

Занятие 2. Цитология.ppt
- Количество слайдов: 143
 Тема 2: Цитология. Межклеточные контакты. Органеллы. Ядро. Деление клетки.
Тема 2: Цитология. Межклеточные контакты. Органеллы. Ядро. Деление клетки.
 МЕЖКЛЕТОЧНЫЕ СОЕДИНЕНИЯ • • • Внешние клеточные мембраны участвуют в образовании межклеточных контактов, которые обеспечивают межклеточные взаимодействия. Простое межклеточное соединение – сближение плазмолемм соседних клеток на расстояние 15 -20 нм. Основная роль принадлежит клеточным адгезионным молекулам (КАМ), таким как кадгерины, интегрины, способные распознавать и связывать плазмолеммы соседних клеток.
МЕЖКЛЕТОЧНЫЕ СОЕДИНЕНИЯ • • • Внешние клеточные мембраны участвуют в образовании межклеточных контактов, которые обеспечивают межклеточные взаимодействия. Простое межклеточное соединение – сближение плазмолемм соседних клеток на расстояние 15 -20 нм. Основная роль принадлежит клеточным адгезионным молекулам (КАМ), таким как кадгерины, интегрины, способные распознавать и связывать плазмолеммы соседних клеток.
 • Сложные межклеточные соединения – небольшие парные специализированные участки плазматических мембран соседних клеток. • изолирующие (запирающие) (Плотный контакт), • сцепляющие, обусловливающие механическое сцепление и соединение клеток (поясок сцепления , десмосома) • коммуникационные соединения, обеспечивающие химическую (метаболическую, ионную) и электрическую связь между клетками.
• Сложные межклеточные соединения – небольшие парные специализированные участки плазматических мембран соседних клеток. • изолирующие (запирающие) (Плотный контакт), • сцепляющие, обусловливающие механическое сцепление и соединение клеток (поясок сцепления , десмосома) • коммуникационные соединения, обеспечивающие химическую (метаболическую, ионную) и электрическую связь между клетками.
 контакты • Плотный контакт (zonula occludens)") • К изолирующим соединениям относятся плотные (изоляционные) контакты • Плотный контакт (zonula occludens) обеспечивают максимальное сближение мембран соседних клеток, между которыми остается промежуток в 2 -3 нм. ( чаще всего возникает в эпителии) • . Плотные контакты образуют непрерывные пояса вокруг каждой клетки, крепко прижимая их друг к другу и предотвращая протекание межклеточной жидкости между ними. Такие контакты необходимы, в частности, для обеспечения водонепроницаемости кожи. В формировании тесных контактов принимают участие белки окклюдины, клаудины и другие
• К изолирующим соединениям относятся плотные (изоляционные) контакты • Плотный контакт (zonula occludens) обеспечивают максимальное сближение мембран соседних клеток, между которыми остается промежуток в 2 -3 нм. ( чаще всего возникает в эпителии) • . Плотные контакты образуют непрерывные пояса вокруг каждой клетки, крепко прижимая их друг к другу и предотвращая протекание межклеточной жидкости между ними. Такие контакты необходимы, в частности, для обеспечения водонепроницаемости кожи. В формировании тесных контактов принимают участие белки окклюдины, клаудины и другие
 соединениям относят поясок сцепления и десмосомы. • Якорные контакты") • К сцепляющим (заякоривающим) соединениям относят поясок сцепления и десмосомы. • Якорные контакты физически соединяют клетки между собой, обеспечивают целостность и прочность тканей, в частности эпителиальных и мышечных. • элементы цитоскелета соседних клеток объединяются в единую структуру: с помощью якорных белков кадгенринов, они прикрепляются в межклеточном пространстве к кадгеринам соседних клеток. • Два основных типа: адгезионные, объединяющие микрофиламенты соседних клеток; и десмосомы, в образовании которых принимают участие промежуточные филаменты.
• К сцепляющим (заякоривающим) соединениям относят поясок сцепления и десмосомы. • Якорные контакты физически соединяют клетки между собой, обеспечивают целостность и прочность тканей, в частности эпителиальных и мышечных. • элементы цитоскелета соседних клеток объединяются в единую структуру: с помощью якорных белков кадгенринов, они прикрепляются в межклеточном пространстве к кадгеринам соседних клеток. • Два основных типа: адгезионные, объединяющие микрофиламенты соседних клеток; и десмосомы, в образовании которых принимают участие промежуточные филаменты.
 также опоясывает клетки в виде ленты,") • Поясок сцепления (zonula adherens, опоясывающая десмосома) также опоясывает клетки в виде ленты, но локализуется на латеральной поверхности клеточной мембраны ниже, чем плотный контакт. Здесь клетки связаны друг с другом интегральными гликопротеидами, к которым примыкает слой примембранных белков (винкулин и др. ).
• Поясок сцепления (zonula adherens, опоясывающая десмосома) также опоясывает клетки в виде ленты, но локализуется на латеральной поверхности клеточной мембраны ниже, чем плотный контакт. Здесь клетки связаны друг с другом интегральными гликопротеидами, к которым примыкает слой примембранных белков (винкулин и др. ).

- парная структура, состоящая из уплотненных участков цитоплазмы, прилегающих к") Десмосома • Десмосома (macula adherens)- парная структура, состоящая из уплотненных участков цитоплазмы, прилегающих к плазмолеммам соседних клеток, так называемых пластинок прикрепления, разделенных межклеточной щелью. • Каждая пластинка прикрепления имеет форму диска (диаметр около 0. 5 мкм) и содержит особые белки (десмоплакины и др. ), к которым прикреплены пучки промежуточных филаментов (тонофиламентов). При этом находящиеся в межклеточном пространстве Са 2+-связывающие белки взаимодействуют с пластинками прикрепления, благодаря чему усиливается механическое сцепление клеток. • Десмосомы не имеют определенной локализации и разбросаны по поверхности клетки
Десмосома • Десмосома (macula adherens)- парная структура, состоящая из уплотненных участков цитоплазмы, прилегающих к плазмолеммам соседних клеток, так называемых пластинок прикрепления, разделенных межклеточной щелью. • Каждая пластинка прикрепления имеет форму диска (диаметр около 0. 5 мкм) и содержит особые белки (десмоплакины и др. ), к которым прикреплены пучки промежуточных филаментов (тонофиламентов). При этом находящиеся в межклеточном пространстве Са 2+-связывающие белки взаимодействуют с пластинками прикрепления, благодаря чему усиливается механическое сцепление клеток. • Десмосомы не имеют определенной локализации и разбросаны по поверхности клетки
 Десмосомы
Десмосомы

 представляет") • Коммуникационные соединения представлены щелевыми контактами и синапсами. • Щелевое соединение (нексус) представляет собой участок протяженностью 0, 5 -3 мкм, где плазмолеммы разделены узкой межклеточной щелью (2 -3 нм). При этом в структуре плазмолемм соседних клеток друг против друга располагаются трубчатые трансмембранные структуры – коннексоны (из белка коннексина), которые образуют межцитоплазматические каналы, обеспечивающие свободный обмен низкомолекулярными соединениями между клетками. • Число коннексонов в одном щелевом контакте обычно исчисляется сотнями. • Функциональная роль щелевых соединений заключается в переносе ионов и мелких молекул от клетки к клетке.
• Коммуникационные соединения представлены щелевыми контактами и синапсами. • Щелевое соединение (нексус) представляет собой участок протяженностью 0, 5 -3 мкм, где плазмолеммы разделены узкой межклеточной щелью (2 -3 нм). При этом в структуре плазмолемм соседних клеток друг против друга располагаются трубчатые трансмембранные структуры – коннексоны (из белка коннексина), которые образуют межцитоплазматические каналы, обеспечивающие свободный обмен низкомолекулярными соединениями между клетками. • Число коннексонов в одном щелевом контакте обычно исчисляется сотнями. • Функциональная роль щелевых соединений заключается в переносе ионов и мелких молекул от клетки к клетке.
 Синаптические соединения • Высокоспециализированные • контакты нервных клеток, • проводящие импульсы в одном • направлении. • Синаптические контакты • устанавливаются также между • нейронами и мышечными или • железистыми клетками
Синаптические соединения • Высокоспециализированные • контакты нервных клеток, • проводящие импульсы в одном • направлении. • Синаптические контакты • устанавливаются также между • нейронами и мышечными или • железистыми клетками
 Органеллы
Органеллы
 • Органеллы – постоянно присутствующие в цитоплазме структуры, имеющие определенное строение и специализированные на выполнении определенных функций в клетке. • Они подразделяются на • органеллы общего значения • специальные органеллы.
