Aminoasit tekrarından oluşur. (52 tekrar) •") RNA Polimerazın karboksi ucu (CTD) • CTD; 7 Aminoasit tekrarından oluşur. (52 tekrar) • CTD fosfatlanması, polimerazı başlama kompleksinden ayırır. • Böylece Polimerazın transkripsiyonu başlatmasını sağlayan diğer proteinler bağlanabilir. • Fosforile olmuş CTD m. RNA sentezi ve işlenmesine katılan enzim kompleksleri, uzama ve işleme faktörleri için bağlanma bölgesi oluşturur.
RNA Polimerazın karboksi ucu (CTD) • CTD; 7 Aminoasit tekrarından oluşur. (52 tekrar) • CTD fosfatlanması, polimerazı başlama kompleksinden ayırır. • Böylece Polimerazın transkripsiyonu başlatmasını sağlayan diğer proteinler bağlanabilir. • Fosforile olmuş CTD m. RNA sentezi ve işlenmesine katılan enzim kompleksleri, uzama ve işleme faktörleri için bağlanma bölgesi oluşturur.
 Transkripsiyonun başlaması • TF IIH nin Helikaz etkinliği ile DNA 2 ipliği birbirinden ayrılır. TF IIH ın protein kinaz etkinliği ile • RNA Polimerazın karboksi ucu (CTD) fosforillenince, transkripsiyon başlar. • Sentez başlayınca tüm faktörler RNA pol II den ayrılır • Sentez tamamlanınca hem ekson hem de intronları içeren öncül m. RNA (pre-m. RNA) oluşur.
Transkripsiyonun başlaması • TF IIH nin Helikaz etkinliği ile DNA 2 ipliği birbirinden ayrılır. TF IIH ın protein kinaz etkinliği ile • RNA Polimerazın karboksi ucu (CTD) fosforillenince, transkripsiyon başlar. • Sentez başlayınca tüm faktörler RNA pol II den ayrılır • Sentez tamamlanınca hem ekson hem de intronları içeren öncül m. RNA (pre-m. RNA) oluşur.
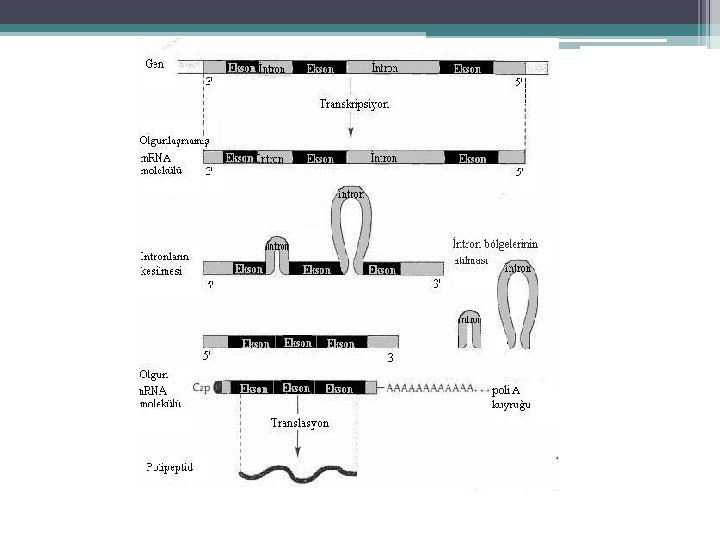
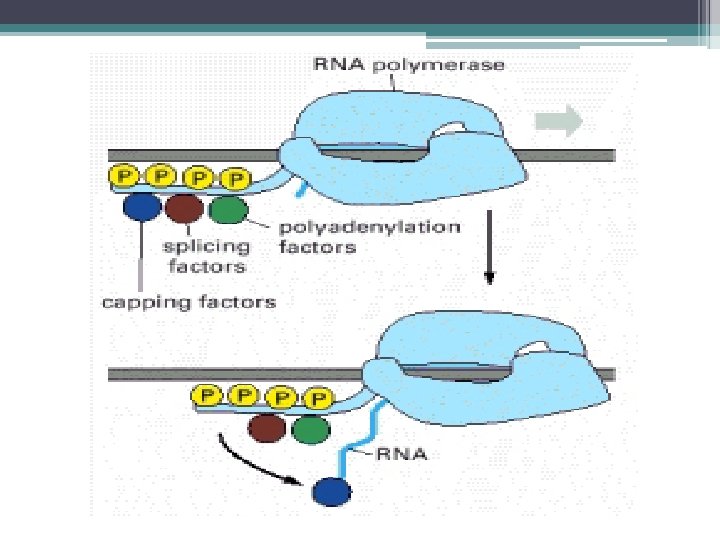
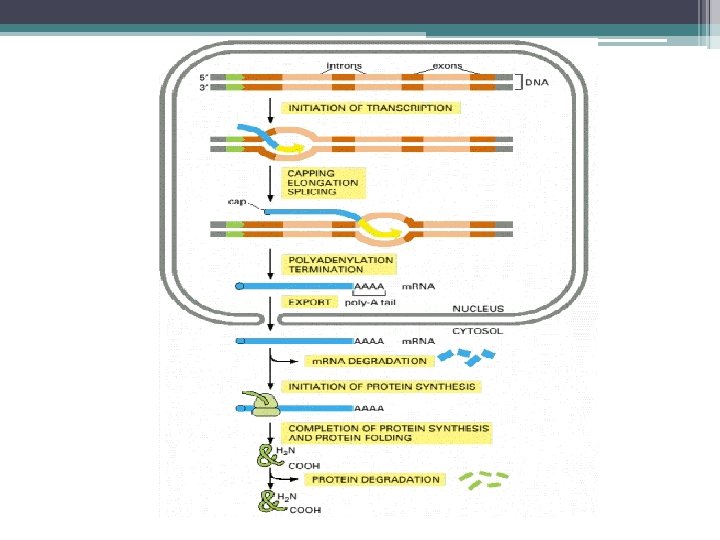
 m. RNA işlenmesi • Sonraki Aşamalar: ▫ Primer transkriptin 5’ ucuna CAP grubunun eklenmesi ▫ 3’ ucuna Poli-A yapısının eklenmesi ▫ Kırpılma (Splicing) İntronların kesilip çıkartılması, eksonların bir araya getirilmesi. ▫ Kimyasal değişiklikler (metillenme) ▫ m. RNA transkriptin 5’ ucunda metile olmuş CAP yapısı 3’ ucunda Poli-A dizisi bulunur.
m. RNA işlenmesi • Sonraki Aşamalar: ▫ Primer transkriptin 5’ ucuna CAP grubunun eklenmesi ▫ 3’ ucuna Poli-A yapısının eklenmesi ▫ Kırpılma (Splicing) İntronların kesilip çıkartılması, eksonların bir araya getirilmesi. ▫ Kimyasal değişiklikler (metillenme) ▫ m. RNA transkriptin 5’ ucunda metile olmuş CAP yapısı 3’ ucunda Poli-A dizisi bulunur.
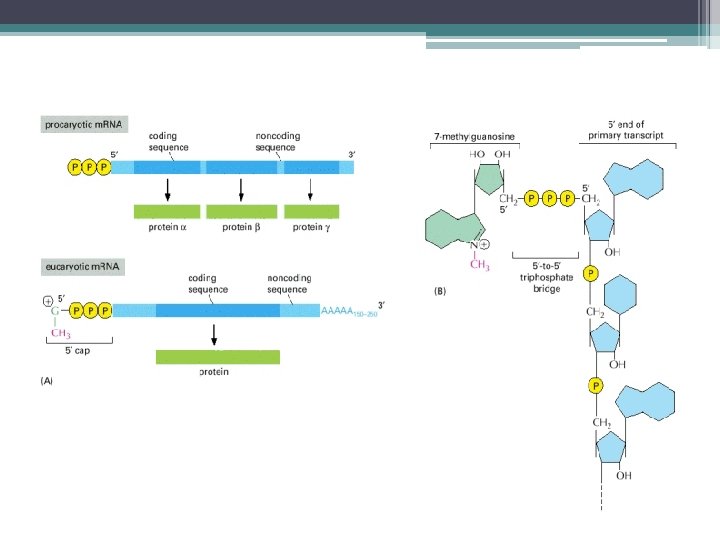
 bölgesi • m. RNA nın 5’") Ökaryot m. RNA ların 5’ ucunda, kapsül (CAP) bölgesi • m. RNA nın 5’ ve 3’ terminal kısmında translasyonu yapılmayan diziler bulunur (3’-UTR ve 5’-UTR) • Ökaryot m. RNA ların 5’ ucunda, kapsül veya başlık denilen bir bölge vardır. • Bu yapının protein sentezinin başlangıç aşamasında Ribozomun bağlanma bölgesinin (AUG başlangıç kodonu) belirlenmesinde iş gördüğü • m. RNA yı ekzonukleaz yıkımından koruduğu düşünülmekte.
Ökaryot m. RNA ların 5’ ucunda, kapsül (CAP) bölgesi • m. RNA nın 5’ ve 3’ terminal kısmında translasyonu yapılmayan diziler bulunur (3’-UTR ve 5’-UTR) • Ökaryot m. RNA ların 5’ ucunda, kapsül veya başlık denilen bir bölge vardır. • Bu yapının protein sentezinin başlangıç aşamasında Ribozomun bağlanma bölgesinin (AUG başlangıç kodonu) belirlenmesinde iş gördüğü • m. RNA yı ekzonukleaz yıkımından koruduğu düşünülmekte.
 bölgesinin eklenmesi İşlenmede ilk basamak; •") Ökaryot m. RNA ların 5’ ucuna, başlık (CAP) bölgesinin eklenmesi İşlenmede ilk basamak; • Transkriptin 5’ ucuna 7 metilguanozin başlık denen yapının eklenmesidir (m 7 G) İlk 20 – 30 nükleotidin transkripsiyonundan sonra başlık eklenir. 5’ başlık • m. RNA nın 5’ ucuna ters yönde bir GTP eklenmesi ile bir 5’-5’ bağlantı oluşturulur. • Eklenen guanosin daha sonra N 7 konumunda metillenir ve • m. RNA daki ilk bir veya iki nukleotidin ribozlarına metil grupları eklenir.
Ökaryot m. RNA ların 5’ ucuna, başlık (CAP) bölgesinin eklenmesi İşlenmede ilk basamak; • Transkriptin 5’ ucuna 7 metilguanozin başlık denen yapının eklenmesidir (m 7 G) İlk 20 – 30 nükleotidin transkripsiyonundan sonra başlık eklenir. 5’ başlık • m. RNA nın 5’ ucuna ters yönde bir GTP eklenmesi ile bir 5’-5’ bağlantı oluşturulur. • Eklenen guanosin daha sonra N 7 konumunda metillenir ve • m. RNA daki ilk bir veya iki nukleotidin ribozlarına metil grupları eklenir.
 başlık ile") Ökaryot m. RNAlarının işlenmesi 5’ ucunun 7 -metil guanosin (m 7 G) başlık ile ve 3’ ucunun poliadenillenme ile değiştirilmesini, ayrıca İntronların kesilip çıkarılmasını kapsar. 5’ başlık m. RNA nın 5’ ucuna ters yönde bir GTP eklenmesi ile Bir 5’-5’ bağlantı oluşturulur. Eklenen guanosin daha sonra N 7 konumunda metillenir ve m. RNA daki ilk bir veya iki nukleotidin ribozlarına metil grupları eklenir.
Ökaryot m. RNAlarının işlenmesi 5’ ucunun 7 -metil guanosin (m 7 G) başlık ile ve 3’ ucunun poliadenillenme ile değiştirilmesini, ayrıca İntronların kesilip çıkarılmasını kapsar. 5’ başlık m. RNA nın 5’ ucuna ters yönde bir GTP eklenmesi ile Bir 5’-5’ bağlantı oluşturulur. Eklenen guanosin daha sonra N 7 konumunda metillenir ve m. RNA daki ilk bir veya iki nukleotidin ribozlarına metil grupları eklenir.
 Ökaryot m. RNA ların 3’ ucunda Poli-A kuyruğu vardır • 3’ ucunda çok sayıda Adeninden oluşan Poli-A kuyruğu vardır. • Poli-A polimeraz tarafından m. RNA ya eklenir. • Poli-A dizisine, Poli-A bağlayan proteinlerin bağlanması ile m. RNA 3’ ekzonukleaz yıkımından korunur. • m. RNA kararlılığını sağlar, • m. RNA ların sitoplazmaya taşınmasında görev yapar.
Ökaryot m. RNA ların 3’ ucunda Poli-A kuyruğu vardır • 3’ ucunda çok sayıda Adeninden oluşan Poli-A kuyruğu vardır. • Poli-A polimeraz tarafından m. RNA ya eklenir. • Poli-A dizisine, Poli-A bağlayan proteinlerin bağlanması ile m. RNA 3’ ekzonukleaz yıkımından korunur. • m. RNA kararlılığını sağlar, • m. RNA ların sitoplazmaya taşınmasında görev yapar.
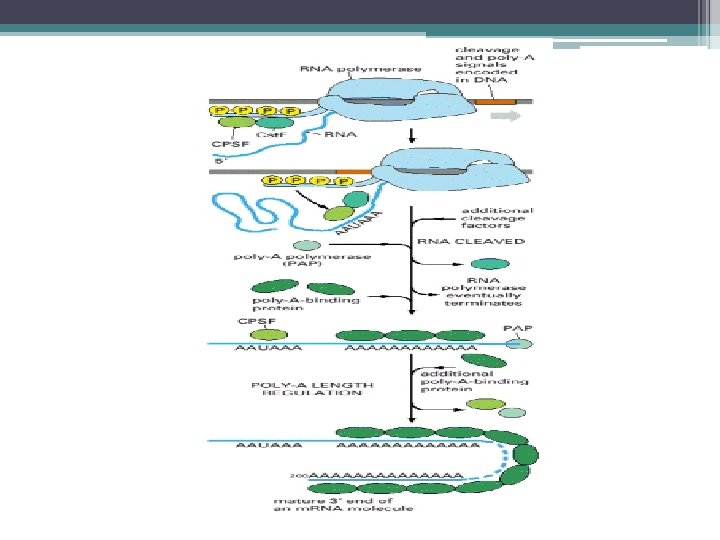
 Poli–A Kuyruğunun Eklenmesi • 3’ ucu primer transkriptin kesilmesi ve Poli-A kuyruğunun eklenmesi • Poliadenillenme sinyalleri; • Aşağıda G U dan zengin bir dizi • Yukarda korunmuş bir heksanükleotid (memelide AAUAAA) • Onun yukarısında U dan zengin bir dizi • RNA yı kesen bir endonükleaz ve • ~200 nükleotidlik bir Poli-A kuyruğu ekleyen bir • Poli-A polimeraz bu dizileri tanır.
