

схема регуляции генной активности.ppt
- Количество слайдов: 31
 Тема: Регуляция экспрессии генов у про- и эукариот
Тема: Регуляция экспрессии генов у про- и эукариот
 План лекции: 1. Общие принципы и механизмы регуляции активности генов. 2. Регуляция активности генов у прокариот. 3. Регуляция активности генов у эукариот. 4. Уровни регуляции активности генов и их характеристики
План лекции: 1. Общие принципы и механизмы регуляции активности генов. 2. Регуляция активности генов у прокариот. 3. Регуляция активности генов у эукариот. 4. Уровни регуляции активности генов и их характеристики
 генов позволяет приспособить организм к изменяющимся условиям среды. В клетке") Регуляция активности (экспрессии) генов позволяет приспособить организм к изменяющимся условиям среды. В клетке в каждый момент ее существования должно синтезироваться и функционировать необходимое количество различных белков.
Регуляция активности (экспрессии) генов позволяет приспособить организм к изменяющимся условиям среды. В клетке в каждый момент ее существования должно синтезироваться и функционировать необходимое количество различных белков.
 У бактерий набор белков регулируется наличием м-РНК, пригодных для биосинтеза белка. Так как процессы транскрипции и трансляции протекают почти одновременно, то внести какие-либо изменения в структуру м-РНК практически невозможно.
У бактерий набор белков регулируется наличием м-РНК, пригодных для биосинтеза белка. Так как процессы транскрипции и трансляции протекают почти одновременно, то внести какие-либо изменения в структуру м-РНК практически невозможно.
 У эукариот молекулы РНК подвергаются существенным модификациям благодаря ядерной мембране, разделяющей зоны транскрипции и трансляции.
У эукариот молекулы РНК подвергаются существенным модификациям благодаря ядерной мембране, разделяющей зоны транскрипции и трансляции.
 В любом случае первым и главным элементом контроля реализации генетической информации от ДНК к белку у всех организмов является контроль на уровне транскрипции.
В любом случае первым и главным элементом контроля реализации генетической информации от ДНК к белку у всех организмов является контроль на уровне транскрипции.
 Регуляция экспрессии у прокариот Регуляция генетической активности у прокариот идет на уровне транскрипции. В 1961 г. франц. ученые Жакоб и Моно предложили модель оперона как систему регуляции экспрессии генов бактерий.
Регуляция экспрессии у прокариот Регуляция генетической активности у прокариот идет на уровне транскрипции. В 1961 г. франц. ученые Жакоб и Моно предложили модель оперона как систему регуляции экспрессии генов бактерий.
 Оперон Промотор- место связывания РНК-П Оператор- место связывания белкарепрессора Структурные гены- кодируют ферменты ----------------------------Ген-регулятор кодирует белокрепрессор. Он не входит в состав оперона.
Оперон Промотор- место связывания РНК-П Оператор- место связывания белкарепрессора Структурные гены- кодируют ферменты ----------------------------Ген-регулятор кодирует белокрепрессор. Он не входит в состав оперона.
 В отсутствие этого субстрата белокрепрессор имеет") Индуцибельные опероны Регулятором является исходный субстрат (S ) В отсутствие этого субстрата белокрепрессор имеет высокое сродство к оператору, поэтому РНК-П не может начать транскрипцию (оперон выключен). При накоплении метаболита (S) в клетке некоторое его количество связывается с белком-репрессором и понижает его сродство к оператору. Оперон включается и синтезируются ферменты для расщепления S.
Индуцибельные опероны Регулятором является исходный субстрат (S ) В отсутствие этого субстрата белокрепрессор имеет высокое сродство к оператору, поэтому РНК-П не может начать транскрипцию (оперон выключен). При накоплении метаболита (S) в клетке некоторое его количество связывается с белком-репрессором и понижает его сродство к оператору. Оперон включается и синтезируются ферменты для расщепления S.