• Органеллы – постоянно присутствующие в цитоплазме структуры, имеющие определенное строение и специализированные на выполнении определенных функций в клетке. • Они подразделяются на • органеллы общего значения • специальные органеллы.
 • • Органеллы общего значения имеются во всех клетках и необходимы для обеспечения их жизнедеятельности. К ним относятся: • эндоплазматическая сеть (ЭПС), • митохондрии, • рибосомы • комплекс Гольджи • лизосомы • пероксисомы, • клеточный центр • компоненты цитоскелета.
• • Органеллы общего значения имеются во всех клетках и необходимы для обеспечения их жизнедеятельности. К ним относятся: • эндоплазматическая сеть (ЭПС), • митохондрии, • рибосомы • комплекс Гольджи • лизосомы • пероксисомы, • клеточный центр • компоненты цитоскелета.
 • Специальные органеллы содержатся лишь в некоторых специализированных клетках, где они обеспечивают выполнение специальных функций. • К специальным органеллам относят реснички, жгутики, миофибриллы, акросома. • Все специальные органеллы образуются при развитии клетки как производные органелл общего значения, так, например, акросома спермия является производным комплекса Гольджи, реснички и жгутики – микротрубочек цитоскелета и т. д
• Специальные органеллы содержатся лишь в некоторых специализированных клетках, где они обеспечивают выполнение специальных функций. • К специальным органеллам относят реснички, жгутики, миофибриллы, акросома. • Все специальные органеллы образуются при развитии клетки как производные органелл общего значения, так, например, акросома спермия является производным комплекса Гольджи, реснички и жгутики – микротрубочек цитоскелета и т. д
 • В состав многих органелл входит элементарная биологическая мембрана, поэтому органеллы подразделяются также на • мембранные и • немембранные. • Мембранные органеллы: митохондрии, ЭПС, комплекс Гольджи, лизосомы, пероксисомы; • Немембранные органеллы: рибосомы, клеточный центр, компоненты цитоскелета, микроворсинки, реснички, жгутики
• В состав многих органелл входит элементарная биологическая мембрана, поэтому органеллы подразделяются также на • мембранные и • немембранные. • Мембранные органеллы: митохондрии, ЭПС, комплекс Гольджи, лизосомы, пероксисомы; • Немембранные органеллы: рибосомы, клеточный центр, компоненты цитоскелета, микроворсинки, реснички, жгутики
 СИНТЕТИЧЕСКИЙ АППАРАТ КЛЕТКИ • Синтетический аппарат клеток включает органеллы, участвующие в синтезе различных веществ. • К таким органеллам относятся • рибосомы, • эндоплазматическая сеть и • комплекс Гольджи. • Деятельность синтетического аппарата клетки контролируется активностью генов, локализованных в ядре.
СИНТЕТИЧЕСКИЙ АППАРАТ КЛЕТКИ • Синтетический аппарат клеток включает органеллы, участвующие в синтезе различных веществ. • К таким органеллам относятся • рибосомы, • эндоплазматическая сеть и • комплекс Гольджи. • Деятельность синтетического аппарата клетки контролируется активностью генов, локализованных в ядре.
 • Рибосомы – мелкие, плотные немембранные органеллы, диаметром 15 -30 нм. • Функция рибосом – синтез белка путем соединения аминокислот в полипептидные цепочки • Каждая рибосома состоит из двух субъединиц: большой и малой.
• Рибосомы – мелкие, плотные немембранные органеллы, диаметром 15 -30 нм. • Функция рибосом – синтез белка путем соединения аминокислот в полипептидные цепочки • Каждая рибосома состоит из двух субъединиц: большой и малой.
 • Субъединицы рибосом собираются в ядре из р. РНК, которая образуется в ядрышке, и белков, которые синтезируются в цитоплазме и поступают в ядро. Затем субъединицы рибосом через ядерные поры перемещаются в цитоплазму, где они участвуют в синтезе белка.
• Субъединицы рибосом собираются в ядре из р. РНК, которая образуется в ядрышке, и белков, которые синтезируются в цитоплазме и поступают в ядро. Затем субъединицы рибосом через ядерные поры перемещаются в цитоплазму, где они участвуют в синтезе белка.
 • Рибосомы могут встречаться в цитоплазме как отдельные гранулы (функционально неактивные, не транслирующие рибосомы), так и в форме скоплений – полирибосом (полисом) – активные рибосомы. Отдельные рибосомы полисом удерживаются вместе нитью информационной РНК.
• Рибосомы могут встречаться в цитоплазме как отдельные гранулы (функционально неактивные, не транслирующие рибосомы), так и в форме скоплений – полирибосом (полисом) – активные рибосомы. Отдельные рибосомы полисом удерживаются вместе нитью информационной РНК.
 • ЭПС – система уплощенных, трубчатых, везикулярных структур, ограниченных мембраной. Название") ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ (ЭПС) • ЭПС – система уплощенных, трубчатых, везикулярных структур, ограниченных мембраной. Название обусловлено тем, что её многочисленные элементы (цистерны, трубочки, пузырьки) образуют единую, непрерывную трехмерную сеть. • Степень развития ЭПС варьирует в различных клетках, и даже в разных участках одной и той же клетки, и зависит от функциональной активности клеток. Различают две разновидности ЭПС: • гранулярную ЭПС (гр. ЭПС) и • гладкую, или агранулярную ЭПС (а. ЭПС), которые связаны между собой в переходной области.
ЭНДОПЛАЗМАТИЧЕСКАЯ СЕТЬ (ЭПС) • ЭПС – система уплощенных, трубчатых, везикулярных структур, ограниченных мембраной. Название обусловлено тем, что её многочисленные элементы (цистерны, трубочки, пузырьки) образуют единую, непрерывную трехмерную сеть. • Степень развития ЭПС варьирует в различных клетках, и даже в разных участках одной и той же клетки, и зависит от функциональной активности клеток. Различают две разновидности ЭПС: • гранулярную ЭПС (гр. ЭПС) и • гладкую, или агранулярную ЭПС (а. ЭПС), которые связаны между собой в переходной области.
 • Гранулярная ЭПС образована мембранными трубочками и уплощенными цистернами, на наружной (обращенной в сторону гиалоплазмы) поверхности которых расположены рибосомы.
• Гранулярная ЭПС образована мембранными трубочками и уплощенными цистернами, на наружной (обращенной в сторону гиалоплазмы) поверхности которых расположены рибосомы.
 вновь синтезированных белковых молекул от") гр. ЭПС • Основная функция гр. ЭПС: сегрегация (отделение) вновь синтезированных белковых молекул от гиалоплазмы. • Таким образом, гр. ЭПС обеспечивает: • биосинтез белков, предназначенных для экспорта из клетки; • биосинтез ферментов лизосом • биосинтез мембранных белков. • Белковые молекулы накапливаются внутри просвета цистерн, приобретают вторичную и третичную структуру, а также подвергаются начальным посттрансляционным изменениям – гидроксилированию, сульфатированию, фосфорилированию и гликозилированию (присоединение к белкам олигосахаридов с образованием гликопротеинов).
гр. ЭПС • Основная функция гр. ЭПС: сегрегация (отделение) вновь синтезированных белковых молекул от гиалоплазмы. • Таким образом, гр. ЭПС обеспечивает: • биосинтез белков, предназначенных для экспорта из клетки; • биосинтез ферментов лизосом • биосинтез мембранных белков. • Белковые молекулы накапливаются внутри просвета цистерн, приобретают вторичную и третичную структуру, а также подвергаются начальным посттрансляционным изменениям – гидроксилированию, сульфатированию, фосфорилированию и гликозилированию (присоединение к белкам олигосахаридов с образованием гликопротеинов).
 Гранулярная эндоплазматическая сеть
Гранулярная эндоплазматическая сеть
 Гранулярная эндоплазматическая сеть
Гранулярная эндоплазматическая сеть
 Гранулярная эндоплазматическая сеть
Гранулярная эндоплазматическая сеть
 • Гр. ЭПС присутствует во всех клетках, но наиболее развита в клетках, специализирующихся на белковом синтезе: • в эпителиальных клетках поджелудочной железы, вырабатывающих пищеварительные ферменты; • в фибробластах соединительной ткани, синтезирующих коллаген; • в плазматических клетках, продуцирующих иммуноглобулины. • В этих клетках элементы гр. ЭПС образуют параллельные скопления цистерн; при этом просвет цистерн часто расширен. • Для всех этих клеток характерна выраженная базофилия цитоплазмы в области расположения элементов гр. ЭПС.