Poli–A Kuyruğunun Eklenmesi • 3’ ucu primer transkriptin kesilmesi ve Poli-A kuyruğunun eklenmesi • Poliadenillenme sinyalleri; • Aşağıda G U dan zengin bir dizi • Yukarda korunmuş bir heksanükleotid (memelide AAUAAA) • Onun yukarısında U dan zengin bir dizi • RNA yı kesen bir endonükleaz ve • ~200 nükleotidlik bir Poli-A kuyruğu ekleyen bir • Poli-A polimeraz bu dizileri tanır.
 Ökaryot m. RNAlarının 3’ uçlarının oluşumu: Memeli hücrelerinde poliadenilasyon sinyalleri yukarı ve aşağı GU dan zengin diziler ve AAUAAA heksanukleotid dizisinden oluşur. Bir endonukleaz prem. RNA yı AAUAAA nın 10 -30 nukleotid aşağısından bir CA dizisinden keser. Sonra poli-A polimeraz ~200 A içeren poli-A kuyruğunu RNA nın 3’ ucuna ekler
Ökaryot m. RNAlarının 3’ uçlarının oluşumu: Memeli hücrelerinde poliadenilasyon sinyalleri yukarı ve aşağı GU dan zengin diziler ve AAUAAA heksanukleotid dizisinden oluşur. Bir endonukleaz prem. RNA yı AAUAAA nın 10 -30 nukleotid aşağısından bir CA dizisinden keser. Sonra poli-A polimeraz ~200 A içeren poli-A kuyruğunu RNA nın 3’ ucuna ekler
 Ökaryot m. RNA ların 3’ ucuna Poli-A kuyruğunun eklenmesi
Ökaryot m. RNA ların 3’ ucuna Poli-A kuyruğunun eklenmesi
 m. RNA nın nukleustan sitoplazmaya taşınması
m. RNA nın nukleustan sitoplazmaya taşınması
 m. RNA nın nukleustan sitoplazmaya taşınması
m. RNA nın nukleustan sitoplazmaya taşınması
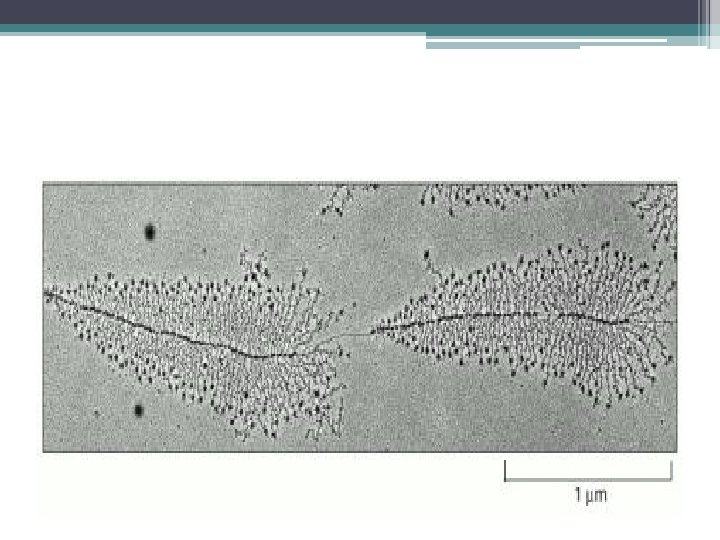
 Poli -A kuyruk uzunluğu m. RNA çevrimini denetler • Erken gelişimde Poli -A kuyruk uzunluğu m. RNA çevrimini denetler • Düzenleyici rol oynar • Döllenmemiş yumurtada kısa Poli -A kuyruklu çok sayıda m. RNA (30 – 50 nükleotid uzunlukta) proteine çevrilmeden saklanır • Döllenme depolanan m. RNA Poli -A kuyruklarının uzamasını uyarır • Protein sentezini aktive eder
Poli -A kuyruk uzunluğu m. RNA çevrimini denetler • Erken gelişimde Poli -A kuyruk uzunluğu m. RNA çevrimini denetler • Düzenleyici rol oynar • Döllenmemiş yumurtada kısa Poli -A kuyruklu çok sayıda m. RNA (30 – 50 nükleotid uzunlukta) proteine çevrilmeden saklanır • Döllenme depolanan m. RNA Poli -A kuyruklarının uzamasını uyarır • Protein sentezini aktive eder
") Intronların Kesilip Çıkarılması; Kırpılma (Splicing)
Intronların Kesilip Çıkarılması; Kırpılma (Splicing)
 İn vitro kesip ekleme İntron içeren bir gen bir bakteriyofaj RNA polimerazı tarafından tanınan promotörün aşağı bölgesinde klonlanır. Plazmid, eklenen genin 3’ ucundan kesim yapan bir restriksiyon enzimi ile kesilerek, doğrusal bir DNA Molekülü oluşturulur. Daha sonra bu DNA, bakteriyofaj polimerazı ile in vitro olarak kopyalanarak pre-m. RNA üretilir. Bu pre-m. RNA memeli hücrelerinin nuklear ekstraktlarına eklenerek kırpılma(kesip ekleme) Reaksiyonları in vitro olarak çalışılabilir
İn vitro kesip ekleme İntron içeren bir gen bir bakteriyofaj RNA polimerazı tarafından tanınan promotörün aşağı bölgesinde klonlanır. Plazmid, eklenen genin 3’ ucundan kesim yapan bir restriksiyon enzimi ile kesilerek, doğrusal bir DNA Molekülü oluşturulur. Daha sonra bu DNA, bakteriyofaj polimerazı ile in vitro olarak kopyalanarak pre-m. RNA üretilir. Bu pre-m. RNA memeli hücrelerinin nuklear ekstraktlarına eklenerek kırpılma(kesip ekleme) Reaksiyonları in vitro olarak çalışılabilir
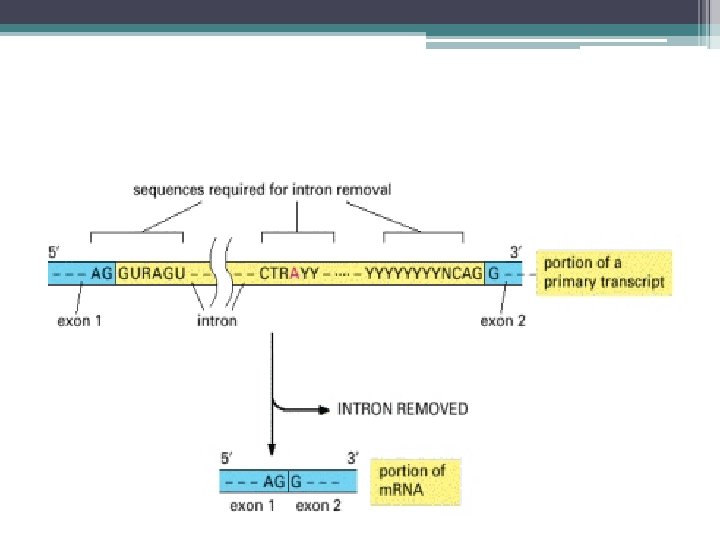
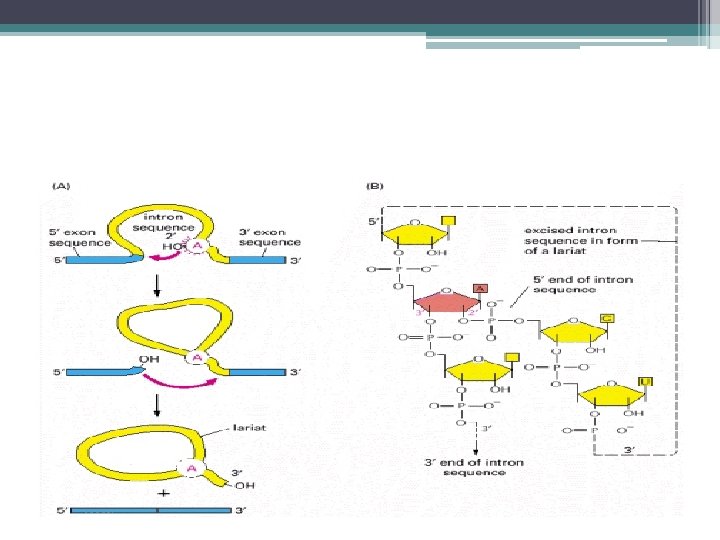
 • Pre-m. RNA kırpılması 2 aşamada gerçekleşir; 1 -") Intronların Kesilip Çıkarılması; Kırpılma (Splicing) • Pre-m. RNA kırpılması 2 aşamada gerçekleşir; 1 - pre-m. RNA nın 5’ kesim noktasından (SS) (GU) kesilmesi ve • intronun kesilen 5’ ucunun intron içindeki bir Adenin ile (dallanma noktası) birleştirilmesi 2 - İntronun 3’ kesim noktasından kesilmesi (AG) ve • eş zamanlı olarak eksonların birleştirilmesi • intronun ilmek benzeri bir yapı şeklinde çıkarılması ve nukleusta yıkılması
Intronların Kesilip Çıkarılması; Kırpılma (Splicing) • Pre-m. RNA kırpılması 2 aşamada gerçekleşir; 1 - pre-m. RNA nın 5’ kesim noktasından (SS) (GU) kesilmesi ve • intronun kesilen 5’ ucunun intron içindeki bir Adenin ile (dallanma noktası) birleştirilmesi 2 - İntronun 3’ kesim noktasından kesilmesi (AG) ve • eş zamanlı olarak eksonların birleştirilmesi • intronun ilmek benzeri bir yapı şeklinde çıkarılması ve nukleusta yıkılması
(GU) kesilmesi ve") Pre-m. RNA nın kırpılması : 1 -pre-m. RNA nın 5’kesim noktasından (SS)(GU) kesilmesi ve intronun kesilen 5’ucunun intron içindeki bir Adenin ile (dallanma noktası) birleştirilmesi 2 - İntronun 3’ kesim noktasından kesilmesi (AG) ve eş zamanlı olarak eksonların birleştirilmesi intronun ilmek benzeri bir yapı şeklinde çıkarılması ve nukleusta yıkılması
Pre-m. RNA nın kırpılması : 1 -pre-m. RNA nın 5’kesim noktasından (SS)(GU) kesilmesi ve intronun kesilen 5’ucunun intron içindeki bir Adenin ile (dallanma noktası) birleştirilmesi 2 - İntronun 3’ kesim noktasından kesilmesi (AG) ve eş zamanlı olarak eksonların birleştirilmesi intronun ilmek benzeri bir yapı şeklinde çıkarılması ve nukleusta yıkılması
 Pre-m. RNA ların kırpılma işlemi için gerekli olan 3 önemli dizi • 5’ kesim noktası (SS) dizileri (GU) • 3’ kesim noktası dizileri (AG) • İntron içinde dallanma noktasındaki diziler (adenin) kement benzeri yapıyı oluşturmak üzere intronun 5’ ucunun bağlandığı nokta
Pre-m. RNA ların kırpılma işlemi için gerekli olan 3 önemli dizi • 5’ kesim noktası (SS) dizileri (GU) • 3’ kesim noktası dizileri (AG) • İntron içinde dallanma noktasındaki diziler (adenin) kement benzeri yapıyı oluşturmak üzere intronun 5’ ucunun bağlandığı nokta
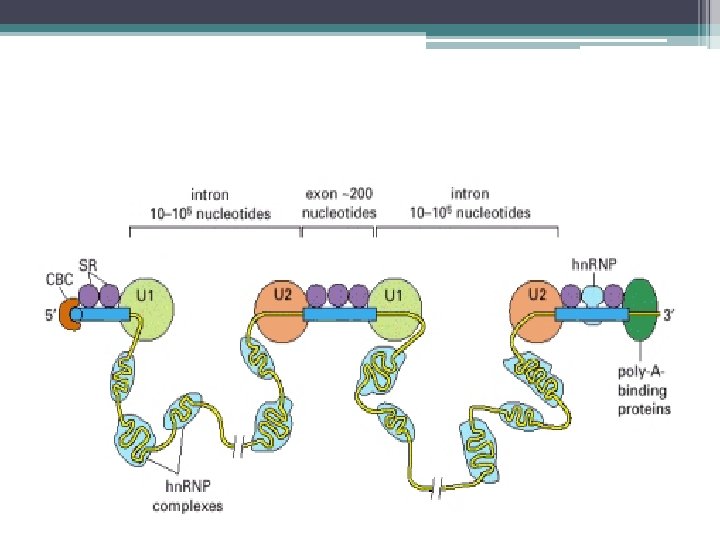
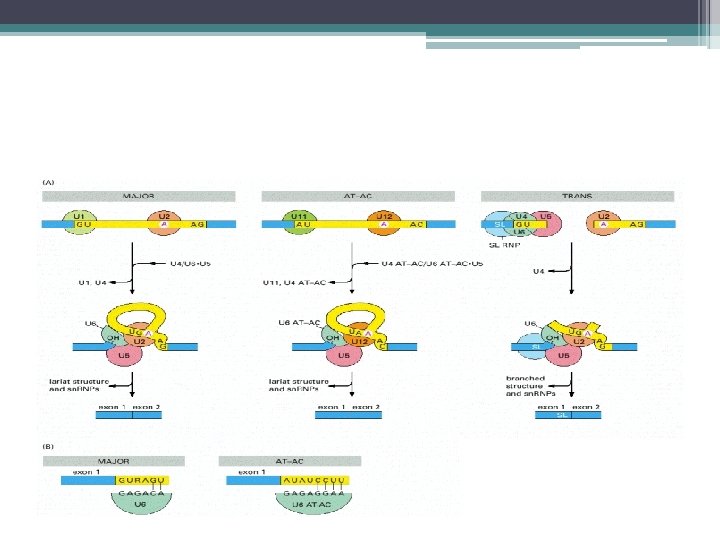
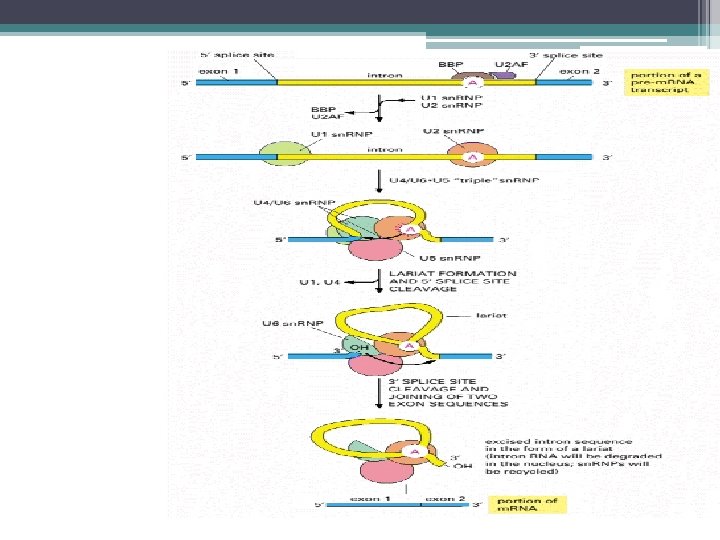
 Kırpılma işlemi • Kırpılma işlemi; • Splisozom denilen RNA+protein kompleksi ile yapılır. • RNA bileşenleri U 1 , U 2 , U 4 , U 5 , U 6 denen 5 adet küçük nüklear RNA lardır (sn. RNA) • 50 – 200 nükleotidlik sn. RNA lar • 6 – 10 protein molekülü ile kompleks yaparak= küçük nüklear ribonukleoprotein taneciklerini (sn. RNP) oluşturur.