 В отсутствие этого продукта белокрепрессор имеет низкое") Репрессибельные опероны Регулятором служит конечный продукт (Р) В отсутствие этого продукта белокрепрессор имеет низкое сродство к оператору, поэтому РНК-П транскрибирует гены оперона. Оперон включен и синтезируются ферменты, способствующие образованию вещества Р. При накоплении данного в-ва некоторое его кол-во связывается с белком-репрессором и повышает его сродство к оператору. Оперон выключается и синтез ферментов прекращается.
Репрессибельные опероны Регулятором служит конечный продукт (Р) В отсутствие этого продукта белокрепрессор имеет низкое сродство к оператору, поэтому РНК-П транскрибирует гены оперона. Оперон включен и синтезируются ферменты, способствующие образованию вещества Р. При накоплении данного в-ва некоторое его кол-во связывается с белком-репрессором и повышает его сродство к оператору. Оперон выключается и синтез ферментов прекращается.
 Способы регуляции экспрессии генов прокариот Регуляция связывания РНК-П с промоторами Регуляция перемещения связавшейся РНК-П от промотора к собственно генам
Способы регуляции экспрессии генов прокариот Регуляция связывания РНК-П с промоторами Регуляция перемещения связавшейся РНК-П от промотора к собственно генам
 Лактозный оперон Лактоза- дисахарид, на котором растут клетки бактерий Lac Z – галактозидазу для расщепления лактозы до глюкозы и галактозы Lac Y- пермеазу для проникновения лактозы из внешней среды в клетку Lac A - трансацетилазу
Лактозный оперон Лактоза- дисахарид, на котором растут клетки бактерий Lac Z – галактозидазу для расщепления лактозы до глюкозы и галактозы Lac Y- пермеазу для проникновения лактозы из внешней среды в клетку Lac A - трансацетилазу
 Организация и функционирование Lac – оперона по Жакобу и Моно
Организация и функционирование Lac – оперона по Жакобу и Моно
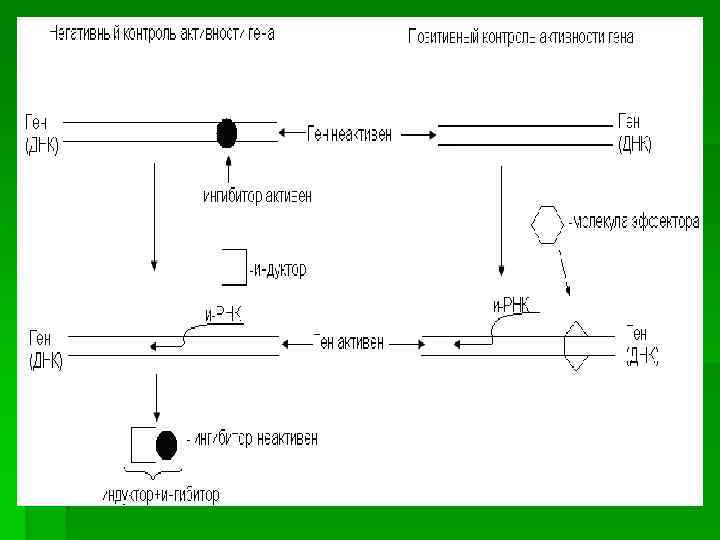
 Организация и функционирование Lac – оперона по Жакобу и Моно
Организация и функционирование Lac – оперона по Жакобу и Моно
 Функционирование лактозного оперона у E. Coli: а– локализация сайтов связывания молекул РНК – полимеразы и репрессора в регуляторный области гена lac Z [ Lewin, 1994. P. 417]; б – структура лактозного оперона (как и у всех генов, регулируемых САР и с. АМР, промотор содержит два района: участок связывания с РНК-полимеразой и участок связывания с комплексом САР и с. АМР); в – негативная, г – позитивная регуляция lac-оперона [ б-г – Curtis, Barnes, 1989. P. 325].
Функционирование лактозного оперона у E. Coli: а– локализация сайтов связывания молекул РНК – полимеразы и репрессора в регуляторный области гена lac Z [ Lewin, 1994. P. 417]; б – структура лактозного оперона (как и у всех генов, регулируемых САР и с. АМР, промотор содержит два района: участок связывания с РНК-полимеразой и участок связывания с комплексом САР и с. АМР); в – негативная, г – позитивная регуляция lac-оперона [ б-г – Curtis, Barnes, 1989. P. 325].