• Гр. ЭПС присутствует во всех клетках, но наиболее развита в клетках, специализирующихся на белковом синтезе: • в эпителиальных клетках поджелудочной железы, вырабатывающих пищеварительные ферменты; • в фибробластах соединительной ткани, синтезирующих коллаген; • в плазматических клетках, продуцирующих иммуноглобулины. • В этих клетках элементы гр. ЭПС образуют параллельные скопления цистерн; при этом просвет цистерн часто расширен. • Для всех этих клеток характерна выраженная базофилия цитоплазмы в области расположения элементов гр. ЭПС.
 Агранулярная ЭПС • агр. ЭПС представляет собой трехмерную сеть мембранных трубочек, канальцев, пузырьков, на поверхности которых рибосомы отсутствуют.
Агранулярная ЭПС • агр. ЭПС представляет собой трехмерную сеть мембранных трубочек, канальцев, пузырьков, на поверхности которых рибосомы отсутствуют.
 Агранулярная эндоплазматическая сеть
Агранулярная эндоплазматическая сеть
 Агранулярная эндоплазматическая сеть
Агранулярная эндоплазматическая сеть
 Агранулярная эндоплазматическая сеть + включения гликогена
Агранулярная эндоплазматическая сеть + включения гликогена
 Функции а. ЭПС • участие в синтезе липидов, в том числе мембранных, холестерина и стероидов; • метаболизм гликогена • нейтрализация и детоксикация эндогенных и экзогенных токсичных веществ; • накопление ионов Са (особенно в специализированной а. ЭПС – саркоплазматической сети мышечных клеток).
Функции а. ЭПС • участие в синтезе липидов, в том числе мембранных, холестерина и стероидов; • метаболизм гликогена • нейтрализация и детоксикация эндогенных и экзогенных токсичных веществ; • накопление ионов Са (особенно в специализированной а. ЭПС – саркоплазматической сети мышечных клеток).
 • Агр. ЭПС хорошо развита: • в клетках, активно продуцирующих стероидные гормоны – клетки коркового вещества надпочечников, интерстициальные гландулоциты яичка, клетки желтого тела яичника. • в клетках печени, где её ферменты участвуют в метаболизме гликогена, а также в процессах, которые обеспечивают нейтрализацию и детоксикацию эндогенных биологически активных веществ (гормонов) и экзогенных вредных веществ (алкоголя, лекарственных веществ и др. )
• Агр. ЭПС хорошо развита: • в клетках, активно продуцирующих стероидные гормоны – клетки коркового вещества надпочечников, интерстициальные гландулоциты яичка, клетки желтого тела яичника. • в клетках печени, где её ферменты участвуют в метаболизме гликогена, а также в процессах, которые обеспечивают нейтрализацию и детоксикацию эндогенных биологически активных веществ (гормонов) и экзогенных вредных веществ (алкоголя, лекарственных веществ и др. )
 КОМПЛЕКС ГОЛЬДЖИ • Комплекс Гольджи – мембранная органелла, образованная тремя основными элементами: • скопления уплощенных цистерн; • мелкие пузырьки; • конденсирующие вакуоли. • Комплекс этих элементов называется диктиосомой.
КОМПЛЕКС ГОЛЬДЖИ • Комплекс Гольджи – мембранная органелла, образованная тремя основными элементами: • скопления уплощенных цистерн; • мелкие пузырьки; • конденсирующие вакуоли. • Комплекс этих элементов называется диктиосомой.
 Комплекс Гольджи • Цистерны имеют вид изогнутых дисков с несколько расширенными периферическими отделами. Цистерны образуют группу в виде стопки из 3 -30 элементов. Выпуклая сторона этой группы обращена обычно к ядру, вогнутая – к плазмолемме. От периферических расширений цистерн отщепляются пузырьки и вакуоли. • Пузырьки – мелкие (диаметр 40 -80 нм), окруженные мембраной сферические элементы с содержимым умеренной электронной плотности. • Вакуоли – крупные (диаметр 0. 1 -1. 0 мкм), сферические образования, отделяющиеся от зрелой поверхности комплекса Гольджи в некоторых железистых клетках. Вакуоли содержат секреторный продукт, находящийся в процессе конденсации.
Комплекс Гольджи • Цистерны имеют вид изогнутых дисков с несколько расширенными периферическими отделами. Цистерны образуют группу в виде стопки из 3 -30 элементов. Выпуклая сторона этой группы обращена обычно к ядру, вогнутая – к плазмолемме. От периферических расширений цистерн отщепляются пузырьки и вакуоли. • Пузырьки – мелкие (диаметр 40 -80 нм), окруженные мембраной сферические элементы с содержимым умеренной электронной плотности. • Вакуоли – крупные (диаметр 0. 1 -1. 0 мкм), сферические образования, отделяющиеся от зрелой поверхности комплекса Гольджи в некоторых железистых клетках. Вакуоли содержат секреторный продукт, находящийся в процессе конденсации.
 • Комплекс Гольджи обладает полярностью: • в каждой диктиосоме выделяют две поверхности: • формирующаяся (незрелая, или цисповерхность) и • зрелая (трансповерхность).
• Комплекс Гольджи обладает полярностью: • в каждой диктиосоме выделяют две поверхности: • формирующаяся (незрелая, или цисповерхность) и • зрелая (трансповерхность).
 Комплекс Гольджи • Цис-поверхность выпуклой формы обращена в сторону ЭПС и связана с ней системой мелких транспортных пузырьков, отщепляющих от ЭПС. • Таким образом, белки в транспортных пузырьках проникают через цисповерхность.
Комплекс Гольджи • Цис-поверхность выпуклой формы обращена в сторону ЭПС и связана с ней системой мелких транспортных пузырьков, отщепляющих от ЭПС. • Таким образом, белки в транспортных пузырьках проникают через цисповерхность.
; • обработка белковых молекул") Функции комплекса Гольджи • синтез полисахаридов и гликопротеинов (гликокаликса, слизи); • обработка белковых молекул (терминальное гликозилирование – включение углеводных компонентов; фосфорилирование – добавление фосфатных групп; ацилирование – добавление жирных кислот; сульфатирование – добавление сульфатных остатков и т. д. ; • сортировка белков на трансповерхности; • упаковка секреторных продуктов в мембранные структуры
Функции комплекса Гольджи • синтез полисахаридов и гликопротеинов (гликокаликса, слизи); • обработка белковых молекул (терминальное гликозилирование – включение углеводных компонентов; фосфорилирование – добавление фосфатных групп; ацилирование – добавление жирных кислот; сульфатирование – добавление сульфатных остатков и т. д. ; • сортировка белков на трансповерхности; • упаковка секреторных продуктов в мембранные структуры
 Комплекс Гольджи
Комплекс Гольджи
 Комплекс Гольджи
Комплекс Гольджи
 МИТОХОНДРИИ • Митохондрии – мембранные органеллы, присутствующих во всех эукариотических клетках, и представляющие собой энергетический аппарат клетки.
МИТОХОНДРИИ • Митохондрии – мембранные органеллы, присутствующих во всех эукариотических клетках, и представляющие собой энергетический аппарат клетки.
 Функции митохондрий: • основная – обеспечение клетки легко доступной энергией; • участие в биосинтезе стероидов; • участие в окислении жирных кислот.
Функции митохондрий: • основная – обеспечение клетки легко доступной энергией; • участие в биосинтезе стероидов; • участие в окислении жирных кислот.
 • Митохондрии могут иметь эллиптическую, палочковидную или нитевидную форму. Их размеры составляют 0. 2 -2 мкм в ширину и до 10 мкм в длину. • Число митохондрий в разных клетках и их распределение в пределах клетки варьирует.
• Митохондрии могут иметь эллиптическую, палочковидную или нитевидную форму. Их размеры составляют 0. 2 -2 мкм в ширину и до 10 мкм в длину. • Число митохондрий в разных клетках и их распределение в пределах клетки варьирует.
 • Под электронным микроскопом митохондрии имеют характерную структуру. • Каждая митохондрия состоит из наружной и внутренней мембран, между которыми находится межмембранное пространство. • Внутренняя мембрана образует складки - кристы, обращенные внутрь митохондрии. На кристах имеются элементарные частицы (оксисомы, или F 1 -частицы), на которых происходит сопряжение процессов окисления и фосфорилирования (АДФ → АТФ). Пространство, ограниченное внутренней мембраной, заполнено митохондриальным матриксом, мелкозернистым материалом различной электронной плотности.
• Под электронным микроскопом митохондрии имеют характерную структуру. • Каждая митохондрия состоит из наружной и внутренней мембран, между которыми находится межмембранное пространство. • Внутренняя мембрана образует складки - кристы, обращенные внутрь митохондрии. На кристах имеются элементарные частицы (оксисомы, или F 1 -частицы), на которых происходит сопряжение процессов окисления и фосфорилирования (АДФ → АТФ). Пространство, ограниченное внутренней мембраной, заполнено митохондриальным матриксом, мелкозернистым материалом различной электронной плотности.