Kırpılma işlemi • Kırpılma işlemi; • Splisozom denilen RNA+protein kompleksi ile yapılır. • RNA bileşenleri U 1 , U 2 , U 4 , U 5 , U 6 denen 5 adet küçük nüklear RNA lardır (sn. RNA) • 50 – 200 nükleotidlik sn. RNA lar • 6 – 10 protein molekülü ile kompleks yaparak= küçük nüklear ribonukleoprotein taneciklerini (sn. RNP) oluşturur.
 Splisozomun Oluşumu ve Kırpılma • U 1 sn. RNP ler pre-m. RNA nın 5’ kesim bölgesine (GU) bağlanır. • U 2 sn. RNP ler intronun dallanma noktası dizisine bağlanır • U 4/U 6 ve U 5 sn. RNP kompleksi splisozoma katılır • U 5; 5’ kesim noktasının öncesindeki dizilere bağlanır • U 6, U 4 den ayrılır ve 5’ kesim bölgesindeki U 1 in yerine bağlanır, • U 1 ayrılır • U 5; 3’ kesim bölgesine de bağlanır
Splisozomun Oluşumu ve Kırpılma • U 1 sn. RNP ler pre-m. RNA nın 5’ kesim bölgesine (GU) bağlanır. • U 2 sn. RNP ler intronun dallanma noktası dizisine bağlanır • U 4/U 6 ve U 5 sn. RNP kompleksi splisozoma katılır • U 5; 5’ kesim noktasının öncesindeki dizilere bağlanır • U 6, U 4 den ayrılır ve 5’ kesim bölgesindeki U 1 in yerine bağlanır, • U 1 ayrılır • U 5; 3’ kesim bölgesine de bağlanır
 Splisozomun yapılanması • İlk basamak U 1 sn. RNP lerin pre-m. RNA nın 5’ kesim bölgesine (GU) bağlanması. • Sonra U 2 sn. RNP lerin intronun dallanma noktasına bağlanması. • U 4/U 6 ve U 5 sn. RNP leri içeren kompleksin splisozoma katılması. • U 5, 5’ kesim noktasının yukarısındaki dizilere bağlanır ve U 6/ U 4 kement benzeri yapı oluşmadan önce U 1 ile yer değiştirir U 1 ayrılır. • Daha sonra U 5 , 3’ kesim bölgesine bağlanır • İntronun çıkartılması ve eksonların birleştirilmesi
Splisozomun yapılanması • İlk basamak U 1 sn. RNP lerin pre-m. RNA nın 5’ kesim bölgesine (GU) bağlanması. • Sonra U 2 sn. RNP lerin intronun dallanma noktasına bağlanması. • U 4/U 6 ve U 5 sn. RNP leri içeren kompleksin splisozoma katılması. • U 5, 5’ kesim noktasının yukarısındaki dizilere bağlanır ve U 6/ U 4 kement benzeri yapı oluşmadan önce U 1 ile yer değiştirir U 1 ayrılır. • Daha sonra U 5 , 3’ kesim bölgesine bağlanır • İntronun çıkartılması ve eksonların birleştirilmesi
 Splisozomun Oluşumu ve Kırpılma • Kement benzeri bir yapı oluşur • Intron kesilip çıkarılır • Eksonlar birleştirilir • sn. RNA lar; 1 - pre m. RNA ların kesim bölgelerini ve dallanma noktalarındaki ortak dizileri tanır 2 - Kesilip ekleme reaksiyonunu katalizlerler
Splisozomun Oluşumu ve Kırpılma • Kement benzeri bir yapı oluşur • Intron kesilip çıkarılır • Eksonlar birleştirilir • sn. RNA lar; 1 - pre m. RNA ların kesim bölgelerini ve dallanma noktalarındaki ortak dizileri tanır 2 - Kesilip ekleme reaksiyonunu katalizlerler
 U 1 sn RNA nın 5’ kesim bölgesine bağlanması U 1 sn RNA nın 5’ ucu, komplementer baz eşleşmesi ile 5’ kesim noktasındaki ortak Dizilere bağlanır.
U 1 sn RNA nın 5’ kesim bölgesine bağlanması U 1 sn RNA nın 5’ ucu, komplementer baz eşleşmesi ile 5’ kesim noktasındaki ortak Dizilere bağlanır.
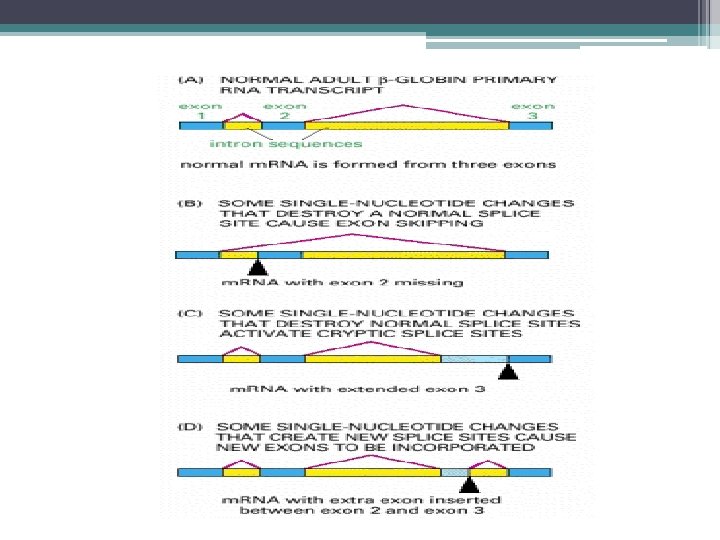
 pre-m. RNA dan, İntronların Kesilip Çıkarılması, Eksonların Biraraya Getirilmesi • Kırpılmaya katılan diziler; • İntronların 5’ ve 3’ terminalinde dinukleotid dizileri GU ve AG • Bunlarda mutasyon m. RNA kırpılmasında hatalara neden olur
pre-m. RNA dan, İntronların Kesilip Çıkarılması, Eksonların Biraraya Getirilmesi • Kırpılmaya katılan diziler; • İntronların 5’ ve 3’ terminalinde dinukleotid dizileri GU ve AG • Bunlarda mutasyon m. RNA kırpılmasında hatalara neden olur
 Gen ekspresyonunun kontrolü
Gen ekspresyonunun kontrolü
 aşamasında • m. RNA nın") Gen ekspresyonunun kontrolü • m. RNA nın transkripsiyonu (yazılımı) aşamasında • m. RNA nın işlenmesi sırasında • m. RNA nın taşınması sırasında • m. RNA nın yıkım aşamasında • Protein etkinliğinin kontrolü
Gen ekspresyonunun kontrolü • m. RNA nın transkripsiyonu (yazılımı) aşamasında • m. RNA nın işlenmesi sırasında • m. RNA nın taşınması sırasında • m. RNA nın yıkım aşamasında • Protein etkinliğinin kontrolü
 Represörler ve transkripsiyonun negatif kontrolü: • 1950 lerde Jacob ve Monod ilk çalışmalar • E. coli’de • Laktoz (yıkılarak glukoz ve galaktoza çevrilir) metabolizmasına katılan enzimlerin ekspresyonunu incelediler • Laktozun yıkılması; • Yıkımı katalizleyen β- galaktozidaz enzimi ve diğer iki enzim (laktoz permeaz ve transasetilaz) ortamda laktoz varsa ifade edilir. • Laktoz kendi metabolizmasında yer alan enzimlerin ekspresyonunu uyarır. • E. coli’de Mutantları tanımladılar. O ve i geni mutantları; sürekli ekspresyon yapan veya uyarılamayan
Represörler ve transkripsiyonun negatif kontrolü: • 1950 lerde Jacob ve Monod ilk çalışmalar • E. coli’de • Laktoz (yıkılarak glukoz ve galaktoza çevrilir) metabolizmasına katılan enzimlerin ekspresyonunu incelediler • Laktozun yıkılması; • Yıkımı katalizleyen β- galaktozidaz enzimi ve diğer iki enzim (laktoz permeaz ve transasetilaz) ortamda laktoz varsa ifade edilir. • Laktoz kendi metabolizmasında yer alan enzimlerin ekspresyonunu uyarır. • E. coli’de Mutantları tanımladılar. O ve i geni mutantları; sürekli ekspresyon yapan veya uyarılamayan
 E. Colide transkripsiyonun negatif kontrolü: • Lac operonu negatif kontrolü • Lac operonu denen üç genden oluşan tek birim; (z; beta Galaktozidaz, y; permeaz, a; transasetilaz) • İ geni ; transkripsiyonu engelleyen bir represörü kodlar , laktoz represöre bağlanır ve represörün operatöre bağlanmasını önler.
E. Colide transkripsiyonun negatif kontrolü: • Lac operonu negatif kontrolü • Lac operonu denen üç genden oluşan tek birim; (z; beta Galaktozidaz, y; permeaz, a; transasetilaz) • İ geni ; transkripsiyonu engelleyen bir represörü kodlar , laktoz represöre bağlanır ve represörün operatöre bağlanmasını önler.
 E. Colide transkripsiyonun negatif kontrolü: • represör laktoz yokluğunda operatöre bağlanır ve polimerazın bu üç genin transkripsiyonunu yapmak üzere promotöre bağlanmasını önler. • ortamda Laktoz varsa laktoz represöre bağlanır ve represörün operatöre bağlanmasını önler. • Böylece operonun transkripsiyonu uyarılır.
E. Colide transkripsiyonun negatif kontrolü: • represör laktoz yokluğunda operatöre bağlanır ve polimerazın bu üç genin transkripsiyonunu yapmak üzere promotöre bağlanmasını önler. • ortamda Laktoz varsa laktoz represöre bağlanır ve represörün operatöre bağlanmasını önler. • Böylece operonun transkripsiyonu uyarılır.
 E. Colide transkripsiyonun negatif kontrolü: Lac operonu negatif kontrolü Lac operonu denen üç genden oluşan tek birim; (z; beta. Galaktozidaz, y; permeaz, a; transasetilaz) İ geni laktoz yokluğunda operatöre bağlanan ve polimerazın bu üç genin transkripsiyonunu yapmak üzere promotöre bağlanmasını önleyen bir represörü kodlar. Laktoz ortamda mevcut ise represöre bağlanır ve represörün operatöre bağlanmasını önler. Böylece operonun transkripsiyonu uyarılır. !960 larda protein izole edildi operator DNA ya bağlandığı gösterildi.
E. Colide transkripsiyonun negatif kontrolü: Lac operonu negatif kontrolü Lac operonu denen üç genden oluşan tek birim; (z; beta. Galaktozidaz, y; permeaz, a; transasetilaz) İ geni laktoz yokluğunda operatöre bağlanan ve polimerazın bu üç genin transkripsiyonunu yapmak üzere promotöre bağlanmasını önleyen bir represörü kodlar. Laktoz ortamda mevcut ise represöre bağlanır ve represörün operatöre bağlanmasını önler. Böylece operonun transkripsiyonu uyarılır. !960 larda protein izole edildi operator DNA ya bağlandığı gösterildi.
 Lac operonu çalışmaları • Sonuç: • Transkripsiyonun • DNA dizileri ile • regülatör proteinlerin etkileşmesi yoluyla denetlendiğinin gösterilmesi
Lac operonu çalışmaları • Sonuç: • Transkripsiyonun • DNA dizileri ile • regülatör proteinlerin etkileşmesi yoluyla denetlendiğinin gösterilmesi
 E. coli’de transkripsiyonun Pozitif kontrolü: • Glukoz, diğer şekerlerin yıkılmasında görevli enzimleri kodlayan genlerin ekspresyonunu kontrol eder • Glukoz varsa glukoz kullanılır diğer sekerlerin yıkım yollarındaki enzimler sentezlenmez. Örn. Ortamda hem Glukoz hem de laktoz varsa lac operonu uyarılmaz. (laktoz varlığında bile) • bakteri glukoz kullanır • Glukoz lac operonunu baskılar.
E. coli’de transkripsiyonun Pozitif kontrolü: • Glukoz, diğer şekerlerin yıkılmasında görevli enzimleri kodlayan genlerin ekspresyonunu kontrol eder • Glukoz varsa glukoz kullanılır diğer sekerlerin yıkım yollarındaki enzimler sentezlenmez. Örn. Ortamda hem Glukoz hem de laktoz varsa lac operonu uyarılmaz. (laktoz varlığında bile) • bakteri glukoz kullanır • Glukoz lac operonunu baskılar.
 E. coli’de transkripsiyonun Pozitif kontrolü: • Glukoz lac operonunu baskılar. • Bu baskılama c. AMP düzeyine bağlı pozitif kontrol • Düşük glukoz düzeyi adenil siklaz enzimini aktifler • . c. AMP sentezi artar • c. AMP katabolit aktivatör protein (CAP) denen transkripsiyon düzenleyici proteine bağlanır. • c. AMP+CAP kompleksi Lac operonu içindeki hedef DNA dizisine bağlanır.
E. coli’de transkripsiyonun Pozitif kontrolü: • Glukoz lac operonunu baskılar. • Bu baskılama c. AMP düzeyine bağlı pozitif kontrol • Düşük glukoz düzeyi adenil siklaz enzimini aktifler • . c. AMP sentezi artar • c. AMP katabolit aktivatör protein (CAP) denen transkripsiyon düzenleyici proteine bağlanır. • c. AMP+CAP kompleksi Lac operonu içindeki hedef DNA dizisine bağlanır.