 связывается с белкомактиватором катаболизма (БАК, или") В лактозном опероне левая часть промотора (активатор) связывается с белкомактиватором катаболизма (БАК, или САР в англ. терминологии), а правая часть – с РНКП. CАP-белок в отличие от белкарепрессора играет позитивную роль, помогая РНК-П начать транскрипцию. Связывание CАP с промотором происходит если CАP соединяется с ц. АМФ.
В лактозном опероне левая часть промотора (активатор) связывается с белкомактиватором катаболизма (БАК, или САР в англ. терминологии), а правая часть – с РНКП. CАP-белок в отличие от белкарепрессора играет позитивную роль, помогая РНК-П начать транскрипцию. Связывание CАP с промотором происходит если CАP соединяется с ц. АМФ.
 Глюкозы нет Если источником энергии служит лактоза и во внешней среде нет глюкозы, репрессор инактивируется. При этом промотор открыт для РНК-П, транскрипция идет. .
Глюкозы нет Если источником энергии служит лактоза и во внешней среде нет глюкозы, репрессор инактивируется. При этом промотор открыт для РНК-П, транскрипция идет. .
 Глюкоза есть Если во внешней среде достаточно глюкозы, то биологического смысла в использовании лактозы уже нет. Поэтому глюкоза препятствует активации лактозного оперона даже в присутствии больших количеств лактозы.
Глюкоза есть Если во внешней среде достаточно глюкозы, то биологического смысла в использовании лактозы уже нет. Поэтому глюкоза препятствует активации лактозного оперона даже в присутствии больших количеств лактозы.
 , поэтому CАP") В отсутствие глюкозы в клетке уровень ц. АМФ возрастает (сигнал голода) , поэтому CАP связан с лактозным промотором, к которому легко присоединяется РНК-П. В этих условиях активность оперона зависит только от того, свободен или нет оператор, т. е. от наличия во внешней среде лактозы.
В отсутствие глюкозы в клетке уровень ц. АМФ возрастает (сигнал голода) , поэтому CАP связан с лактозным промотором, к которому легко присоединяется РНК-П. В этих условиях активность оперона зависит только от того, свободен или нет оператор, т. е. от наличия во внешней среде лактозы.
 В присутствии глюкозы Уровень ц. АМФ остается низким, промотор остается без CАP и теряет сродство к РНК-П. Лактозный оперон не работает и в качестве источника питания используется глюкоза.
В присутствии глюкозы Уровень ц. АМФ остается низким, промотор остается без CАP и теряет сродство к РНК-П. Лактозный оперон не работает и в качестве источника питания используется глюкоза.
 Триптофановый оперон Это репрессибельный оперон с двойным механизмом регуляции: 1. регулируется перемещение РНК-П по оператору 2. окончание транскрипции на аттенюаторе
Триптофановый оперон Это репрессибельный оперон с двойным механизмом регуляции: 1. регулируется перемещение РНК-П по оператору 2. окончание транскрипции на аттенюаторе
 Т. О. состоит из 5 цистронов. За промотором и оператором находится лидерный отдел, который заканчивается аттенюатором. При транскрипции этого отдела образуется лидерный участок м. РНК. При трансляции образуется лидерный пептид.
Т. О. состоит из 5 цистронов. За промотором и оператором находится лидерный отдел, который заканчивается аттенюатором. При транскрипции этого отдела образуется лидерный участок м. РНК. При трансляции образуется лидерный пептид.
 Особенность лидерного пептида: среди его 14 аминокислотных остатков содержатся 2 остатка триптофана, т. е. той самой аминокислоты, синтез которой контролируется опероном.
Особенность лидерного пептида: среди его 14 аминокислотных остатков содержатся 2 остатка триптофана, т. е. той самой аминокислоты, синтез которой контролируется опероном.
 Триптофан есть Синтез лидерного пептида идет без задержки. При достижении РНК-П аттенюатора срабатывает сигнал об окончании транскрипции.