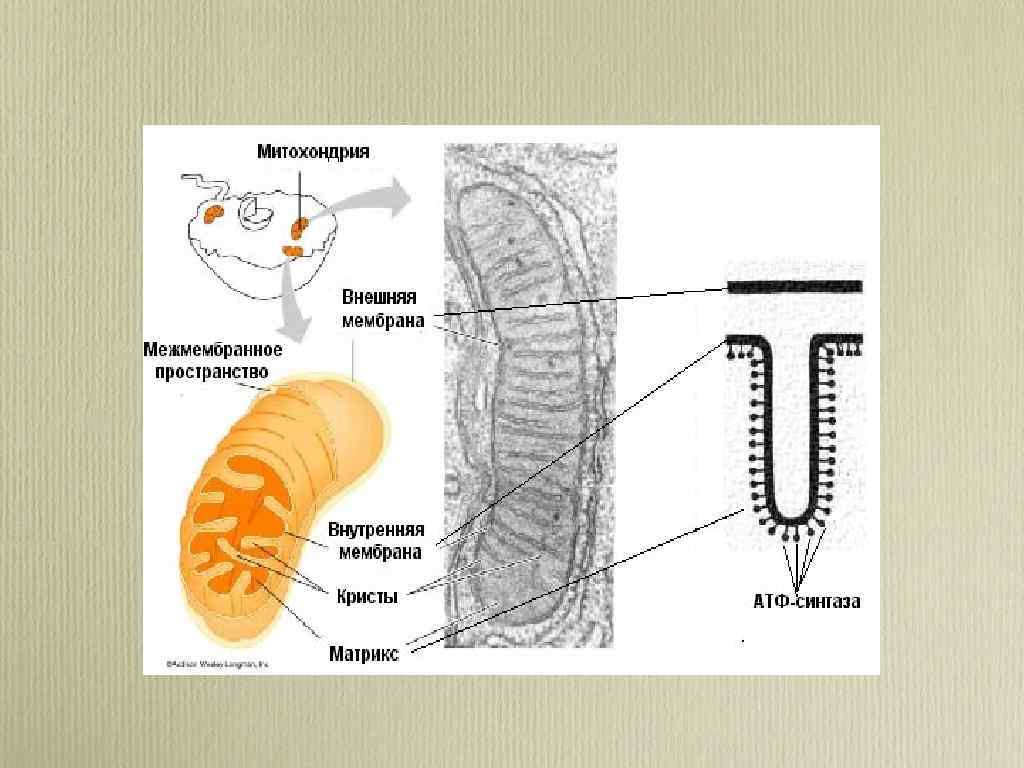
") • Чаще всего кристы располагаются перпендикулярно длинной оси митохондрий и имеют пластинчатую (ламеллярную) форму.
• Чаще всего кристы располагаются перпендикулярно длинной оси митохондрий и имеют пластинчатую (ламеллярную) форму.
 • В клетках, синтезирующих стероидные гормоны, кристы имеют вид трубочек или пузырьков - тубулярновезикулярные кристы. • В этих клетках ферменты стероидного синтеза частично локализуются на внутренней мембране митохондрий
• В клетках, синтезирующих стероидные гормоны, кристы имеют вид трубочек или пузырьков - тубулярновезикулярные кристы. • В этих клетках ферменты стероидного синтеза частично локализуются на внутренней мембране митохондрий
 • Число и площадь крист отражает функциональную активность клеток: наибольшая площадь крист характерна, например, для митохондрий клеток сердечной мышцы, где потребность в энергии постоянно очень велика.
• Число и площадь крист отражает функциональную активность клеток: наибольшая площадь крист характерна, например, для митохондрий клеток сердечной мышцы, где потребность в энергии постоянно очень велика.
 • Митохондриальный матрикс – мелкозернистое вещество, заполняющее полость митохондрии, содержит несколько сотен ферментов: ферменты цикла Кребса, окисления жирных кислот, белкового синтеза. • Здесь иногда встречаются митохондриальные гранулы, а также локализуются митохондриальные ДНК, и. РНК, т. РНК, р. РНК и митохондриальные рибосомы.
• Митохондриальный матрикс – мелкозернистое вещество, заполняющее полость митохондрии, содержит несколько сотен ферментов: ферменты цикла Кребса, окисления жирных кислот, белкового синтеза. • Здесь иногда встречаются митохондриальные гранулы, а также локализуются митохондриальные ДНК, и. РНК, т. РНК, р. РНК и митохондриальные рибосомы.
 • Митохондриальные гранулы – частицы высокой электронной плотности диаметром 20 -50 нм, содержащие ионы Са и Мg.
• Митохондриальные гранулы – частицы высокой электронной плотности диаметром 20 -50 нм, содержащие ионы Са и Мg.
 Митохондрия
Митохондрия
 Митохондрии
Митохондрии
 Митохондрии
Митохондрии
 Митохондрии с тубуло-везикулярными кристами
Митохондрии с тубуло-везикулярными кристами
 макромолекул внеклеточного и") ЛИЗОСОМЫ • Лизосомы – мембранные органеллы, которые обеспечивают внутриклеточное переваривание (расщепление) макромолекул внеклеточного и внутриклеточного происхождения, и обновление компонентов клетки.
ЛИЗОСОМЫ • Лизосомы – мембранные органеллы, которые обеспечивают внутриклеточное переваривание (расщепление) макромолекул внеклеточного и внутриклеточного происхождения, и обновление компонентов клетки.
 • Морфологически лизосомы представляют собой округлые пузырьки, ограниченные мембраной и содержащие большое количество различных гидролаз (более 60 ферментов).
• Морфологически лизосомы представляют собой округлые пузырьки, ограниченные мембраной и содержащие большое количество различных гидролаз (более 60 ферментов).
, протеазы, нуклеазы, сульфатазы,") • Наиболее характерными ферментами лизосом являются: кислая фосфатаза (маркёр лизосом), протеазы, нуклеазы, сульфатазы, липазы, гликозидазы. • Все литические ферменты лизосом представляют собой кислые гидролазы, т. е. оптимум их активности проявляется при р. Н≈5 • Повреждение мембраны приводит к разрушению клетки вследствие самопереваривания.
• Наиболее характерными ферментами лизосом являются: кислая фосфатаза (маркёр лизосом), протеазы, нуклеазы, сульфатазы, липазы, гликозидазы. • Все литические ферменты лизосом представляют собой кислые гидролазы, т. е. оптимум их активности проявляется при р. Н≈5 • Повреждение мембраны приводит к разрушению клетки вследствие самопереваривания.

 и вторичные (активные). • Первичные лизосомы") Первичные лизосомы • Лизосомы подразделяются на первичные (неактивные) и вторичные (активные). • Первичные лизосомы (гидролазные пузырьки) – неактивные структуры, еще не вступившие в процессы расщепления субстратов. Округлые пузырьки небольшого размера c гомогенным, плотным матриксом.
Первичные лизосомы • Лизосомы подразделяются на первичные (неактивные) и вторичные (активные). • Первичные лизосомы (гидролазные пузырьки) – неактивные структуры, еще не вступившие в процессы расщепления субстратов. Округлые пузырьки небольшого размера c гомогенным, плотным матриксом.
 • Вторичные лизосомы – органеллы, активно участвующие в процессах внутриклеточного переваривания, – результат слияния первичной лизосомы с фагосомой или аутофагосомой. Обычно содержимое вторичных лизосом гетерогенно.
• Вторичные лизосомы – органеллы, активно участвующие в процессах внутриклеточного переваривания, – результат слияния первичной лизосомы с фагосомой или аутофагосомой. Обычно содержимое вторичных лизосом гетерогенно.
 Фаголизосомы • Фаголизосома формируется путем слияния первичной лизосомы с фагосомой - мембранным пузырьком, содержащим материал, захваченный клеткой извне. Процесс разрушения этого материала называется гетерофагией.
Фаголизосомы • Фаголизосома формируется путем слияния первичной лизосомы с фагосомой - мембранным пузырьком, содержащим материал, захваченный клеткой извне. Процесс разрушения этого материала называется гетерофагией.
 • Аутофаголизосома образуется при слиянии первичной лизосомы с аутофагосомой мембранным пузырьком, содержащим собственные компоненты клетки, которые подлежат разрушению. • Процесс переваривания внутриклеточного материала называется аутофагией.
• Аутофаголизосома образуется при слиянии первичной лизосомы с аутофагосомой мембранным пузырьком, содержащим собственные компоненты клетки, которые подлежат разрушению. • Процесс переваривания внутриклеточного материала называется аутофагией.