 E. coli’de transkripsiyonun Pozitif kontrolü: • c. AMP+CAP kompleksi Lac operonu içindeki hedef DNA dizisine bağlanır. • Bu dizi transkripsiyon başlama yerinin 60 baz yukarısındadır. • CAP, RNA polimerazın alfa altbirimi ile etkileşime girer. • Polimeraz promotöre bağlanır • Transkripsiyon aktive olur
E. coli’de transkripsiyonun Pozitif kontrolü: • c. AMP+CAP kompleksi Lac operonu içindeki hedef DNA dizisine bağlanır. • Bu dizi transkripsiyon başlama yerinin 60 baz yukarısındadır. • CAP, RNA polimerazın alfa altbirimi ile etkileşime girer. • Polimeraz promotöre bağlanır • Transkripsiyon aktive olur
 E. coli’de transkripsiyonun Pozitif kontrolü: Düşük glukoz düzeyi adenil siklaz enzimini aktifler. c. AMP sentezi artar • c. AMP katabolit aktivatör protein (CAP) denen transkripsiyon düzenleyici proteine bağlanır. • c. AMP+CAP kompleksi Lac operonu içindeki hedef DNA dizisine bağlanır. Bu dizi transkripsiyon başlama yerinin 60 baz yukarısındadır. • CAP, RNA polimerazın alfa altbirimi ile Etkileşime girer. Polimeraz promotöre Bağlanır. Transkripsiyon aktive olur
E. coli’de transkripsiyonun Pozitif kontrolü: Düşük glukoz düzeyi adenil siklaz enzimini aktifler. c. AMP sentezi artar • c. AMP katabolit aktivatör protein (CAP) denen transkripsiyon düzenleyici proteine bağlanır. • c. AMP+CAP kompleksi Lac operonu içindeki hedef DNA dizisine bağlanır. Bu dizi transkripsiyon başlama yerinin 60 baz yukarısındadır. • CAP, RNA polimerazın alfa altbirimi ile Etkileşime girer. Polimeraz promotöre Bağlanır. Transkripsiyon aktive olur
 Ökaryot Gen Ekspresyonunun Kontrolü • Transkripsiyonun başlangıç aşamasında kontrol; • Bakterilerdeki gibi düzenleyici dizilere bağlanan ve RNA polimerazın etkinliğini düzenleyen proteinler tarafından kontrol edilir • Prokaryotlardan farklı olarak Ökaryotlarda kromatin yapısı değişiklikleri Transkripsiyon kontrolünde anahtar rol oynar. • Başlangıç dizisinin ön tarafında bulunan çok sayıdaki regülatör diziler transkripsiyonun kontrolünden sorumludur.
Ökaryot Gen Ekspresyonunun Kontrolü • Transkripsiyonun başlangıç aşamasında kontrol; • Bakterilerdeki gibi düzenleyici dizilere bağlanan ve RNA polimerazın etkinliğini düzenleyen proteinler tarafından kontrol edilir • Prokaryotlardan farklı olarak Ökaryotlarda kromatin yapısı değişiklikleri Transkripsiyon kontrolünde anahtar rol oynar. • Başlangıç dizisinin ön tarafında bulunan çok sayıdaki regülatör diziler transkripsiyonun kontrolünden sorumludur.
 Ökaryotlarda Transkripsiyon • Prokaryot ve Ökaryot Hücreler Arasındaki Farklar: 1 - Bakteride tek bir RNA polimeraz (tüm genlerin transkripsiyonundan sorumlu) ▫ Ökaryotik hücrelerde, birden fazla farklı RNA polimerazlar bulunur. 2 - Ökaryotik RNA polimerazlar promotör diziye, ek proteinler ile bağlanır. Transkripsiyon daha karmaşık Daha gelişmiş gen ekspresyonu regülasyonu gerekli
Ökaryotlarda Transkripsiyon • Prokaryot ve Ökaryot Hücreler Arasındaki Farklar: 1 - Bakteride tek bir RNA polimeraz (tüm genlerin transkripsiyonundan sorumlu) ▫ Ökaryotik hücrelerde, birden fazla farklı RNA polimerazlar bulunur. 2 - Ökaryotik RNA polimerazlar promotör diziye, ek proteinler ile bağlanır. Transkripsiyon daha karmaşık Daha gelişmiş gen ekspresyonu regülasyonu gerekli
 Ökaryotik RNA Polimerazları ▫ 3 farklı Nükleer RNA Polimeraz bulunur ▫ RNA Polimeraz III 1 -m. RNA’ lar ; RNA Polimeraz II ile 2 -r. RNA’ lar ve t. RNA’ lar ; RNA Polimeraz I ve III ile (Pol I 28 s, 18 s, 5. 8 s r. RNA ların; Pol III 5 s r. RNA ların sentezi) 3 -Küçük RNA lar; sn. RNA (küçük nükleer RNA ; kesip ekleme ) , sc. RNA [küçük sitoplazmik RNA; protein taşınması) RNA Pol III ve RNA Pol II tarafından sentezlenir.
Ökaryotik RNA Polimerazları ▫ 3 farklı Nükleer RNA Polimeraz bulunur ▫ RNA Polimeraz III 1 -m. RNA’ lar ; RNA Polimeraz II ile 2 -r. RNA’ lar ve t. RNA’ lar ; RNA Polimeraz I ve III ile (Pol I 28 s, 18 s, 5. 8 s r. RNA ların; Pol III 5 s r. RNA ların sentezi) 3 -Küçük RNA lar; sn. RNA (küçük nükleer RNA ; kesip ekleme ) , sc. RNA [küçük sitoplazmik RNA; protein taşınması) RNA Pol III ve RNA Pol II tarafından sentezlenir.
 önünde • -30 konumunda") Ökaryotlarda promotör bölge • Ökaryotlarda promotör bölge; başlangıç noktasının (+1) önünde • -30 konumunda TATA dizisi (genel transkripsiyon faktörleri için bağlanma bölgeleri) • -80 konumunda CAAT dizisi, • Bazı genlerde GC elemanı • Ökaryotik RNA polimerazların transkripsiyonu başlatmak için transkripsiyon faktörleri denen özel proteinlere ihtiyacı vardır.
Ökaryotlarda promotör bölge • Ökaryotlarda promotör bölge; başlangıç noktasının (+1) önünde • -30 konumunda TATA dizisi (genel transkripsiyon faktörleri için bağlanma bölgeleri) • -80 konumunda CAAT dizisi, • Bazı genlerde GC elemanı • Ökaryotik RNA polimerazların transkripsiyonu başlatmak için transkripsiyon faktörleri denen özel proteinlere ihtiyacı vardır.
 Ökaryotik Promotör diziler • Herpeks simpleks timidin kinaz geni promotörü TATA kutusunun önünde verimli transkripsiyon için gerekli olan üç dizi elemanı içerir; bir CCAAT kutusu ve iki GC kutusu (konsensus dizisi GGGCGG)
Ökaryotik Promotör diziler • Herpeks simpleks timidin kinaz geni promotörü TATA kutusunun önünde verimli transkripsiyon için gerekli olan üç dizi elemanı içerir; bir CCAAT kutusu ve iki GC kutusu (konsensus dizisi GGGCGG)
 Transkripsiyonun başlaması • RNA Polimeraz II ve transkripsiyon faktörleri tarafından promotör bölgelerin tanınması ile başlar. • 5 genel transkripsiyon faktörüne ihtiyaç vardır. ▫ ▫ ▫ TF II A, TF II B, TF II D, TF II E, TF II F, TF II H
Transkripsiyonun başlaması • RNA Polimeraz II ve transkripsiyon faktörleri tarafından promotör bölgelerin tanınması ile başlar. • 5 genel transkripsiyon faktörüne ihtiyaç vardır. ▫ ▫ ▫ TF II A, TF II B, TF II D, TF II E, TF II F, TF II H
 Ökaryot regülatör dizilerin belirlenmesi Klonlanan regülatör dizi kolay belirlenen bir raportör gene bağlanır • Bu plazmid kültürdeki hücrelere transfeksiyonla verilir. • regülatör dizi raportör genin transkripsiyonunu ilerletir ve enzim varlığı hücrede belirlenir
Ökaryot regülatör dizilerin belirlenmesi Klonlanan regülatör dizi kolay belirlenen bir raportör gene bağlanır • Bu plazmid kültürdeki hücrelere transfeksiyonla verilir. • regülatör dizi raportör genin transkripsiyonunu ilerletir ve enzim varlığı hücrede belirlenir
 Regülatör diziler • Promotörler ve • Güçlendiriciler • RNA Polimeraz II ile transkripsiyonu yapılan genlerin genel transkripsiyon faktörleri için bağlanma bölgesi oluşturan • TATA kutusunu kapsayan esas promotör elemanları vardır. • TATA kutusunun yukarı bölgesinde diğer regülatör diziler yerleşmiştir. Örn. CCAAT ve GGGCGG (GC kutusu) konsensus dizileri (TATA kutusunun 100 bç yukarısında) (özel genlerin ekspresyonunu kontrol eden)
Regülatör diziler • Promotörler ve • Güçlendiriciler • RNA Polimeraz II ile transkripsiyonu yapılan genlerin genel transkripsiyon faktörleri için bağlanma bölgesi oluşturan • TATA kutusunu kapsayan esas promotör elemanları vardır. • TATA kutusunun yukarı bölgesinde diğer regülatör diziler yerleşmiştir. Örn. CCAAT ve GGGCGG (GC kutusu) konsensus dizileri (TATA kutusunun 100 bç yukarısında) (özel genlerin ekspresyonunu kontrol eden)
 Ökaryotik bir Promotör • H. Simpleks timidin kinaz geni promotörü. TATA kutusunun önünde verimli transkripsiyon için gerekli olan üç dizi elemanı içerir; bir CCAAT kutusu ve iki GC kutusu (konsensus dizisi GGGCGG)
Ökaryotik bir Promotör • H. Simpleks timidin kinaz geni promotörü. TATA kutusunun önünde verimli transkripsiyon için gerekli olan üç dizi elemanı içerir; bir CCAAT kutusu ve iki GC kutusu (konsensus dizisi GGGCGG)
 Memeli hücrelerindeki pek çok gen • Transkripsiyon başlangıç yerine daha uzak olarak yerleşmiş (bazen 10 kb den fazla) regülatör diziler tarafından kontrol edilir. • Bunlar güçlendirici (enhancer) olarak adlandırılır • İlk olarak SV 40 promotöründe tanımlanmıştır • Transkripsiyonun verimliliği için TATA kutusu ve altı adet GC kutusuna ek olarak daha yukarıda yerleşmiş iki adet 72 bç lik tekrar gereklidir. • Etkinliği ne uzaklığı ile ne de yönü ile ilişkili değil • Promotörün hem yukarı hem de aşağı yönde yerleştiğinde uyarabilirler
Memeli hücrelerindeki pek çok gen • Transkripsiyon başlangıç yerine daha uzak olarak yerleşmiş (bazen 10 kb den fazla) regülatör diziler tarafından kontrol edilir. • Bunlar güçlendirici (enhancer) olarak adlandırılır • İlk olarak SV 40 promotöründe tanımlanmıştır • Transkripsiyonun verimliliği için TATA kutusu ve altı adet GC kutusuna ek olarak daha yukarıda yerleşmiş iki adet 72 bç lik tekrar gereklidir. • Etkinliği ne uzaklığı ile ne de yönü ile ilişkili değil • Promotörün hem yukarı hem de aşağı yönde yerleştiğinde uyarabilirler
 SV 40 güçlendiricisi: SV 40 ın erken gen ekspresyonu için kullandığı promotörü, TATA kutusu ve üç grup halinde tekrarlanan altı GC kutusu içerir. Buna ek olarak verimli transkripsiyon için yukarı yönde yerleşmiş iki adet 72 baz çiftlik tekrardan oluşmuş güçlendirici gereklidir.
SV 40 güçlendiricisi: SV 40 ın erken gen ekspresyonu için kullandığı promotörü, TATA kutusu ve üç grup halinde tekrarlanan altı GC kutusu içerir. Buna ek olarak verimli transkripsiyon için yukarı yönde yerleşmiş iki adet 72 baz çiftlik tekrardan oluşmuş güçlendirici gereklidir.
 Güçlendiriciler, • Bu diziler promotörün hem yukarı hemde aşağı yönünde yerleştiklerinde transkripsiyonu uyarabilirler. • Başlama yerinden uzakta bulunduklarında dahi iş görürler • Güçlendiriciler de promotörler gibi transkripsiyon faktörlerinin bağlanması ile iş görürler • Uzaktaki Güçlendiriciye bağlanan TF ler promotör bölgede RNA polimeraza bağlanan proteinlerle DNA ilmiklenmesi sayesinde etkileşirler.
Güçlendiriciler, • Bu diziler promotörün hem yukarı hemde aşağı yönünde yerleştiklerinde transkripsiyonu uyarabilirler. • Başlama yerinden uzakta bulunduklarında dahi iş görürler • Güçlendiriciler de promotörler gibi transkripsiyon faktörlerinin bağlanması ile iş görürler • Uzaktaki Güçlendiriciye bağlanan TF ler promotör bölgede RNA polimeraza bağlanan proteinlerle DNA ilmiklenmesi sayesinde etkileşirler.
. Güçlendirici etkisinde olmayan gen düşük seviyede transkripsiyon Yapar. Bir güçlendirici") Güçlendiricilerin çalışma şekli (A). Güçlendirici etkisinde olmayan gen düşük seviyede transkripsiyon Yapar. Bir güçlendirici (E) eklenmesi (örn. SV 40 ın 72 baz çifti tekrarları) transkripsiyonu uyarır. Güçlendirici, sadece promotörün tam yukarı yönünde yerleştiğinde (B) değil, transkripsiyon başlama yerinin birkaç kilobaz yukarısı veya aşağısına eklendiğinde de (Cve D) Aktivite gösterir.
Güçlendiricilerin çalışma şekli (A). Güçlendirici etkisinde olmayan gen düşük seviyede transkripsiyon Yapar. Bir güçlendirici (E) eklenmesi (örn. SV 40 ın 72 baz çifti tekrarları) transkripsiyonu uyarır. Güçlendirici, sadece promotörün tam yukarı yönünde yerleştiğinde (B) değil, transkripsiyon başlama yerinin birkaç kilobaz yukarısı veya aşağısına eklendiğinde de (Cve D) Aktivite gösterir.