Триптофан есть Синтез лидерного пептида идет без задержки. При достижении РНК-П аттенюатора срабатывает сигнал об окончании транскрипции.
 Триптофана нет Синтез лидерного пептида задерживается и сигнал об окончании трансляции на аттенюаторе не срабатывает и синтезируются ферменты, необходимые для пополнения запаса триптофана в клетке.
Триптофана нет Синтез лидерного пептида задерживается и сигнал об окончании трансляции на аттенюаторе не срабатывает и синтезируются ферменты, необходимые для пополнения запаса триптофана в клетке.
 Аттенюатор Это участок ДНК между оператором и генами, на котором при определенных условиях прекращается транскрипция оперона. За промотором и оператором находится лидерный отдел, именно он оканчивается аттенюатором.
Аттенюатор Это участок ДНК между оператором и генами, на котором при определенных условиях прекращается транскрипция оперона. За промотором и оператором находится лидерный отдел, именно он оканчивается аттенюатором.
 Когда триптофана мало Синтез ЛП задерживается и сигнал об окончании транскрипции на аттенюаторе не срабатывает, поэтому синтезируются ферменты, необходимые для пополнения запаса триптофана в клетке.
Когда триптофана мало Синтез ЛП задерживается и сигнал об окончании транскрипции на аттенюаторе не срабатывает, поэтому синтезируются ферменты, необходимые для пополнения запаса триптофана в клетке.
 Координированная репрессия у Salmonella typhimurium
Координированная репрессия у Salmonella typhimurium
 Литература: 1. Биология. Под ред. Ярыгина В. Н. М. , 2001. 2. Бочков Н. П. и др. Медицинская генетика. М. , 1984. 3. Генетика. Под ред. Иванова В. И. М. , 2006. 4. Заяц Р. Г. и др. Общая и медицинская генетика. Ростов-на-Дону, 2002. 5. Инге-Вечтомов С. Г. Генетика с основами селекции. М. , 1989. 6. Кемп П. , Армс К. Введение в биологию. М. , 1988. 7. Медицинская биология и генетика. Под ред. проф. Куандыкова Е. У. Алматы, 2004. 8. Фаллер Д. М. , Шилдс Д. Молекулярная биология клетки. М. , 2006. 9. Муминов Т. А. , Куандыков Е. У. Основы молекулярной биологии (курс лекций). Алматы, 2007. 10. Мушкамбаров Н. Н. , Кузнецов С. Л. Молекулярная биология. М. , 2003.
Литература: 1. Биология. Под ред. Ярыгина В. Н. М. , 2001. 2. Бочков Н. П. и др. Медицинская генетика. М. , 1984. 3. Генетика. Под ред. Иванова В. И. М. , 2006. 4. Заяц Р. Г. и др. Общая и медицинская генетика. Ростов-на-Дону, 2002. 5. Инге-Вечтомов С. Г. Генетика с основами селекции. М. , 1989. 6. Кемп П. , Армс К. Введение в биологию. М. , 1988. 7. Медицинская биология и генетика. Под ред. проф. Куандыкова Е. У. Алматы, 2004. 8. Фаллер Д. М. , Шилдс Д. Молекулярная биология клетки. М. , 2006. 9. Муминов Т. А. , Куандыков Е. У. Основы молекулярной биологии (курс лекций). Алматы, 2007. 10. Мушкамбаров Н. Н. , Кузнецов С. Л. Молекулярная биология. М. , 2003.
: 1. Что такое оперон? 2. Уровни регуляции активности генов у") Контрольные вопросы (обратная связь): 1. Что такое оперон? 2. Уровни регуляции активности генов у прокариот. 3. Механизмы регуляции активности генов у прокариот. 4. Особенности регуляции активности генов у эукариот. 5. Дифференциальная экспрессия генов и ее значение в жизнедеятельности организмов.
Контрольные вопросы (обратная связь): 1. Что такое оперон? 2. Уровни регуляции активности генов у прокариот. 3. Механизмы регуляции активности генов у прокариот. 4. Особенности регуляции активности генов у эукариот. 5. Дифференциальная экспрессия генов и ее значение в жизнедеятельности организмов.