 • Остаточные тельца – лизосомы, содержащие непереваренный материал, которые могут находиться в цитоплазме длительное время. • В некоторых долгоживущих клетках (нейроны, кардиомиоциты, гепатоциты) в остаточных тельцах накапливается коричневый эндогенный пигмент липофусцин – «пигмент старения» .
• Остаточные тельца – лизосомы, содержащие непереваренный материал, которые могут находиться в цитоплазме длительное время. • В некоторых долгоживущих клетках (нейроны, кардиомиоциты, гепатоциты) в остаточных тельцах накапливается коричневый эндогенный пигмент липофусцин – «пигмент старения» .
 ПЕРОКСИСОМЫ • Пероксисомы – сферические мембранные органеллы диаметром 0. 05 – 1. 5 мкм, с умеренно плотным гомогенным или мелкозернистым матриксом. • Матрикс пероксисом содержит до 50 различных ферментов, важнейшие из которых: каталаза (маркёр пероксисом), пероксидаза, оксидазы аминокислот, уратоксидаза.
ПЕРОКСИСОМЫ • Пероксисомы – сферические мембранные органеллы диаметром 0. 05 – 1. 5 мкм, с умеренно плотным гомогенным или мелкозернистым матриксом. • Матрикс пероксисом содержит до 50 различных ферментов, важнейшие из которых: каталаза (маркёр пероксисом), пероксидаза, оксидазы аминокислот, уратоксидаза.
 Функции пероксисом: • окисление аминокислот и других субстратов; • защита клетки от действия перекиси водорода, сильного окислителя, образующегося в результате окисления органических соединений, и оказывающего повреждающий эффект на клетку. При этом каталаза пероксисом разлагает перекись водорода на воду и кислород. • участие в расщеплении жирных кислот; • участие в обезвреживании ряда веществ (спирт и др. ).
Функции пероксисом: • окисление аминокислот и других субстратов; • защита клетки от действия перекиси водорода, сильного окислителя, образующегося в результате окисления органических соединений, и оказывающего повреждающий эффект на клетку. При этом каталаза пероксисом разлагает перекись водорода на воду и кислород. • участие в расщеплении жирных кислот; • участие в обезвреживании ряда веществ (спирт и др. ).
 Лизосомы и пероксисомы
Лизосомы и пероксисомы
 ЦИТОСКЕЛЕТ • Цитоскелет – сложная трехмерная сеть немембранных органелл: • микротрубочек • микрофиламентов • промежуточных филаментов.
ЦИТОСКЕЛЕТ • Цитоскелет – сложная трехмерная сеть немембранных органелл: • микротрубочек • микрофиламентов • промежуточных филаментов.
 • Основная функция цитоскелета – опорнодвигательная: • поддержание и изменение формы клеток; • перемещение компонентов внутри клетки; • транспорт веществ внутрь клетки и из клетки; • обеспечение подвижности клетки
• Основная функция цитоскелета – опорнодвигательная: • поддержание и изменение формы клеток; • перемещение компонентов внутри клетки; • транспорт веществ внутрь клетки и из клетки; • обеспечение подвижности клетки
 Микротрубочки • Микротрубочки – полые цилиндрические образования различной длины, наиболее крупные компоненты цитоскелета.
Микротрубочки • Микротрубочки – полые цилиндрические образования различной длины, наиболее крупные компоненты цитоскелета.
 Функции микротрубочек – поддержание стабильной формы клеток, и порядка распределения её компонентов; – обеспечение внутриклеточного транспорта, в том числе органелл, пузырьков, секреторных гранул (благодаря некоторым белкам, ассоциированным с микротрубочками); – образование основы центриолей и ахроматинового веретена деления и обеспечение движения хромосом в процессе митоза; – образование основы ресничек и жгутиков, а также обеспечение их движения.
Функции микротрубочек – поддержание стабильной формы клеток, и порядка распределения её компонентов; – обеспечение внутриклеточного транспорта, в том числе органелл, пузырьков, секреторных гранул (благодаря некоторым белкам, ассоциированным с микротрубочками); – образование основы центриолей и ахроматинового веретена деления и обеспечение движения хромосом в процессе митоза; – образование основы ресничек и жгутиков, а также обеспечение их движения.
 • Стенка микротрубочки состоит из спирально расположенных нитей – протофиламентов, образованных из молекул белкα-тубулина.
• Стенка микротрубочки состоит из спирально расположенных нитей – протофиламентов, образованных из молекул белкα-тубулина.
 • Микротрубочки могут располагаться в цитоплазме в виде отдельных элементов, в виде пучков, где они связаны тонкими поперечными мостиками, или могут частично сливаться друг с другом, образуя дуплеты (в аксонеме ресничек и жгутиков) и триплеты (в базальном тельце и центриолях
• Микротрубочки могут располагаться в цитоплазме в виде отдельных элементов, в виде пучков, где они связаны тонкими поперечными мостиками, или могут частично сливаться друг с другом, образуя дуплеты (в аксонеме ресничек и жгутиков) и триплеты (в базальном тельце и центриолях
 Клеточный центр • Клеточный центр образован двумя полыми цилиндрическими структурами центриолями, которые расположены под прямым углом друг к другу.
Клеточный центр • Клеточный центр образован двумя полыми цилиндрическими структурами центриолями, которые расположены под прямым углом друг к другу.
 • Каждая центриоль представляет собой короткий цилиндр длиной ~ 0, 5 мкм и диаметром ~ 0, 2 мкм, состоящий из 9 триплетов частично слившихся трубочек (А, В и С), связанных поперечными белковыми мостиками. • Формула строения центриоли описывается как (9 × 3) + 0, так как в центральной части микротрубочки отсутствуют. Каждый триплет центриоли связан с глобулярными белковыми тельцами – сателлитами, от которых отходят микротрубочки, образующие центросферу.
• Каждая центриоль представляет собой короткий цилиндр длиной ~ 0, 5 мкм и диаметром ~ 0, 2 мкм, состоящий из 9 триплетов частично слившихся трубочек (А, В и С), связанных поперечными белковыми мостиками. • Формула строения центриоли описывается как (9 × 3) + 0, так как в центральной части микротрубочки отсутствуют. Каждый триплет центриоли связан с глобулярными белковыми тельцами – сателлитами, от которых отходят микротрубочки, образующие центросферу.
 Центриоли • В неделящейся клетке выявляется одна пара центриолей – диплосома, которая располагается обычно вблизи ядра. • Перед делением клетки в S-периоде интерфазы происходит дупликация центриолей: под прямым углом к каждой зрелой (материнской) центриоли пары образуется новая (дочерняя) центриоль. • В ранней профазе митоза пары центриолей расходятся к полюсам клетки и служат центрами образования микротрубочек ахроматинового веретена деления.
Центриоли • В неделящейся клетке выявляется одна пара центриолей – диплосома, которая располагается обычно вблизи ядра. • Перед делением клетки в S-периоде интерфазы происходит дупликация центриолей: под прямым углом к каждой зрелой (материнской) центриоли пары образуется новая (дочерняя) центриоль. • В ранней профазе митоза пары центриолей расходятся к полюсам клетки и служат центрами образования микротрубочек ахроматинового веретена деления.
 Реснички и жгутики • Реснички и жгутики являются выростами цитоплазмы, обладающие подвижностью. Основу ресничек и жгутиков составляет каркас из микротрубочек, называемый аксонемой. • Длина ресничек равна 2 -10 мкм, а их количество на поверхности одной клетки может составлять до нескольких сотен. • В организме человека жгутик есть только в одном типе клеток – сперматозоидах. При этом один сперматозоид имеет один жгутик длиной 50 -70 мкм.
Реснички и жгутики • Реснички и жгутики являются выростами цитоплазмы, обладающие подвижностью. Основу ресничек и жгутиков составляет каркас из микротрубочек, называемый аксонемой. • Длина ресничек равна 2 -10 мкм, а их количество на поверхности одной клетки может составлять до нескольких сотен. • В организме человека жгутик есть только в одном типе клеток – сперматозоидах. При этом один сперматозоид имеет один жгутик длиной 50 -70 мкм.
 и одной") • Аксонема образована 9 периферическими парами микротрубочек (микротрубочки А и В) и одной центрально расположенной парой; • такое строение описывается формулой • (9 × 2) + 2. • Центральная пара микротрубочек окружена центральной оболочкой, от которой к периферическим дуплетам расходятся радиальные спицы. • Периферические дублеты связаны друг с другом мостиками белка нексина, а от микротрубочки А к микротрубочке В соседнего дублета отходят “ручки” из белка динеина, который обладает АТФазной активностью, что необходимо для скольжения соседних дублетов в аксонеме, вызывающих движение (биение) ресничек и жгутиков
• Аксонема образована 9 периферическими парами микротрубочек (микротрубочки А и В) и одной центрально расположенной парой; • такое строение описывается формулой • (9 × 2) + 2. • Центральная пара микротрубочек окружена центральной оболочкой, от которой к периферическим дуплетам расходятся радиальные спицы. • Периферические дублеты связаны друг с другом мостиками белка нексина, а от микротрубочки А к микротрубочке В соседнего дублета отходят “ручки” из белка динеина, который обладает АТФазной активностью, что необходимо для скольжения соседних дублетов в аксонеме, вызывающих движение (биение) ресничек и жгутиков
 • В основании каждой реснички или жгутика лежит базальное тельце, сходное по строению с центриолью.