 DNA nın ilmiklenmesi DNA nın ilmik oluşturabilmesi sayesinde uzak güçlendiricilere bağlı transkripsiyon faktörleri, RNA polimeraz II/ aracı kompleksi veya promotördeki genel transkripsiyon faktörleriyle etkileşebilirler. Bu yüzden transkripsiyon faktörlerinin DNA ya, promotörün tam yukarısında bağlanmaları ile uzak bir güçlendiriciye bağlanmaları arasında işlevsellik açısından farklılık yoktur
DNA nın ilmiklenmesi DNA nın ilmik oluşturabilmesi sayesinde uzak güçlendiricilere bağlı transkripsiyon faktörleri, RNA polimeraz II/ aracı kompleksi veya promotördeki genel transkripsiyon faktörleriyle etkileşebilirler. Bu yüzden transkripsiyon faktörlerinin DNA ya, promotörün tam yukarısında bağlanmaları ile uzak bir güçlendiriciye bağlanmaları arasında işlevsellik açısından farklılık yoktur
 • Özel Transkripsiyon düzenleyici proteinlerin güçlendiricilere bağlanması • Gelişim ve farklılaşma süreclerinde • Hormon ve GF lere yanıtlarda • Gen ifadesi kontrolünden sorumlu • B lenfositlerinde Ig genlerinin transkripsiyonunu kontrol eden güçlendiriciler diğer hücrelerde aktif değil • Farklılaşmış hücrelerde Ig genlerinin dokuya özgü ekspresyondan sorumlu
• Özel Transkripsiyon düzenleyici proteinlerin güçlendiricilere bağlanması • Gelişim ve farklılaşma süreclerinde • Hormon ve GF lere yanıtlarda • Gen ifadesi kontrolünden sorumlu • B lenfositlerinde Ig genlerinin transkripsiyonunu kontrol eden güçlendiriciler diğer hücrelerde aktif değil • Farklılaşmış hücrelerde Ig genlerinin dokuya özgü ekspresyondan sorumlu
 Regülatör diziler • Güçlendiriciler, • farklı dokuda, farklı genlerin promotör etkinliğini arttırabilir. • Genin ekspresyonunun yapılıp yapılmayacağını da belirler. • Dokuya özgü proteinlerle etkileşirler.
Regülatör diziler • Güçlendiriciler, • farklı dokuda, farklı genlerin promotör etkinliğini arttırabilir. • Genin ekspresyonunun yapılıp yapılmayacağını da belirler. • Dokuya özgü proteinlerle etkileşirler.
 • Güçlendiriciler • Farklı düzenleyici proteinlerin bağlandığı çok sayıda dizi elemanı içerir. • Bu proteinler beraberce gen ekspresyonu düzenlenmesini sağlar • Ig ağır zincir güçlendiricisi 200 bç dir ve 9 işlevsel dizi elemanı içerir
• Güçlendiriciler • Farklı düzenleyici proteinlerin bağlandığı çok sayıda dizi elemanı içerir. • Bu proteinler beraberce gen ekspresyonu düzenlenmesini sağlar • Ig ağır zincir güçlendiricisi 200 bç dir ve 9 işlevsel dizi elemanı içerir
 İmmünoglobulin güçlendiricisi • En çok çalışılan memeli güçlendiricisi • B lenfositlerde immünoglobulin genlerinin transkripsiyonunu kontrol eden güçlendiricidir • Bu güçlendirici lenfositlerden başka hücrelerde aktif değil • Dokuya özgü ekspresyondan sorumlu • Güçlendiriciler genellikle Farklı düzenleyici proteinlerin bağlandığı çok sayıda dizi elemanı içerirler. • Bunlardan herhangi birindeki mutasyon güçlendirici etkinliğini düşürür. Fakat durdurmaz.
İmmünoglobulin güçlendiricisi • En çok çalışılan memeli güçlendiricisi • B lenfositlerde immünoglobulin genlerinin transkripsiyonunu kontrol eden güçlendiricidir • Bu güçlendirici lenfositlerden başka hücrelerde aktif değil • Dokuya özgü ekspresyondan sorumlu • Güçlendiriciler genellikle Farklı düzenleyici proteinlerin bağlandığı çok sayıda dizi elemanı içerirler. • Bunlardan herhangi birindeki mutasyon güçlendirici etkinliğini düşürür. Fakat durdurmaz.
 İmmünoglobulin güçlendiricisi İmmünoglobulin ağır zincir güçlendiricisi, yaklaşık 200 baz uzunluktadır ve B lenfositlerinde, birlikte transkripsiyonu uyaran 9 işlevsel dizi elemanı içerir
İmmünoglobulin güçlendiricisi İmmünoglobulin ağır zincir güçlendiricisi, yaklaşık 200 baz uzunluktadır ve B lenfositlerinde, birlikte transkripsiyonu uyaran 9 işlevsel dizi elemanı içerir
 Regülatör diziler ▫ ▫ transkripsiyon başlangıç yerine uzaktır. Transkripsiyonu başlatmaz Transkripsiyon miktarını kontrol eder. Etkileri hücre tipine bağlıdır ▫ Promotörlerin çoğunun etkinliği, ayrı bir düzenleyici dizi tarafından düzenlenir
Regülatör diziler ▫ ▫ transkripsiyon başlangıç yerine uzaktır. Transkripsiyonu başlatmaz Transkripsiyon miktarını kontrol eder. Etkileri hücre tipine bağlıdır ▫ Promotörlerin çoğunun etkinliği, ayrı bir düzenleyici dizi tarafından düzenlenir
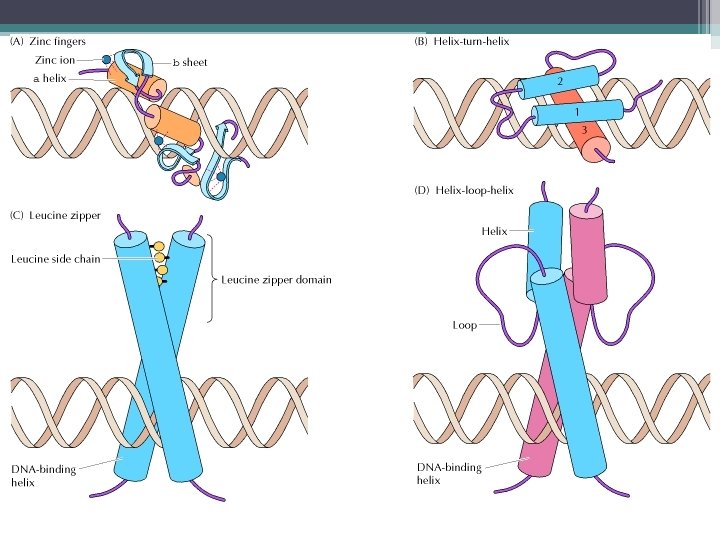
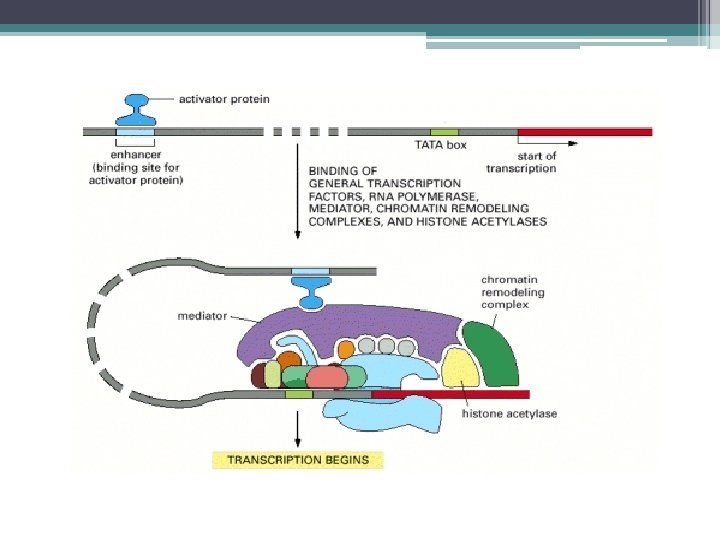
 Transkripsiyon kontrol eden proteinler • Regülatör dizilere bağlanan ve RNA Polimeraz II etkinliğini düzenleyen proteinler tarafından yapılır. • Bunlardan • transkripsiyon aktivatörleri : ▫ Transkripsiyon faktörleri ile etkileşerek (TFIIB ve TFIID) RNA polimerazın aktifleşmesini sağlayarak ▫ Kromatin yapısını değiştirerek ▫ Transkripsiyonu uyaran ko-aktivatörlerle etkileşerek etkilerini gösterirler. ▫ transkripsiyon aktivatörleri ▫ Promotöre veya güçlendirici dizilere bağlanır
Transkripsiyon kontrol eden proteinler • Regülatör dizilere bağlanan ve RNA Polimeraz II etkinliğini düzenleyen proteinler tarafından yapılır. • Bunlardan • transkripsiyon aktivatörleri : ▫ Transkripsiyon faktörleri ile etkileşerek (TFIIB ve TFIID) RNA polimerazın aktifleşmesini sağlayarak ▫ Kromatin yapısını değiştirerek ▫ Transkripsiyonu uyaran ko-aktivatörlerle etkileşerek etkilerini gösterirler. ▫ transkripsiyon aktivatörleri ▫ Promotöre veya güçlendirici dizilere bağlanır
![Transkripsiyon düzenleyici proteinler • Promotör dizilerden; • CAAT ve GGGCGG [GC Kutusu] dizilerine bağlanan](https://present5.com/presentation/2bfdcbc56d3720c0bfec630349eb995d/image-111.jpg "Transkripsiyon düzenleyici proteinler • Promotör dizilerden; • CAAT ve GGGCGG [GC Kutusu] dizilerine bağlanan") Transkripsiyon düzenleyici proteinler • Promotör dizilerden; • CAAT ve GGGCGG [GC Kutusu] dizilerine bağlanan ve • transkripsiyonu uyaran proteinler vardır. Örn. • SP-1 faktörü (spesifisite-1 faktörü) GC kutusuna bağlanır. GC yoksa uyaramaz. • İlk bulunan transkripsiyon aktivatörüdür • SP-1, iki altbirimden oluşur. • Biri DNA’ ya, • diğeri transkripsiyon faktörlerine bağlanır. .
Transkripsiyon düzenleyici proteinler • Promotör dizilerden; • CAAT ve GGGCGG [GC Kutusu] dizilerine bağlanan ve • transkripsiyonu uyaran proteinler vardır. Örn. • SP-1 faktörü (spesifisite-1 faktörü) GC kutusuna bağlanır. GC yoksa uyaramaz. • İlk bulunan transkripsiyon aktivatörüdür • SP-1, iki altbirimden oluşur. • Biri DNA’ ya, • diğeri transkripsiyon faktörlerine bağlanır. .
 Transkripsiyon aktivatörlerinin belirlenmesi için denatüre etmeyen jelde elektroforez yöntemi kullanılması
Transkripsiyon aktivatörlerinin belirlenmesi için denatüre etmeyen jelde elektroforez yöntemi kullanılması
 Transkripsiyon represörleri • Regülatör dizilere bağlanarak, transkripsiyonu baskılar. Promotöre • transkripsiyon faktörlerinin ya da • RNA polimerazın bağlanmasını engelleyerek etkilerini gerçekleştirirler • Bazı represörler regülatör diziye bağlanmak için aktivatörler ile yarışır.
Transkripsiyon represörleri • Regülatör dizilere bağlanarak, transkripsiyonu baskılar. Promotöre • transkripsiyon faktörlerinin ya da • RNA polimerazın bağlanmasını engelleyerek etkilerini gerçekleştirirler • Bazı represörler regülatör diziye bağlanmak için aktivatörler ile yarışır.
Bazı represörler aktivatörlerin regülatör dizilere bağlanmasını engeller B)Diğerleri aracı proteinlerle") Ökaryot represörlerinin işleyiş şekli. A)Bazı represörler aktivatörlerin regülatör dizilere bağlanmasını engeller B)Diğerleri aracı proteinlerle etkileşerek (veya genel transkripsiyon faktörleri ile ve kromatin yapılanmasını değiştiren ko -represörlerle etkileşerek transkripsiyonu engeller
Ökaryot represörlerinin işleyiş şekli. A)Bazı represörler aktivatörlerin regülatör dizilere bağlanmasını engeller B)Diğerleri aracı proteinlerle etkileşerek (veya genel transkripsiyon faktörleri ile ve kromatin yapılanmasını değiştiren ko -represörlerle etkileşerek transkripsiyonu engeller
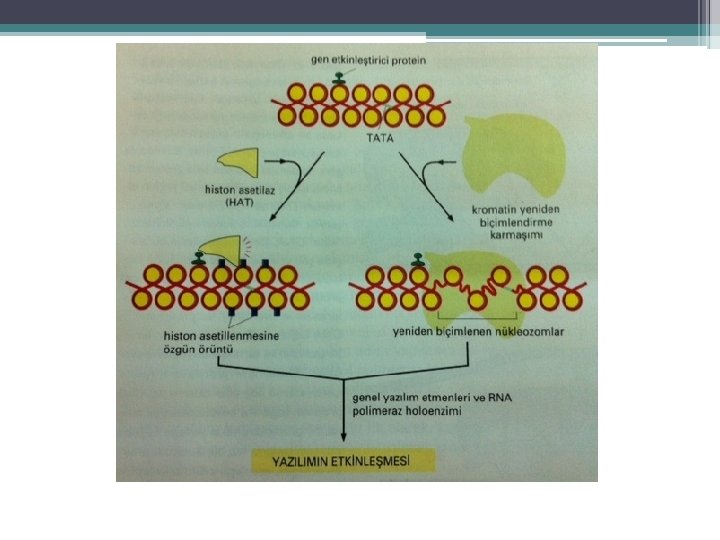
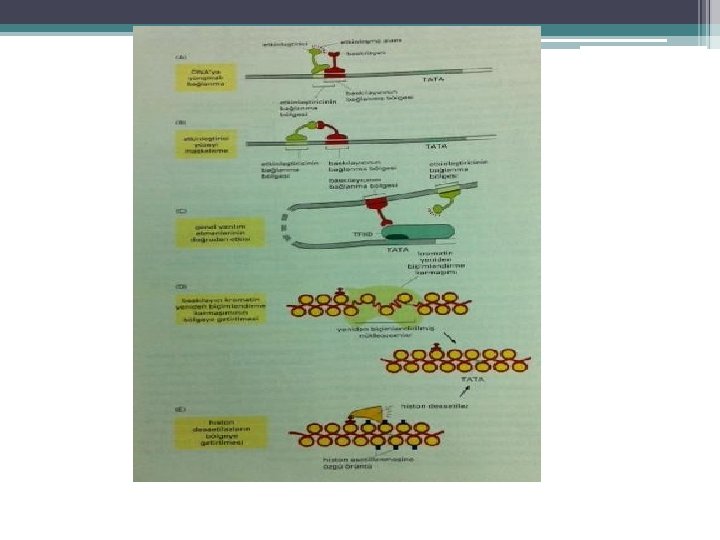
 KROMATİN YAPISI İLE TRANSKRİPSİYON İLİŞKİSİ Aktivatör ve represörler Hem Transkripsiyon sisteminin diğer bileşenleri ile etkileşerek Hem de kromatin yapısındaki değişiklikler ile düzenler. Ökaryotlarda; • DNA histonlarla sıkıca bağlı • Nükleozom • H 2 A, H 2 B, H 3, H 4 histonların her birinden (2 şer adet) • 146 bç DNA • Aktif genler 30 nm. lik İplik yapısında • Az yoğun kromatin
KROMATİN YAPISI İLE TRANSKRİPSİYON İLİŞKİSİ Aktivatör ve represörler Hem Transkripsiyon sisteminin diğer bileşenleri ile etkileşerek Hem de kromatin yapısındaki değişiklikler ile düzenler. Ökaryotlarda; • DNA histonlarla sıkıca bağlı • Nükleozom • H 2 A, H 2 B, H 3, H 4 histonların her birinden (2 şer adet) • 146 bç DNA • Aktif genler 30 nm. lik İplik yapısında • Az yoğun kromatin
 Az yoğun kromatin de görülen özellikler • Histon modifikasyonu • Nükleozom yeniden düzenlenmesi • HMGN proteinlerinin (non-histon protein ) aktif transkripsiyonu yapılan genlerin nükleozomlarına bağlanması HMGN proteinleri nukleozom üzerinde ; • H 1’in bağlanma yerine bağlanıyor. • H 1’in nükleozom ile etkileşimini bozarak yoğun olmayan kromatin yapısının devamlılığını sağlar. • Transkripsiyonu uyarır.