• В основании каждой реснички или жгутика лежит базальное тельце, сходное по строению с центриолью.
 Микроворсинки • Микроворсинки – пальцевидные выросты цитоплазмы клетки диаметром 0. 1 мкм и длиной 1 мкм, основу которых образуют актиновые микрофиламенты. • Микроворсинки обеспечивают многократное увеличение площади поверхности клетки. На апикальной поверхности некоторых клеток, активно участвующих в процессах расщепления и всасывания веществ, имеется до несколько тысяч микроворсинок, образующих в совокупности щёточную каемку (эпителий тонкой кишки и почечных канальцев).
Микроворсинки • Микроворсинки – пальцевидные выросты цитоплазмы клетки диаметром 0. 1 мкм и длиной 1 мкм, основу которых образуют актиновые микрофиламенты. • Микроворсинки обеспечивают многократное увеличение площади поверхности клетки. На апикальной поверхности некоторых клеток, активно участвующих в процессах расщепления и всасывания веществ, имеется до несколько тысяч микроворсинок, образующих в совокупности щёточную каемку (эпителий тонкой кишки и почечных канальцев).
 Реснички
Реснички
 Реснички и микроворсинки
Реснички и микроворсинки
 Микроворсинки в продольном и поперечном срезе
Микроворсинки в продольном и поперечном срезе
 Микрофиламенты • Микрофиламенты – тонкие белковые нити диаметром 57 нм, расположенные в цитоплазме поодиночке, в виде сетей или упорядоченными пучками (в скелетной и сердечной мышцах). • Основной белок микрофиламентов – актин
Микрофиламенты • Микрофиламенты – тонкие белковые нити диаметром 57 нм, расположенные в цитоплазме поодиночке, в виде сетей или упорядоченными пучками (в скелетной и сердечной мышцах). • Основной белок микрофиламентов – актин
 • Промежуточные филаменты – прочные и устойчивые белковые нити толщиной около 10 нм (что является промежуточным значением между толщиной микротрубочек и микрофиламентов). • Главная функция промежуточных филаментов – поддерживающая и опорная • Выделяются 6 основных классов промежуточных филаментов:
• Промежуточные филаменты – прочные и устойчивые белковые нити толщиной около 10 нм (что является промежуточным значением между толщиной микротрубочек и микрофиламентов). • Главная функция промежуточных филаментов – поддерживающая и опорная • Выделяются 6 основных классов промежуточных филаментов:
 • Цитокератины – промежуточные филаменты, характерные для клеток эпителия (являются главным компонентом волос и ногтей) • Десмины – промежуточные филаменты мышечных тканей (за исключением миоцитов сосудов) • Виментины – филаменты, характерные для различных клеток мезенхимного происхождения (фибробласты, макрофаги, остеобласты, эндотелий и гладкие миоциты сосудов). • Нейрофиламенты – промежуточные филаменты нейронов, которые играют важную роль в поддержании формы отростков нервных клеток. • Глиальные филаменты встречаются только в клетках глии (астроциты, олигодендроциты). • Ламины – промежуточные филаменты ядер различных типов клеток, образующие кариоскелет.
• Цитокератины – промежуточные филаменты, характерные для клеток эпителия (являются главным компонентом волос и ногтей) • Десмины – промежуточные филаменты мышечных тканей (за исключением миоцитов сосудов) • Виментины – филаменты, характерные для различных клеток мезенхимного происхождения (фибробласты, макрофаги, остеобласты, эндотелий и гладкие миоциты сосудов). • Нейрофиламенты – промежуточные филаменты нейронов, которые играют важную роль в поддержании формы отростков нервных клеток. • Глиальные филаменты встречаются только в клетках глии (астроциты, олигодендроциты). • Ламины – промежуточные филаменты ядер различных типов клеток, образующие кариоскелет.
 • Цитокератины – промежуточные филаменты, характерные для клеток эпителия
• Цитокератины – промежуточные филаменты, характерные для клеток эпителия
.") • Десмины – промежуточные филаменты мышечных тканей (за исключением миоцитов сосудов).
• Десмины – промежуточные филаменты мышечных тканей (за исключением миоцитов сосудов).
 • Виментины – филаменты, характерные для различных клеток мезенхимного происхождения (фибробласты, макрофаги, остеобласты, эндотелий и гладкие миоциты сосудов).
• Виментины – филаменты, характерные для различных клеток мезенхимного происхождения (фибробласты, макрофаги, остеобласты, эндотелий и гладкие миоциты сосудов).
 • Нейрофиламенты – промежуточные филаменты нейронов • Глиальные клетки содержат глиальный фибриллярный кислый белок и встречаются только в клетках глии (астроциты, олигодендроциты).
• Нейрофиламенты – промежуточные филаменты нейронов • Глиальные клетки содержат глиальный фибриллярный кислый белок и встречаются только в клетках глии (астроциты, олигодендроциты).
 ЦИТОЛОГИЯ ЯДРО ДЕЛЕНИЕ КЛЕТОК
ЦИТОЛОГИЯ ЯДРО ДЕЛЕНИЕ КЛЕТОК
 Ядро интерфазной клетки
Ядро интерфазной клетки
 Ядро – важнейший компонент клетки, содержащий её генетический аппарат. Компоненты ядра n n n В ядре неделящейся (интерфазной) клетки выявляются следующие компоненты: ядерная оболочка (кариолемма) хроматин ядрышко кариоплазма
Ядро – важнейший компонент клетки, содержащий её генетический аппарат. Компоненты ядра n n n В ядре неделящейся (интерфазной) клетки выявляются следующие компоненты: ядерная оболочка (кариолемма) хроматин ядрышко кариоплазма
 на светооптическом уровне практически не") Ядерная оболочка n n n Ядерная оболочка (кариолемма, нуклеолемма) на светооптическом уровне практически не определяется. Под электронном микроскопом обнаруживается, что она состоит из двух мембран – наружной и внутренней мембран, разделенных полостью шириной 15 -40 нм – перинуклеарной цистерной. Наружная мембрана составляет единое целое с мембранами гр. ЭПС: на её поверхности имеются рибосомы, а перинуклеарная цистерна сообщается с цистерной гр. ЭПС
Ядерная оболочка n n n Ядерная оболочка (кариолемма, нуклеолемма) на светооптическом уровне практически не определяется. Под электронном микроскопом обнаруживается, что она состоит из двух мембран – наружной и внутренней мембран, разделенных полостью шириной 15 -40 нм – перинуклеарной цистерной. Наружная мембрана составляет единое целое с мембранами гр. ЭПС: на её поверхности имеются рибосомы, а перинуклеарная цистерна сообщается с цистерной гр. ЭПС
; n") n Функции ядра: n хранение генетической информации (в молекулах ДНК, находящихся в хромосомах); n реализация генетической информации, контролирующей различные процессы в клетке: транскрипция информационных, рибосомальных, транспортных РНК → синтетическая активность; апоптоз и т. д. ); n воспроизведение и передача генетической информации при делении клетки.
n Функции ядра: n хранение генетической информации (в молекулах ДНК, находящихся в хромосомах); n реализация генетической информации, контролирующей различные процессы в клетке: транскрипция информационных, рибосомальных, транспортных РНК → синтетическая активность; апоптоз и т. д. ); n воспроизведение и передача генетической информации при делении клетки.

 n n n В определенных точках наружная и внутренняя мембрана смыкаются, образуя ядерные поры. Ядерная пора образована двумя параллельными кольцами диаметром 80 нм, содержащих по 8 белковых гранул, от которых к центру поры тянутся фибриллы, формирующие диафрагму толщиной около 5 нм. В середине диафрагмы лежит центральная гранула. Белковые гранулы ядерной поры структурно связаны с белками ядерной ламины. Совокупность компонентов, входящих в состав ядерной поры, называется комплексом ядерной поры.
n n n В определенных точках наружная и внутренняя мембрана смыкаются, образуя ядерные поры. Ядерная пора образована двумя параллельными кольцами диаметром 80 нм, содержащих по 8 белковых гранул, от которых к центру поры тянутся фибриллы, формирующие диафрагму толщиной около 5 нм. В середине диафрагмы лежит центральная гранула. Белковые гранулы ядерной поры структурно связаны с белками ядерной ламины. Совокупность компонентов, входящих в состав ядерной поры, называется комплексом ядерной поры.
 n Ядерная оболочка клетки содержит 2000 -4000 поровых комплексов. n Число поровых комплексов возрастает с увеличением функциональной активности: в клетках с высокой синтетической активностью ядерные поры занимают до 35% поверхности кариолеммы.
n Ядерная оболочка клетки содержит 2000 -4000 поровых комплексов. n Число поровых комплексов возрастает с увеличением функциональной активности: в клетках с высокой синтетической активностью ядерные поры занимают до 35% поверхности кариолеммы.