Az yoğun kromatin de görülen özellikler • Histon modifikasyonu • Nükleozom yeniden düzenlenmesi • HMGN proteinlerinin (non-histon protein ) aktif transkripsiyonu yapılan genlerin nükleozomlarına bağlanması HMGN proteinleri nukleozom üzerinde ; • H 1’in bağlanma yerine bağlanıyor. • H 1’in nükleozom ile etkileşimini bozarak yoğun olmayan kromatin yapısının devamlılığını sağlar. • Transkripsiyonu uyarır.
") Histon modifikasyonları HİSTON ASETİLLENMESİ • Aktif kromatinde görülür. • Kor Histonların N ucu (kuyruk) nükleozomdan dışa uzanır, lizinden zengin LİZİNLERİN ASETİLLENMESİ ile modifiye edilir • Histonların pozitif yükünü azaltır • onların DNAya bağlanma ve diğer proteinlerle bağlanmasını zayıflatabilir. • Nükleozomal DNA ya transkripsiyon faktörlerinin bağlanmasını kolaylaştırır. • Proteinlerin kromatine ulaşılabilirliğini de arttırır
Histon modifikasyonları HİSTON ASETİLLENMESİ • Aktif kromatinde görülür. • Kor Histonların N ucu (kuyruk) nükleozomdan dışa uzanır, lizinden zengin LİZİNLERİN ASETİLLENMESİ ile modifiye edilir • Histonların pozitif yükünü azaltır • onların DNAya bağlanma ve diğer proteinlerle bağlanmasını zayıflatabilir. • Nükleozomal DNA ya transkripsiyon faktörlerinin bağlanmasını kolaylaştırır. • Proteinlerin kromatine ulaşılabilirliğini de arttırır
 REPRESÖRLERİN Histon deasetilaz (HDAC) ile birleştikleri 1996 da gösterildi. Histon asetilasyonu") AKTİVATÖRLERİN Histon asetiltransferaz(HAT) REPRESÖRLERİN Histon deasetilaz (HDAC) ile birleştikleri 1996 da gösterildi. Histon asetilasyonu ile transkripsiyon ilişkisi ortaya kondu. Tetrahymena nın HAT geni klonlandı. Transkripsiyon kofaktörü (mayanın) Gcn 5 p ile dizisi çok benzer çıktı. Gcn 5 p Transkripsiyon aktivatörleri ile birleşerek transkripsiyonu uyarır. Gcn 5 p HAT etkinliği gösterir HAT ın birçok memeli transkripsiyon faktörü ile ve TFIID ile benzer olduğu gösterildi. • Memeli ve mayalarda Represörler HDAC işlevi görür.
AKTİVATÖRLERİN Histon asetiltransferaz(HAT) REPRESÖRLERİN Histon deasetilaz (HDAC) ile birleştikleri 1996 da gösterildi. Histon asetilasyonu ile transkripsiyon ilişkisi ortaya kondu. Tetrahymena nın HAT geni klonlandı. Transkripsiyon kofaktörü (mayanın) Gcn 5 p ile dizisi çok benzer çıktı. Gcn 5 p Transkripsiyon aktivatörleri ile birleşerek transkripsiyonu uyarır. Gcn 5 p HAT etkinliği gösterir HAT ın birçok memeli transkripsiyon faktörü ile ve TFIID ile benzer olduğu gösterildi. • Memeli ve mayalarda Represörler HDAC işlevi görür.
 • • Ayrıca Histonlar Serin kalıntılarında fosfatlanarak Lizin ve argininden metillenerek Lizinlerin ubikutinlenmesi ile modifiye edilirler (histon kuyruklarından) Bunların hepsi birlikte Bazı düzenleyici proteinleri kromatine getiren ve gen ifadesini düzenleyen bir histon kodu oluşturur
• • Ayrıca Histonlar Serin kalıntılarında fosfatlanarak Lizin ve argininden metillenerek Lizinlerin ubikutinlenmesi ile modifiye edilirler (histon kuyruklarından) Bunların hepsi birlikte Bazı düzenleyici proteinleri kromatine getiren ve gen ifadesini düzenleyen bir histon kodu oluşturur
 • • Aktif kromatin H 3 ün lizin 4 de metillenme Serin 10 da fosfatlanma Lizin 9 ve lizin 14 asetillenmesini kapsayan özgün modifikasyonlar içerir. • İnaktif kromatin ise • Lizin 9 metillenmesi ile ilişkili Kromatin yoğunlaşması ile ilgili • histonları değiştiren enzimler ile kromatin yapısının düzenlenmesi
• • Aktif kromatin H 3 ün lizin 4 de metillenme Serin 10 da fosfatlanma Lizin 9 ve lizin 14 asetillenmesini kapsayan özgün modifikasyonlar içerir. • İnaktif kromatin ise • Lizin 9 metillenmesi ile ilişkili Kromatin yoğunlaşması ile ilgili • histonları değiştiren enzimler ile kromatin yapısının düzenlenmesi
 Nukleozom yeniden biçimlendirme faktörleri • Nukleozomların yapısını • Histonları modifiye etmeden değiştiren protein kompleksleridir • Etki mekanizmalarından biri • Histon oktamerlerinin DNA üzerinde kaymasını katalizleyerek nukleozomların yerleşimini değiştirmek • Transkripsiyon faktörlerinin spesifik DNA dizilerine ulaşmasını sağlamak
Nukleozom yeniden biçimlendirme faktörleri • Nukleozomların yapısını • Histonları modifiye etmeden değiştiren protein kompleksleridir • Etki mekanizmalarından biri • Histon oktamerlerinin DNA üzerinde kaymasını katalizleyerek nukleozomların yerleşimini değiştirmek • Transkripsiyon faktörlerinin spesifik DNA dizilerine ulaşmasını sağlamak
 Nukleozom yeniden biçimlendirme faktörleri • Bu faktörler nukleozom şeklinin değişimine neden olarak da transkripsiyon düzenleyen proteinlerin spesifik DNA dizileri ile etkileşmesini sağlayabilir. • Histon modifiye eden enzimler ve • Nukleozom yeniden biçimlendirme faktörlerinin • Transkripsiyon aktivatörleri ile birlikte bağlanması • Güçlendirici ve promotör bölgelerde kromatin yapısını değiştirerek transkripsiyonun başlamasını sağlar
Nukleozom yeniden biçimlendirme faktörleri • Bu faktörler nukleozom şeklinin değişimine neden olarak da transkripsiyon düzenleyen proteinlerin spesifik DNA dizileri ile etkileşmesini sağlayabilir. • Histon modifiye eden enzimler ve • Nukleozom yeniden biçimlendirme faktörlerinin • Transkripsiyon aktivatörleri ile birlikte bağlanması • Güçlendirici ve promotör bölgelerde kromatin yapısını değiştirerek transkripsiyonun başlamasını sağlar
 DNA metillenmesi • Omurgalılarda transkripsiyon kontrolünde kromatin yapısı ile ilişkili genel bir mekanizmadır. • DNA daki Sitozin birimleri 5 -pozisyonundaki karbona metil grupları takılarak modifiye edilir. • Cp. G adacıklarında metillenme bunları içeren genlerin transkripsiyon etkinliğinin azaltılması ile ilişkilidir • Transkripsiyon aktivatörlerinin bağlanmasını engelleyerek veya • Metillenmiş DNA ya bağlanan represörlerin katılımını sağlayarak • Gelişim sürecinde gen inaktivasyonunun sürekliliğinde ve korunmasında katkı sağlar
DNA metillenmesi • Omurgalılarda transkripsiyon kontrolünde kromatin yapısı ile ilişkili genel bir mekanizmadır. • DNA daki Sitozin birimleri 5 -pozisyonundaki karbona metil grupları takılarak modifiye edilir. • Cp. G adacıklarında metillenme bunları içeren genlerin transkripsiyon etkinliğinin azaltılması ile ilişkilidir • Transkripsiyon aktivatörlerinin bağlanmasını engelleyerek veya • Metillenmiş DNA ya bağlanan represörlerin katılımını sağlayarak • Gelişim sürecinde gen inaktivasyonunun sürekliliğinde ve korunmasında katkı sağlar
 GEN İFADESİNİN KONTROLÜ Gen ifadesini gerçekleştiren her basamakta düzenlenebilir. • En çok kullanılan transkripsiyon un başlangıcının kontrolü • Daha sonra devreye giren denetimler ürün miktarını değiştirebilir • Posttranskripsiyonel kontroller birçok gen için çok önemlidir
GEN İFADESİNİN KONTROLÜ Gen ifadesini gerçekleştiren her basamakta düzenlenebilir. • En çok kullanılan transkripsiyon un başlangıcının kontrolü • Daha sonra devreye giren denetimler ürün miktarını değiştirebilir • Posttranskripsiyonel kontroller birçok gen için çok önemlidir
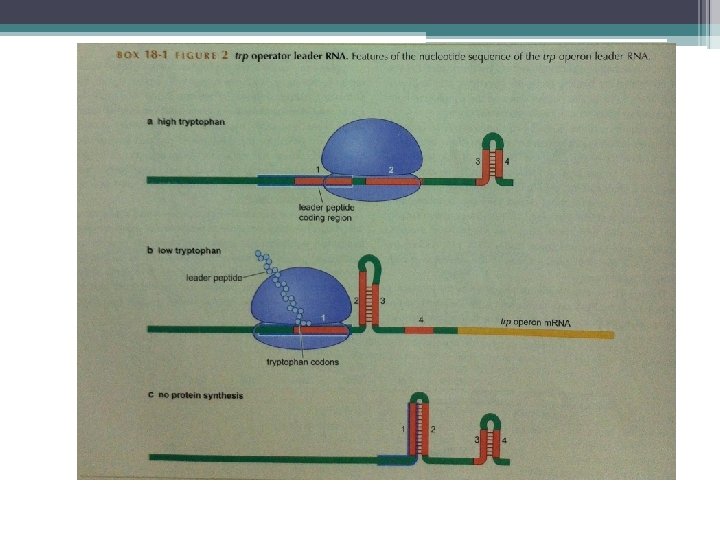
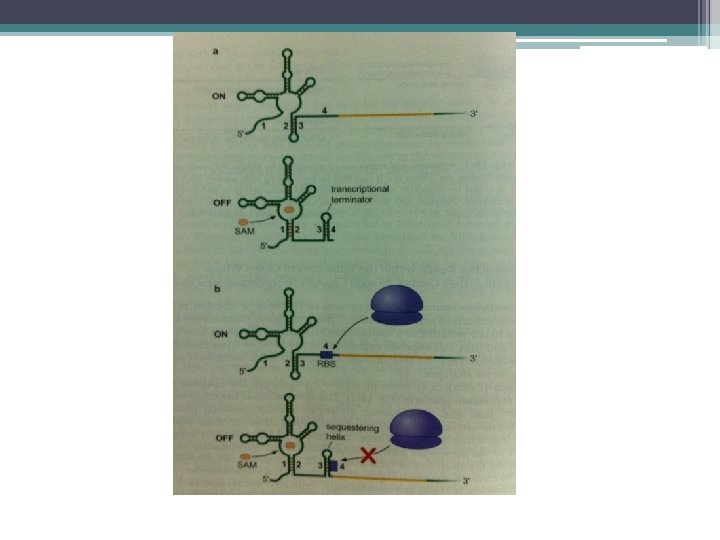
 Transkripsiyonun zayıflatılması • Bakterilerde bazı genlerin ifadesi transkripsiyonun erken sonlandırılması ile önlenir. • Sentezlenen RNA zinciri RNA polimerazın trankripsiyonu sonlandırmasına neden olan bir yapı kazanır. • Ürüne ihtiyaç olduğunda düzenleyici proteinler, oluşan RNA zincirine bağlanır ve • sonlandırmaya neden olan durum ortadan kaldırılır. • RNA’nın transkripsiyonu gerçekleşir.
Transkripsiyonun zayıflatılması • Bakterilerde bazı genlerin ifadesi transkripsiyonun erken sonlandırılması ile önlenir. • Sentezlenen RNA zinciri RNA polimerazın trankripsiyonu sonlandırmasına neden olan bir yapı kazanır. • Ürüne ihtiyaç olduğunda düzenleyici proteinler, oluşan RNA zincirine bağlanır ve • sonlandırmaya neden olan durum ortadan kaldırılır. • RNA’nın transkripsiyonu gerçekleşir.
 Ökaryotlarda transkripsiyonun zayıflatılması farklı mekanizmalarla olur. HIV virüsü iyi incelenmiş bir örnek • Virüs konak genomuna girdikten sonra • hücrenin RNA pol II si viral DNA’nın transkripsiyonunu sağlar • Konak polimeraz birkaç yüz nükleotid sentezledikten sonra transkripsiyonu sonlandırır. • Viral DNA’nın tümünün trankripsiyonu gerçekleşmez. • Uygun şartlar olduğunda virüs tarafından kodlanan Tat proteini erken sonlanmayı önler. • Tat proteini uzayan RNA da özgün bir sap ilmek yapısına bağlanır. (bu yapıya Tar denir. )
Ökaryotlarda transkripsiyonun zayıflatılması farklı mekanizmalarla olur. HIV virüsü iyi incelenmiş bir örnek • Virüs konak genomuna girdikten sonra • hücrenin RNA pol II si viral DNA’nın transkripsiyonunu sağlar • Konak polimeraz birkaç yüz nükleotid sentezledikten sonra transkripsiyonu sonlandırır. • Viral DNA’nın tümünün trankripsiyonu gerçekleşmez. • Uygun şartlar olduğunda virüs tarafından kodlanan Tat proteini erken sonlanmayı önler. • Tat proteini uzayan RNA da özgün bir sap ilmek yapısına bağlanır. (bu yapıya Tar denir. )
 • Tat • RNA polimerazın transkripsiyona devamını sağlayan proteinleri buraya toplar • (bunların bir kısmı erken sonlanmayı önleyen proteinlerdir. ) • Böylece HIV tek bir viral proteinin etkisiyle ve • normal hücresel mekanizmalardan faydalanarak kendi genomunun transkripsiyonunu sağlar.