 Комплекс ядерной поры n По каналу, образованному поровым комплексом, движутся мелкие водорастворимые молекулы и ионы; в ядро активно переносятся белки, синтезируемые в цитоплазме ; из ядра в цитоплазму переносятся субъединицы рибосом.
Комплекс ядерной поры n По каналу, образованному поровым комплексом, движутся мелкие водорастворимые молекулы и ионы; в ядро активно переносятся белки, синтезируемые в цитоплазме ; из ядра в цитоплазму переносятся субъединицы рибосом.
 клетке соответствует хромосомам и состоит из комплекса") n n n Хроматин в интерфазной (неделящейся) клетке соответствует хромосомам и состоит из комплекса ДНК и белка. Выраженность спирализации каждой из хромосом неодинакова по длине. Соответственно, различают два вида хроматина: эухроматин и гетерохроматин. Эухроматин соответствует участкам хромосом, которые деспирализованы и открыты для транскрипции. Эти участки не окрашиваются и не видны в световой микроскоп. Гетерохроматин соответствует конденсированным сегментам хромосом, что делает их недоступными для транскрипции. Гетерохроматин интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид мелких гранул и глыбок.
n n n Хроматин в интерфазной (неделящейся) клетке соответствует хромосомам и состоит из комплекса ДНК и белка. Выраженность спирализации каждой из хромосом неодинакова по длине. Соответственно, различают два вида хроматина: эухроматин и гетерохроматин. Эухроматин соответствует участкам хромосом, которые деспирализованы и открыты для транскрипции. Эти участки не окрашиваются и не видны в световой микроскоп. Гетерохроматин соответствует конденсированным сегментам хромосом, что делает их недоступными для транскрипции. Гетерохроматин интенсивно окрашивается основными красителями, и в световом микроскопе имеет вид мелких гранул и глыбок.
 Структура ядер
Структура ядер
 эухроматин-гетерохроматин n по соотношению эу- и гетерохроматина в ядре можно оценить активность синтетической функции клетки. n При её повышении это соотношение изменяется в пользу эухроматина, при снижении – нарастает содержание гетерохроматина.
эухроматин-гетерохроматин n по соотношению эу- и гетерохроматина в ядре можно оценить активность синтетической функции клетки. n При её повышении это соотношение изменяется в пользу эухроматина, при снижении – нарастает содержание гетерохроматина.
 – скопление гетерохроматина, соответствующее одной из пары Х-хромосом, которая") n Половой хроматин (тельце Барра) – скопление гетерохроматина, соответствующее одной из пары Х-хромосом, которая в интерфазе плотно скручена и неактивна. n Выявление полового хроматина используется как диагностический тест для определения генетического женского пола, что существенно при изучении генетических аномалий и, особенно, в спортивной медицине. n Обычно анализируют эпителиальные клетки слизистой оболочки полости рта, где, как и в большинстве других клеток, половой хроматин выявляется как крупная глыбка гетерохроматина, лежащая рядом с ядерной оболочки. n В нейтрофильных лейкоцитах крови половой хроматин имеет вид маленькой добавочной дольки ядра ( «барабанной палочки» ).
n Половой хроматин (тельце Барра) – скопление гетерохроматина, соответствующее одной из пары Х-хромосом, которая в интерфазе плотно скручена и неактивна. n Выявление полового хроматина используется как диагностический тест для определения генетического женского пола, что существенно при изучении генетических аномалий и, особенно, в спортивной медицине. n Обычно анализируют эпителиальные клетки слизистой оболочки полости рта, где, как и в большинстве других клеток, половой хроматин выявляется как крупная глыбка гетерохроматина, лежащая рядом с ядерной оболочки. n В нейтрофильных лейкоцитах крови половой хроматин имеет вид маленькой добавочной дольки ядра ( «барабанной палочки» ).
 Ядрышко n Ядрышко выявляется в интерфазном ядре на светооптическом уровне как мелкая (~ 1 мкм в диаметре), плотная сферическая структура, интенсивно окрашивающееся основными красителями
Ядрышко n Ядрышко выявляется в интерфазном ядре на светооптическом уровне как мелкая (~ 1 мкм в диаметре), плотная сферическая структура, интенсивно окрашивающееся основными красителями
 Ядрышко n Ядрышко образовано специализированными участками хромосом – ядрышковыми организаторами, на которых происходит синтез рибосомальной РНК, а также её сборка в предшественники рибосомальных субъединиц.
Ядрышко n Ядрышко образовано специализированными участками хромосом – ядрышковыми организаторами, на которых происходит синтез рибосомальной РНК, а также её сборка в предшественники рибосомальных субъединиц.
 Ядрышко n Размеры и объем ядрышек увеличиваются при повышении функциональной активности клетки. Особенно крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также клеток быстрорастущих злокачественных опухолей. n Ядрышко исчезает в профазе митоза, в результате инактивации рибосомных
Ядрышко n Размеры и объем ядрышек увеличиваются при повышении функциональной активности клетки. Особенно крупные ядрышки характерны для эмбриональных и активно синтезирующих белки клеток, а также клеток быстрорастущих злокачественных опухолей. n Ядрышко исчезает в профазе митоза, в результате инактивации рибосомных
 Ядерный матрикс n n n Ядерный матрикс – компонент ядра, в котором располагаются хроматин и ядрышко. Ядерный матрикс образован кариоплазмой и кариоскелетом. Кариоплазма – жидкий компонент ядра, содержащий РНК, ионы, ферменты, метаболиты, растворенные в воде. Кариоскелет состоит из ламины и других фибриллярных белков.
Ядерный матрикс n n n Ядерный матрикс – компонент ядра, в котором располагаются хроматин и ядрышко. Ядерный матрикс образован кариоплазмой и кариоскелетом. Кариоплазма – жидкий компонент ядра, содержащий РНК, ионы, ферменты, метаболиты, растворенные в воде. Кариоскелет состоит из ламины и других фибриллярных белков.
 Интерфазное ядро
Интерфазное ядро
 Сегментированное ядро
Сегментированное ядро
 Ядро
Ядро
 ЯДРО
ЯДРО
 Ядрышки
Ядрышки
 Ядрышко n - Два компонента ядрышка: гранулярный фибриллярный
Ядрышко n - Два компонента ядрышка: гранулярный фибриллярный
 Ядерные поры
Ядерные поры
 Ядерные поры
Ядерные поры
") Ядерная пора (схема строения)
Ядерная пора (схема строения)
 КЛЕТОЧНЫЙ ЦИКЛ n n Клеточный цикл – совокупность процессов, происходящих в клетке между двумя последовательными делениями или между её образованием и гибелью. Клеточный цикл включает в себя собственно митотическое деление и интерфазу – промежуток между делениями.
КЛЕТОЧНЫЙ ЦИКЛ n n Клеточный цикл – совокупность процессов, происходящих в клетке между двумя последовательными делениями или между её образованием и гибелью. Клеточный цикл включает в себя собственно митотическое деление и интерфазу – промежуток между делениями.
 ИНТЕРФАЗА n Интерфаза занимает около 90% всего времени клеточного цикла и подразделяется на три периода: n пресинтетический или постмитотический – G 1 (от англ. gap – промежуток); n синтетический – S; n постсинтетический или премитотический - G 2.
ИНТЕРФАЗА n Интерфаза занимает около 90% всего времени клеточного цикла и подразделяется на три периода: n пресинтетический или постмитотический – G 1 (от англ. gap – промежуток); n синтетический – S; n постсинтетический или премитотический - G 2.
 Пресинтетический период – G 1 n n Пресинтетический период – G 1 – характеризуется активным ростом клетки, синтезом белка и РНК, благодаря чему клетка восстанавливает необходимый набор органелл и достигает нормальных размеров. G 1 период длится от нескольких часов до нескольких дней. В течение этого периода синтезируются особые «запускающие» белки – активаторы S периода. Они обеспечивают достижение клеткой точки R (точки ограничения), после которого она вступает в S-период.
Пресинтетический период – G 1 n n Пресинтетический период – G 1 – характеризуется активным ростом клетки, синтезом белка и РНК, благодаря чему клетка восстанавливает необходимый набор органелл и достигает нормальных размеров. G 1 период длится от нескольких часов до нескольких дней. В течение этого периода синтезируются особые «запускающие» белки – активаторы S периода. Они обеспечивают достижение клеткой точки R (точки ограничения), после которого она вступает в S-период.
 Период репродуктивного покоя n n n Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G 0). Клетки некоторых тканей под влиянием определенных факторов способны возвращаться из периода G 0 в клеточный цикл, клетки других тканей утрачивают эту способность по мере дифференцировки. Абсолютное большинство дифференцированных клеток организма, выполняющих свои специфические функции, не делятся.
Период репродуктивного покоя n n n Если клетка не достигает точки R, она выходит из цикла и вступает в период репродуктивного покоя (G 0). Клетки некоторых тканей под влиянием определенных факторов способны возвращаться из периода G 0 в клеточный цикл, клетки других тканей утрачивают эту способность по мере дифференцировки. Абсолютное большинство дифференцированных клеток организма, выполняющих свои специфические функции, не делятся.
 ДНК, синтезом гистонов и других") Синтетический период n Синтетический период –Sхарактеризуется репликацией (удвоением содержания) ДНК, синтезом гистонов и других белков. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. n S-период длится у большинства клеток 812 часов.
Синтетический период n Синтетический период –Sхарактеризуется репликацией (удвоением содержания) ДНК, синтезом гистонов и других белков. В результате происходит удвоение числа хромосом. Одновременно удваивается число центриолей. n S-период длится у большинства клеток 812 часов.
 Постсинтетический период n Постсинтетический период – G 2 - длится 2 -4 часа и продолжается вплоть до митоза. n В течение этого периода запасается энергия, и синтезируются белки, в частности тубулины, необходимые для процесса деления.
Постсинтетический период n Постсинтетический период – G 2 - длится 2 -4 часа и продолжается вплоть до митоза. n В течение этого периода запасается энергия, и синтезируются белки, в частности тубулины, необходимые для процесса деления.
 является универсальным механизмом деления соматических клеток. Во время митоза") Митоз n n Митоз (кариокинез) является универсальным механизмом деления соматических клеток. Во время митоза родительская клетка делится, и каждая из дочерних клеток получает набор хромосом идентичный родительскому, и, таким образом, происходит равномерное распределение генетического материала. Продолжительность митоза – 1 -3 часа.
Митоз n n Митоз (кариокинез) является универсальным механизмом деления соматических клеток. Во время митоза родительская клетка делится, и каждая из дочерних клеток получает набор хромосом идентичный родительскому, и, таким образом, происходит равномерное распределение генетического материала. Продолжительность митоза – 1 -3 часа.
 Митоз n Митоз включает n 4 основные фазы: n профазу n метафазу, n анафазу и n телофазу.
Митоз n Митоз включает n 4 основные фазы: n профазу n метафазу, n анафазу и n телофазу.
 Профаза n n Профаза начинается с конденсации хромосом, которые становятся видимыми в световой микроскоп как нитевидные структуры. Каждая хромосома состоит из двух параллельно лежащих сестринских хроматид, связанных в области центромеры. Ядерная оболочка распадается на мембранные пузырьки и исчезает к концу профазы, так же как и ядрышко. Кариоплазма смешивается с цитоплазмой. Пары центриолей расходятся к противоположным полюсам клетки и дают начало микротрубочкам митотического веретена.
Профаза n n Профаза начинается с конденсации хромосом, которые становятся видимыми в световой микроскоп как нитевидные структуры. Каждая хромосома состоит из двух параллельно лежащих сестринских хроматид, связанных в области центромеры. Ядерная оболочка распадается на мембранные пузырьки и исчезает к концу профазы, так же как и ядрышко. Кариоплазма смешивается с цитоплазмой. Пары центриолей расходятся к противоположным полюсам клетки и дают начало микротрубочкам митотического веретена.
 метафаза n n В метафазе хромосомы выстраиваются в области экватора митотического веретена (в равной удаленности от центриолей противоположных полюсов), и образуют картину экваториальной (метафазной) пластинки (вид сбоку) или материнской звезды (вид со сторону полюсов). Сестринские хроматиды к концу этой фазы разделяются щелью, однако удерживаются в области центромеры.
метафаза n n В метафазе хромосомы выстраиваются в области экватора митотического веретена (в равной удаленности от центриолей противоположных полюсов), и образуют картину экваториальной (метафазной) пластинки (вид сбоку) или материнской звезды (вид со сторону полюсов). Сестринские хроматиды к концу этой фазы разделяются щелью, однако удерживаются в области центромеры.
 Анафаза n n n Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды (в области центромеры) и движения дочерних хромосом к противоположным полюсам клеток, происходящего вдоль микротрубочек. Анафаза завершается скоплением на полюсах клетки двух идентичных наборов хромосом, которые образуют картину звезд (стадия дочерних звезд). В конце анафазы начинает образовываться клеточная перетяжка, благодаря сокращению актиновых микрофиламентов, концентрирующихся по окружности клетки.
Анафаза n n n Анафаза начинается с синхронного расщепления всех хромосом на сестринские хроматиды (в области центромеры) и движения дочерних хромосом к противоположным полюсам клеток, происходящего вдоль микротрубочек. Анафаза завершается скоплением на полюсах клетки двух идентичных наборов хромосом, которые образуют картину звезд (стадия дочерних звезд). В конце анафазы начинает образовываться клеточная перетяжка, благодаря сокращению актиновых микрофиламентов, концентрирующихся по окружности клетки.
 Телофаза n n Телофаза характеризуется реконструкцией ядер дочерних клеток и завершением их разделения. Ядерная оболочка восстанавливается, хромосомы постепенно деспирализуются, замещаясь картиной хроматина интерфазного ядра, а в конце телофазы вновь появляется ядрышко. Углубление клеточной перетяжки завершается полной цитотомией с формированием двух дочерних клеток. При этом происходит распределение органелл между дочерними клетками
Телофаза n n Телофаза характеризуется реконструкцией ядер дочерних клеток и завершением их разделения. Ядерная оболочка восстанавливается, хромосомы постепенно деспирализуются, замещаясь картиной хроматина интерфазного ядра, а в конце телофазы вновь появляется ядрышко. Углубление клеточной перетяжки завершается полной цитотомией с формированием двух дочерних клеток. При этом происходит распределение органелл между дочерними клетками


 Митоз клетки
Митоз клетки
 Эндомитоз и полиплоидизация n Эндомитоз – процесс увеличения числа хромосом внутри ядерной оболочки без последующего деления клетки, что приводит к повышенному содержанию ДНК в ядре – полиплоидии. n Полиплоидные ядра имеют больший объем. Полиплоидные клетки могут также возникнуть вследствие митотического деления без последующей цитотомией. При таком делении образуются двуядерные клетки с увеличенным вдвое набором хромосом. n Основной смысл развития полиплоидии заключается в усилении функциональной активности клеток. n Наличие полиплоидных – тетра- (4 n, если 1 n – гаплоидный набор хромосом) и октаплоидных (8 n) клеток – нормальное явление для гепатоцитов (клеток печени), переходного эпителия мочевого пузыря, секреторных клеток поджелудочной и слюнных желез. n Уровень полиплоидизации мегакариоцитов красного костного мозга достигает – 16 -32 n.
Эндомитоз и полиплоидизация n Эндомитоз – процесс увеличения числа хромосом внутри ядерной оболочки без последующего деления клетки, что приводит к повышенному содержанию ДНК в ядре – полиплоидии. n Полиплоидные ядра имеют больший объем. Полиплоидные клетки могут также возникнуть вследствие митотического деления без последующей цитотомией. При таком делении образуются двуядерные клетки с увеличенным вдвое набором хромосом. n Основной смысл развития полиплоидии заключается в усилении функциональной активности клеток. n Наличие полиплоидных – тетра- (4 n, если 1 n – гаплоидный набор хромосом) и октаплоидных (8 n) клеток – нормальное явление для гепатоцитов (клеток печени), переходного эпителия мочевого пузыря, секреторных клеток поджелудочной и слюнных желез. n Уровень полиплоидизации мегакариоцитов красного костного мозга достигает – 16 -32 n.
 Строение митотического веретена
Строение митотического веретена
 Интерфаза
Интерфаза
 Профаза
Профаза
 Метафаза
Метафаза
") Метафаза (материнская звезда)
Метафаза (материнская звезда)
 Метафаза
Метафаза
 Анафаза
Анафаза
 Ранняя телофаза
Ранняя телофаза
 Поздняя телофаза
Поздняя телофаза
 Митоз
Митоз
 Митоз
Митоз