• Tat • RNA polimerazın transkripsiyona devamını sağlayan proteinleri buraya toplar • (bunların bir kısmı erken sonlanmayı önleyen proteinlerdir. ) • Böylece HIV tek bir viral proteinin etkisiyle ve • normal hücresel mekanizmalardan faydalanarak kendi genomunun transkripsiyonunu sağlar.
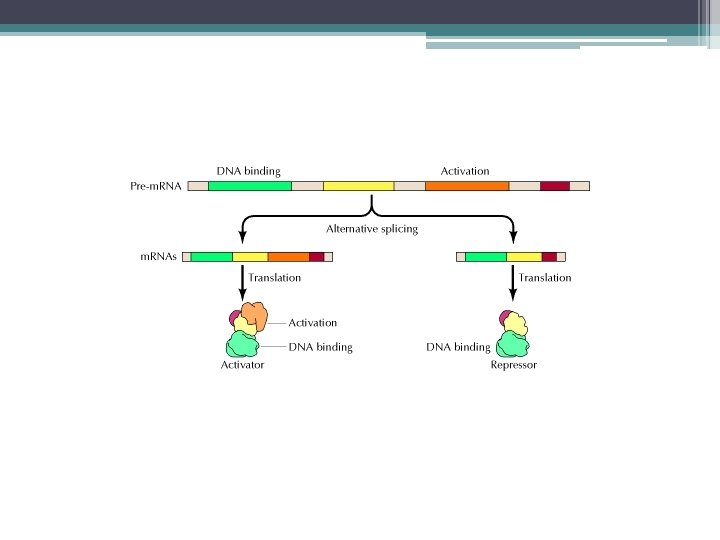
 Alternatif kırpılma ile gen ifadesinin kontrolu
Alternatif kırpılma ile gen ifadesinin kontrolu
 Drozofila’da cinsiyetin belirlenmesi bir dizi RNA kırpılma olayına bağlıdır. • RNA kırpılmasının düzenlenmesi en iyi bilinen örnek Drozofilanın erkek veya dişi olacağının X kromozomu/ otozom oranı ile belirlenmesi • 2 X + 2 otozom = dişi • X + 2 otozom = erkek (0, 5)
Drozofila’da cinsiyetin belirlenmesi bir dizi RNA kırpılma olayına bağlıdır. • RNA kırpılmasının düzenlenmesi en iyi bilinen örnek Drozofilanın erkek veya dişi olacağının X kromozomu/ otozom oranı ile belirlenmesi • 2 X + 2 otozom = dişi • X + 2 otozom = erkek (0, 5)
 RNA lar • Jacob ve Monod, 50 yıl önce gen regülasyonu çalışmaları") Düzenleyici (Regülatör) RNA lar • Jacob ve Monod, 50 yıl önce gen regülasyonu çalışmaları ile represörleri buldular. • Ancak; bunların protein mi RNA mı olduğunu söyleyemediler. • Çoğu durumda düzenleyiciler DNA ya bağlanan proteinler • RNA molekülleri olabileceğine işaret ettiler • Protein düzenleyiciler bulundukça bu düşünce unutuldu
Düzenleyici (Regülatör) RNA lar • Jacob ve Monod, 50 yıl önce gen regülasyonu çalışmaları ile represörleri buldular. • Ancak; bunların protein mi RNA mı olduğunu söyleyemediler. • Çoğu durumda düzenleyiciler DNA ya bağlanan proteinler • RNA molekülleri olabileceğine işaret ettiler • Protein düzenleyiciler bulundukça bu düşünce unutuldu
 • Son yıllarda • RNA düzenleyicilerin araştırılmasında patlama yaşandı (Transkripsiyon seviyesinde ve translasyon seviyesinde iş görüyordu) • Bu yeni alan 2 kaynaktan çıktı • 1 - Mikro. RNA ların keşfi (1990 ların başı) • 2 -RNA interferans olgusunun keşfi ( geç 1990 lar)
• Son yıllarda • RNA düzenleyicilerin araştırılmasında patlama yaşandı (Transkripsiyon seviyesinde ve translasyon seviyesinde iş görüyordu) • Bu yeni alan 2 kaynaktan çıktı • 1 - Mikro. RNA ların keşfi (1990 ların başı) • 2 -RNA interferans olgusunun keşfi ( geç 1990 lar)
 Küçük RNA lar • Küçük RNA lar bakterilerde tanımlandı • Bazısı plazmid replikasyonuna • Diğerleri gen ifadesinin düzenlenmesine katılıyordu
Küçük RNA lar • Küçük RNA lar bakterilerde tanımlandı • Bazısı plazmid replikasyonuna • Diğerleri gen ifadesinin düzenlenmesine katılıyordu

 • yaklaşık 21 -23 nükleotid uzunluğunda kodlayıcı olmayan RNA lardandır") mikro. RNA (mi. RNA) • yaklaşık 21 -23 nükleotid uzunluğunda kodlayıcı olmayan RNA lardandır • Tek iplikli RNA molekülleridir • gen ifadesinin düzenlenmesinde rol oynar. • mi. RNA'lar, . Pri-mi. RNA olarak adlandırılan primer transkriptler işlenerek • önce pre-mi. RNA adlı kısa sap- ilmek yapılarına, • sonra da fonksiyonel mi. RNA'ya dönüşürler. • Olgun mi. RNA molekülleri bir veya daha çok (m. RNA) ile kısmen tamamlayıcıdır ve • başlıca işlevleri gen ifadesini azaltmaktır • 1993'te Lee ve çalışma arkadaşları tarafından keşfedilmişlerdir mikro. RNA terimi ilk 2001'de kullanıma girimiştir
mikro. RNA (mi. RNA) • yaklaşık 21 -23 nükleotid uzunluğunda kodlayıcı olmayan RNA lardandır • Tek iplikli RNA molekülleridir • gen ifadesinin düzenlenmesinde rol oynar. • mi. RNA'lar, . Pri-mi. RNA olarak adlandırılan primer transkriptler işlenerek • önce pre-mi. RNA adlı kısa sap- ilmek yapılarına, • sonra da fonksiyonel mi. RNA'ya dönüşürler. • Olgun mi. RNA molekülleri bir veya daha çok (m. RNA) ile kısmen tamamlayıcıdır ve • başlıca işlevleri gen ifadesini azaltmaktır • 1993'te Lee ve çalışma arkadaşları tarafından keşfedilmişlerdir mikro. RNA terimi ilk 2001'de kullanıma girimiştir
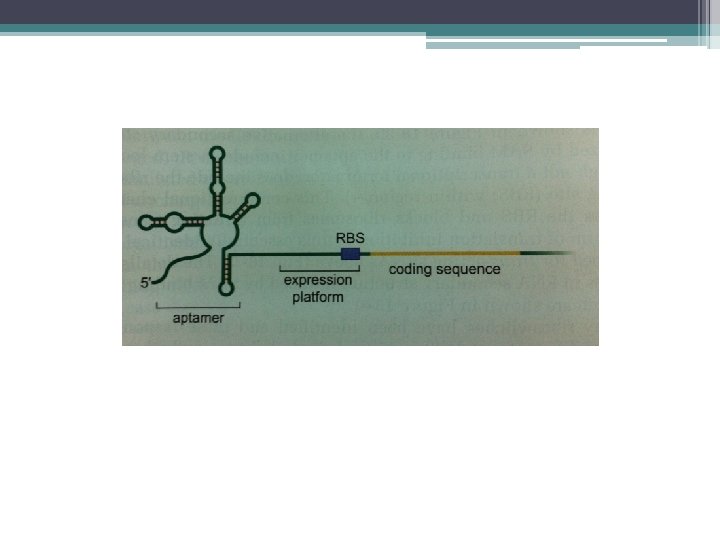
 RNA interferens • RNA interferens, ökaryotlarda temel bir düzenleyici mekanizmadır. • Bazı ökaryotik, genlerin transkriptlerinde düzenleyici RNA elemanları vardır. • Bunlar, regülatör proteinlere bağlanarak (ör. HIVTAT protein) iş görür. • Bugün RNA ların gen regülasyonunda çok daha yaygın bir rol oynadığı açığa çıkmıştır • Çeşitli Çok kısa RNA lar, kendileriyle homoloji bölgeleri içeren bazı genlerin ekspresyonunu baskılar veya susturur. Buna RNA interferans adı verilir
RNA interferens • RNA interferens, ökaryotlarda temel bir düzenleyici mekanizmadır. • Bazı ökaryotik, genlerin transkriptlerinde düzenleyici RNA elemanları vardır. • Bunlar, regülatör proteinlere bağlanarak (ör. HIVTAT protein) iş görür. • Bugün RNA ların gen regülasyonunda çok daha yaygın bir rol oynadığı açığa çıkmıştır • Çeşitli Çok kısa RNA lar, kendileriyle homoloji bölgeleri içeren bazı genlerin ekspresyonunu baskılar veya susturur. Buna RNA interferans adı verilir
 • Bu susturmaya RNA interferens (RNAi) denir. • Çeşitli şekillerde görülür;") RNA interferens (RNAi) • Bu susturmaya RNA interferens (RNAi) denir. • Çeşitli şekillerde görülür; ▫ m. RNA translasyonunu inhibe ederek ▫ m. RNA nın yıkılmasını sağlayarak ▫ m. RNA nın ekspresyonunu yöneten promotörün transkripsiyonel susturulması ile
RNA interferens (RNAi) • Bu susturmaya RNA interferens (RNAi) denir. • Çeşitli şekillerde görülür; ▫ m. RNA translasyonunu inhibe ederek ▫ m. RNA nın yıkılmasını sağlayarak ▫ m. RNA nın ekspresyonunu yöneten promotörün transkripsiyonel susturulması ile
 Kısa RNA lar • Bu kısa RNA lar, çeşitli daha uzun çift iplikli RNA lardan özel enzimler ile oluşturulur.
Kısa RNA lar • Bu kısa RNA lar, çeşitli daha uzun çift iplikli RNA lardan özel enzimler ile oluşturulur.
 • Bu RNA ların işlevlerinin; ▫ Gelişim sırasındaki regülasyondan ▫ Organizmayı viral enfeksiyonlara karşı koruyan mekanizmalara ▫ kadar geniş bir alanda olduğu kesindir. ▫ Ayrıca RNAi; güçlü bir deneysel yöntem olarak özel bir genin ekspresyonunun kapatılması (durdurulması) için kolay bir yol oluşturmaktadır.
• Bu RNA ların işlevlerinin; ▫ Gelişim sırasındaki regülasyondan ▫ Organizmayı viral enfeksiyonlara karşı koruyan mekanizmalara ▫ kadar geniş bir alanda olduğu kesindir. ▫ Ayrıca RNAi; güçlü bir deneysel yöntem olarak özel bir genin ekspresyonunun kapatılması (durdurulması) için kolay bir yol oluşturmaktadır.
 •") Küçük RNA lar Orijinlerine göre farklı isim alırlar Mikro RNA lar, (mi. RNA) • hücrede genlerde şifrelenen prekürsör RNA lardan oluşur ve özgün regülatör işlevleri vardır • • Diğer bir düzenleyici RNA grubu. Yapay olarak yapılan ya da in vivo olarak ds. RNA öncüllerinden yapılanlar, küçük interfering RNA adını alır (si. RNA)
Küçük RNA lar Orijinlerine göre farklı isim alırlar Mikro RNA lar, (mi. RNA) • hücrede genlerde şifrelenen prekürsör RNA lardan oluşur ve özgün regülatör işlevleri vardır • • Diğer bir düzenleyici RNA grubu. Yapay olarak yapılan ya da in vivo olarak ds. RNA öncüllerinden yapılanlar, küçük interfering RNA adını alır (si. RNA)
 Olgun mi. RNA’lar o Tek iplikli ve 21 nukleotidlik RNA molekülleridir. o mi. RNA genlerinin genellikle RNA polimeraz II ile transkripsiyonu yapılır. o Primer transkript (yüzlerce nt uzunlukta olabilir)primi. RNA o pri-mi. RNA Nukleusta Drosha enzimi (RNAz III) ile işlenir pre-mi. RNA oluşur. o (pre-mi. RNA)lar 70 -100 nukleotidlik saç tokası şeklindeki öncül mi. RNAlardır o Sitoplazmaya taşınır (exportin-5 ile)
Olgun mi. RNA’lar o Tek iplikli ve 21 nukleotidlik RNA molekülleridir. o mi. RNA genlerinin genellikle RNA polimeraz II ile transkripsiyonu yapılır. o Primer transkript (yüzlerce nt uzunlukta olabilir)primi. RNA o pri-mi. RNA Nukleusta Drosha enzimi (RNAz III) ile işlenir pre-mi. RNA oluşur. o (pre-mi. RNA)lar 70 -100 nukleotidlik saç tokası şeklindeki öncül mi. RNAlardır o Sitoplazmaya taşınır (exportin-5 ile)
 ile tekrar işlenir. • si-RNA") mi. RNA oluşumu o Sitoplazmada Dicer enzimi (RNAz III) ile tekrar işlenir. • si-RNA benzeri bir dupleks oluşur. • 5’ ucundan zayıf baz eşleşmesi ile RISC = (RNA ile indüklenen susturma kompleksi) içine girer o mi. RNA nın hedefine bağlanması RISC içinde olur.
mi. RNA oluşumu o Sitoplazmada Dicer enzimi (RNAz III) ile tekrar işlenir. • si-RNA benzeri bir dupleks oluşur. • 5’ ucundan zayıf baz eşleşmesi ile RISC = (RNA ile indüklenen susturma kompleksi) içine girer o mi. RNA nın hedefine bağlanması RISC içinde olur.
 o mi. RNA’nın 5’ ucu ile hedef m. RNA’nın 3’ UTR bölgesi arasındaki komplementerlik bağlanma için esastır. o Susturulma için Argonaute ailesinden (AGO) proteinler gerekli.
o mi. RNA’nın 5’ ucu ile hedef m. RNA’nın 3’ UTR bölgesi arasındaki komplementerlik bağlanma için esastır. o Susturulma için Argonaute ailesinden (AGO) proteinler gerekli.
 • RISC mi. RNA veya si. RNA ya ek olarak Argonaute ailesinden proteinler de içerir Argonaute katalitik alt birimdir m. RNA nın ilk kesilmesini gerçekleştirir (Slicer da denir) • Eğer dizi çok iyi komplementer ise hedef yıkılır • Eğer eşleşme iyi değil ise ( birkaç bç eşleşmemişse) • translasyon inhibisyonu olur • RNAİ susturması çok verimlidir • Çok az miktarda ds RNA hedek genlerin ekspresyonunun nerdeyse tamamen durdurulmasını indüklemek için yeterlidir
• RISC mi. RNA veya si. RNA ya ek olarak Argonaute ailesinden proteinler de içerir Argonaute katalitik alt birimdir m. RNA nın ilk kesilmesini gerçekleştirir (Slicer da denir) • Eğer dizi çok iyi komplementer ise hedef yıkılır • Eğer eşleşme iyi değil ise ( birkaç bç eşleşmemişse) • translasyon inhibisyonu olur • RNAİ susturması çok verimlidir • Çok az miktarda ds RNA hedek genlerin ekspresyonunun nerdeyse tamamen durdurulmasını indüklemek için yeterlidir
 prekursör mikro. RNA (pre-mi. RNA)'dan meydana gelir, o da") Micro. RNA (mi. RNA) prekursör mikro. RNA (pre-mi. RNA)'dan meydana gelir, o da bir mikro. RNA primer transkriptinden (pri-mi. RNA'dan) meydana gelir Micro. RNA (mi. RNA) prekursör mikro. RNA (pre-mi. RNA)'dan meydana gelir, o da bir mikro. RNA primer transkriptinden (primi. RNA'dan) meydana gelir.
Micro. RNA (mi. RNA) prekursör mikro. RNA (pre-mi. RNA)'dan meydana gelir, o da bir mikro. RNA primer transkriptinden (pri-mi. RNA'dan) meydana gelir Micro. RNA (mi. RNA) prekursör mikro. RNA (pre-mi. RNA)'dan meydana gelir, o da bir mikro. RNA primer transkriptinden (primi. RNA'dan) meydana gelir.
 is") . g i f RNAi silencing is initiated when double-stranded RNA (ds. RNA) is
. g i f RNAi silencing is initiated when double-stranded RNA (ds. RNA) is
 - mi. RNAs encode a novel class of small, nc. RNAs that regulate fig 1_blurb‑ 3. jpg
- mi. RNAs encode a novel class of small, nc. RNAs that regulate fig 1_blurb‑ 3. jpg
 vitro studies of RNA-induced gene silencing, or RNA This model is based on the results of in vitro studies of RNA-induced gene silencing, or RNA interference (RNAi), in animal extracts (reviewed in Ref. 23). RNAi is believed to operate in a similar manner in plants because small interfering RNAs (si. RNAs) are found in silenced plants, and plants have homologues of the animal gene Dicer. Double-stranded RNA (ds. RNA) from replicating viral RNA, viralvector-derived (VIGS, or virus-induced gene silencing) RNA or hairpin RNA (hp. RNA) transcribed from a transgene, is processed by a Dicer-containing complex to generate si. RNAs. An endonucleasecontaining complex (called the RNAi silencing complex, RISC), is guided by the antisense strand of the si. RNA to cleave specific m. RNAs, so promoting their degradation.
vitro studies of RNA-induced gene silencing, or RNA This model is based on the results of in vitro studies of RNA-induced gene silencing, or RNA interference (RNAi), in animal extracts (reviewed in Ref. 23). RNAi is believed to operate in a similar manner in plants because small interfering RNAs (si. RNAs) are found in silenced plants, and plants have homologues of the animal gene Dicer. Double-stranded RNA (ds. RNA) from replicating viral RNA, viralvector-derived (VIGS, or virus-induced gene silencing) RNA or hairpin RNA (hp. RNA) transcribed from a transgene, is processed by a Dicer-containing complex to generate si. RNAs. An endonucleasecontaining complex (called the RNAi silencing complex, RISC), is guided by the antisense strand of the si. RNA to cleave specific m. RNAs, so promoting their degradation.
 si. RNA ve mi. RNA • Her ikisi de Dicer adlı bir enzimle daha uzun RNA lardan oluşturulur. • Bu RNAse III e benzer bir enzimdir. • Uzun ds. RNA veya mi. RNA öncüllerinin oluşturduğu sap-ilmik yapılarını tanır. • si. RNA ve mi. RNA 21 -23 nükleotid uzunluktadır. Bunlar, homolog hedef genlerin ekspresyonunu 3 şekilde baskılar.
si. RNA ve mi. RNA • Her ikisi de Dicer adlı bir enzimle daha uzun RNA lardan oluşturulur. • Bu RNAse III e benzer bir enzimdir. • Uzun ds. RNA veya mi. RNA öncüllerinin oluşturduğu sap-ilmik yapılarını tanır. • si. RNA ve mi. RNA 21 -23 nükleotid uzunluktadır. Bunlar, homolog hedef genlerin ekspresyonunu 3 şekilde baskılar.
 • m. RNA nın yıkılmasını tetikler • m. RNA translasyonunu inhibe eder veya • Gen içindeki Kromatin modifikasyonlarını indükleyerek transkripsiyonu susturur. • Hangi yol kullanılırsa kullanılsın RISC gereklidir
• m. RNA nın yıkılmasını tetikler • m. RNA translasyonunu inhibe eder veya • Gen içindeki Kromatin modifikasyonlarını indükleyerek transkripsiyonu susturur. • Hangi yol kullanılırsa kullanılsın RISC gereklidir
 RNA susturulması Bir gene homoloji bölgesi içeren bir çift iplikli RNA molekülü hücreye girdiğinde veya hücrede Yapıldığında genin ekspresyonu Durdurulur. Bu etki çift iplikli RNA nın si. RNA veya mi. RNA oluşturmak üzere Dicer denilen enzimle İşlenmesini içerir. Mi. RNA oluşumuna ise Drosha adlı bir diğer enzim daha katılır. si. RNA ve mi. RNA genleri 3 yoldan susturmak üzere RISC adlı komplekse gider 1 -Si. RNA ile homoloji taşıyan m. RNA ya saldırır ve sindirir 2 - m. RNA nın translasyonunu önler veya 3 - Kromatin modifiye eden enzimleri m. RNA nın Ekspresyonunu yöneten promotöre yönlendirir
RNA susturulması Bir gene homoloji bölgesi içeren bir çift iplikli RNA molekülü hücreye girdiğinde veya hücrede Yapıldığında genin ekspresyonu Durdurulur. Bu etki çift iplikli RNA nın si. RNA veya mi. RNA oluşturmak üzere Dicer denilen enzimle İşlenmesini içerir. Mi. RNA oluşumuna ise Drosha adlı bir diğer enzim daha katılır. si. RNA ve mi. RNA genleri 3 yoldan susturmak üzere RISC adlı komplekse gider 1 -Si. RNA ile homoloji taşıyan m. RNA ya saldırır ve sindirir 2 - m. RNA nın translasyonunu önler veya 3 - Kromatin modifiye eden enzimleri m. RNA nın Ekspresyonunu yöneten promotöre yönlendirir
 • RISC mi. RNA veya si. RNA ya ek olarak Argonaute ailesinden proteinler de içerir Argonaute katalitik alt birimdir m. RNA nın ilk kesilmesini gerçekleştirir (Slicer da denir) • Eğer dizi çok iyi komplementer ise hedef yıkılır • Eğer eşleşme iyi değil ise ( birkaç bç eşleşmemişse) • translasyon inhibisyonu olur • RNAİ susturması çok verimlidir • Çok az miktarda ds RNA hedek genlerin ekspresyonunun nerdeyse tamamen durdurulmasını indüklemek için yeterlidir
• RISC mi. RNA veya si. RNA ya ek olarak Argonaute ailesinden proteinler de içerir Argonaute katalitik alt birimdir m. RNA nın ilk kesilmesini gerçekleştirir (Slicer da denir) • Eğer dizi çok iyi komplementer ise hedef yıkılır • Eğer eşleşme iyi değil ise ( birkaç bç eşleşmemişse) • translasyon inhibisyonu olur • RNAİ susturması çok verimlidir • Çok az miktarda ds RNA hedek genlerin ekspresyonunun nerdeyse tamamen durdurulmasını indüklemek için yeterlidir
 Sinir Sistemi ve mi. RNA’lar Posttranskripsiyonel düzenleme • Nöron gelişimi ve • MSS işlevlerine katılan bir mekanizmadır • Bu yolağın karmaşıklığı mi. RNA’ların keşfi ile daha iyi anlaşıldı. Bunlar kodlayıcı olmayan bir küçük RNA sınıfıdır. • Hedef m. RNA’nın translasyonunu baskılar veya • Yıkılmasını indükler (m. RNA’nın 3’ UTR bölgesine baz eşleşmesi tam ise)
Sinir Sistemi ve mi. RNA’lar Posttranskripsiyonel düzenleme • Nöron gelişimi ve • MSS işlevlerine katılan bir mekanizmadır • Bu yolağın karmaşıklığı mi. RNA’ların keşfi ile daha iyi anlaşıldı. Bunlar kodlayıcı olmayan bir küçük RNA sınıfıdır. • Hedef m. RNA’nın translasyonunu baskılar veya • Yıkılmasını indükler (m. RNA’nın 3’ UTR bölgesine baz eşleşmesi tam ise)
 Susturma mekanizmaları 1. Translasyonun başlamasını önlemek • mi. RNA aracılı deadenilasyon ve yıkılma • • • İşlenme cisimleri (processing) P-cisimcikleri m. RNA depolanma ve yıkılma yerleri MSS, mi. RNA’larca çok zengin ve beyine özgü Tek mi. RNA birkaç 100 farklı m. RNA’yı hedefleyebilir. MSS gelişimi ve işlevlerinde çok sayıda rolü var.
Susturma mekanizmaları 1. Translasyonun başlamasını önlemek • mi. RNA aracılı deadenilasyon ve yıkılma • • • İşlenme cisimleri (processing) P-cisimcikleri m. RNA depolanma ve yıkılma yerleri MSS, mi. RNA’larca çok zengin ve beyine özgü Tek mi. RNA birkaç 100 farklı m. RNA’yı hedefleyebilir. MSS gelişimi ve işlevlerinde çok sayıda rolü var.
 Sinir Sisteminde mi. RNA ekspresyonu Beyinden ve nöral hücre hatlarından • Çok sayıda benzeri olmayan mi. RNA izole edildi. • Çoğu poliribozomlarla ilişkili • mi. RNA’ların translasyonun düzenlenmesine katıldığını gösterir • Mikrodizin teknolojisi ile mi. RNA ekspresyonunun sinir sisteminde dokuya özgü şekilde dinamik olarak düzenlendiği gösterildi MSS’de • mir-138 • mir-124 çok fazla ifade edilir.
Sinir Sisteminde mi. RNA ekspresyonu Beyinden ve nöral hücre hatlarından • Çok sayıda benzeri olmayan mi. RNA izole edildi. • Çoğu poliribozomlarla ilişkili • mi. RNA’ların translasyonun düzenlenmesine katıldığını gösterir • Mikrodizin teknolojisi ile mi. RNA ekspresyonunun sinir sisteminde dokuya özgü şekilde dinamik olarak düzenlendiği gösterildi MSS’de • mir-138 • mir-124 çok fazla ifade edilir.
 mir-29 astrositlerde") • Bazısı bölgeye veya hücreye özgü; mir-222 sadece telensefalonda (zebra balığında) mir-29 astrositlerde • Bazısı gelişim sırasunda düzenlenir. mir-92 b nöronal progenitör hücrelerde yüksek Düzeyde iken farklılaşmış nöronlarda yok , veya tam tersi. Örneğin; mir-124
• Bazısı bölgeye veya hücreye özgü; mir-222 sadece telensefalonda (zebra balığında) mir-29 astrositlerde • Bazısı gelişim sırasunda düzenlenir. mir-92 b nöronal progenitör hücrelerde yüksek Düzeyde iken farklılaşmış nöronlarda yok , veya tam tersi. Örneğin; mir-124

") Mechanism of RNA interference. (Reprinted by permission of Phillip D. Zamore. )
Mechanism of RNA interference. (Reprinted by permission of Phillip D. Zamore. )
 . . vitro studies of RNA-induced gene silencing, or RNA 600 × 738
. . vitro studies of RNA-induced gene silencing, or RNA 600 × 738
 bir pre-micro. RNA'nın sap-ilmik yapısı.
bir pre-micro. RNA'nın sap-ilmik yapısı.
 mi. RNA'yı kodlayan genler işlenmiş olgun mi. RNA molekülünden çok daha uzundur. mi. RNA'lar önce birincil (primer) transkript, veya pri-mi. RNA olarak yazılırlar, bu transkriptlerin birer başlığı ve poli-A kuyruğu vardır. Bunlar işlem görüp hücre çekirdeğinde pre-mi. RNA olarak bilinen kısa, 70 nükleotit uzunlukta, sap-ilmik şekilli, öncül (prekürsör) yapılara dönüşür. Bu işlenme hayvanlarda Mikroişlemci kompleks (İng. Microprocessor complex) adlı bir protein kompleksi tarafından gerçekleştirilir. Mikroişlemci kompleks'te Drosha adlı bir nükleaz, ve Pasha adlı çift iplikli RNA bağlayıcı protein bulunur. [3] Pre-mi. RNA'lar sonra sitoplazmada Dicer adlı endonükleaz ile etkileşerek olgun mi. RNA'ya dönüşürler. Dicer aynı zamanda RNA-indüklenmiş susturma kompleksi (İng. RNA-induced silencing complex; RISC) oluşumunun başlatır. [4] Bu kompleks mi. RNA ifadesi ve RNA interferanstan kaynaklanan gen susturmasından
mi. RNA'yı kodlayan genler işlenmiş olgun mi. RNA molekülünden çok daha uzundur. mi. RNA'lar önce birincil (primer) transkript, veya pri-mi. RNA olarak yazılırlar, bu transkriptlerin birer başlığı ve poli-A kuyruğu vardır. Bunlar işlem görüp hücre çekirdeğinde pre-mi. RNA olarak bilinen kısa, 70 nükleotit uzunlukta, sap-ilmik şekilli, öncül (prekürsör) yapılara dönüşür. Bu işlenme hayvanlarda Mikroişlemci kompleks (İng. Microprocessor complex) adlı bir protein kompleksi tarafından gerçekleştirilir. Mikroişlemci kompleks'te Drosha adlı bir nükleaz, ve Pasha adlı çift iplikli RNA bağlayıcı protein bulunur. [3] Pre-mi. RNA'lar sonra sitoplazmada Dicer adlı endonükleaz ile etkileşerek olgun mi. RNA'ya dönüşürler. Dicer aynı zamanda RNA-indüklenmiş susturma kompleksi (İng. RNA-induced silencing complex; RISC) oluşumunun başlatır. [4] Bu kompleks mi. RNA ifadesi ve RNA interferanstan kaynaklanan gen susturmasından
 f. m 360 × 356 -. . . i R transcripts occurs through a multi-step process that is N A _ P r o c e s s i n g _ a n d _ A c t i v i t y _
f. m 360 × 356 -. . . i R transcripts occurs through a multi-step process that is N A _ P r o c e s s i n g _ a n d _ A c t i v i t y _
 421 × 463 RNAi pathway
421 × 463 RNAi pathway