срс (2).pptx
- Количество слайдов: 23



Тексерген: Жүнісбаева. Ж Орындаған: Каримова. Л Мбт 204

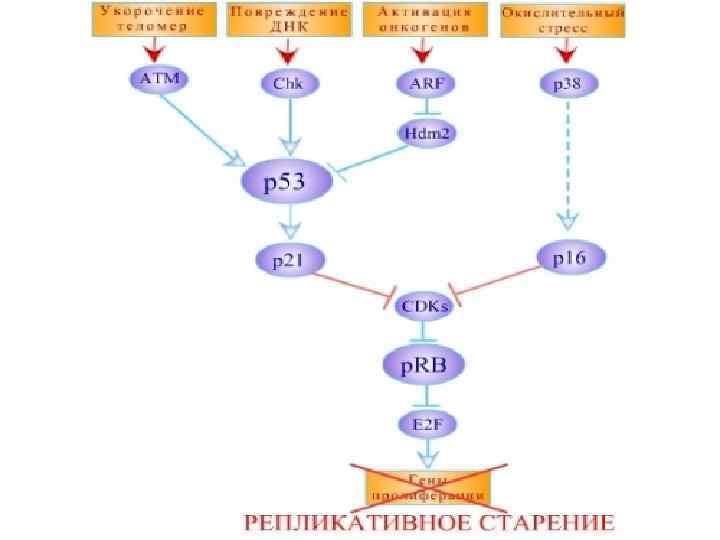
Ген p 53 относится к группе генов "общего контроля". К ним относят и те гены-супрессоры, продукты которых не входят в системы репарации ДНК непосредственно, а принимают участие в организации "контрольного пункта" проверки ДНК перед переходом клетки к следующей стадии клеточного цикла, обеспечивающего две основных контролирующих функции: - проверку того, что предыдущая стадия завершена полностью, и - в случае необходимости создания возможности для прохождения репарации ДНК перед началом репликации. В частности, это означает, что разрывы в цепи ДНК приводят к задержке клеточного цикла в фазе G 1, препятствуя переходу клетки в фазу S. Ключевую роль в этом процессе играет один известный ген-супрессор - p 53, мутации или делеции которого наблюдали примерно в 50% всех злокачественных заболеваний.
 — это транскрипционный фактор, регулирующий клеточный цикл. p 53")
p 53 (белок p 53) — это транскрипционный фактор, регулирующий клеточный цикл. p 53 выполняет функцию супрессора образования злокачес твенных опухолей, соответственно ген. TP 53 является антионкогеном. Мутации гена TP 53 обнаруживаются в клетках около 50 % раковых опухолей. Зачастую его называют «стражем генома» .

Ген человека, кодирующий белок p 53, называется TP 53 (курсив указывает на то, что это название гена, а не белка). Этот ген расположен на хромосоме 17 (17 p 13. 1). Локализация гена в геноме других организмов: Мышь — хромосома 11 Крыса — хромосома 10 Собака — хромосома 5 Свинья — хромосома 12

Человеческий белок p 53 состоит из 393 аминокислотных остатков и имеет 5 доменов: N-концевой домен, активирующий транскрипцию(англ. transcriptio n-activation domain; TAD), аминокислотные остатки 1 -42; Богатый пролином домен, важный для апоптотической активности p 53, аминокислотные остатки 80 -94; ДНК-связывающий домен ( «цинковый палец), аминокислотные остатки 100— 300; Домен, отвечающий за олигомеризацию, аминокислотные остатки 307— 355. Тетрамеризация очень важна для активности p 53 in vivo;

C-концевой домен, задействованный в отсоединении ДНК-связывающего домена от ДНК, аминокислотные остатки 356— 393. Мутации, инактивирующие p 53 при раковой трансформации, обычно затрагивают ДНКсвязывающий домен. Эти мутации приводят к неспособности белка p 53 связываться с ДНК и, следовательно, выполнять функцию активатора транскрипции. Такие мутации обычно являются рецессивными. В случае мутаций в домене, отвечающем за олигомеризацию, мутантный белок зачастую способен образовывать димеры с белком дикого типа, инактивируя его. Такие мутации являются доминантными.

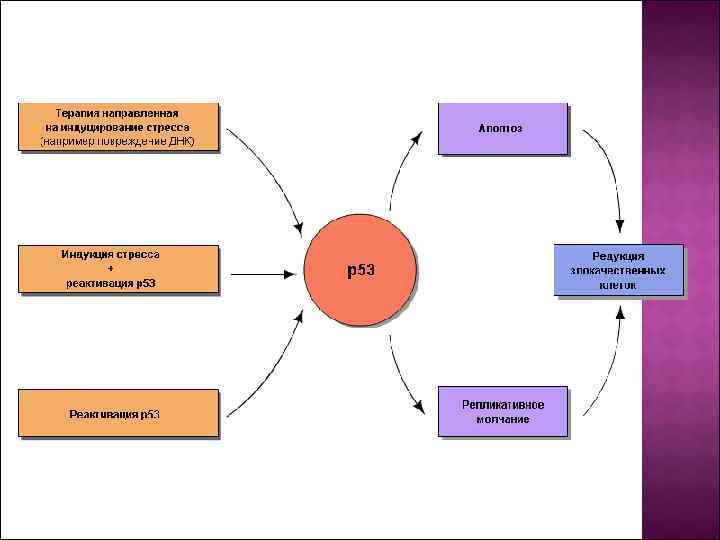
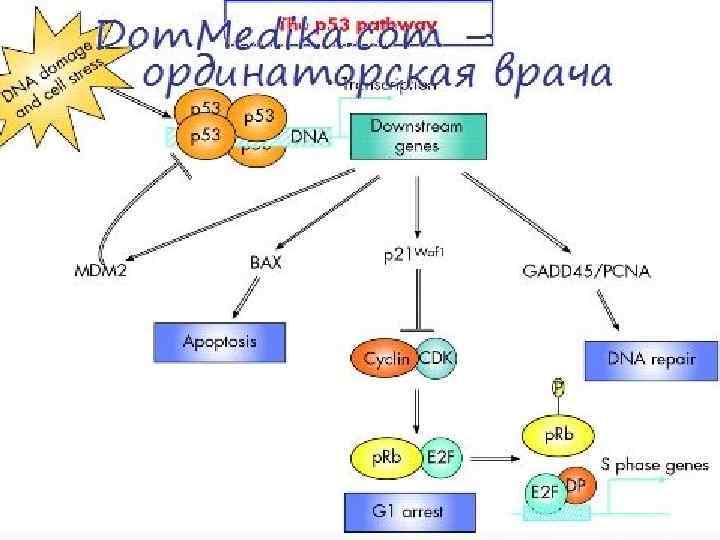
Белок р53 является продуктом гена-супрессора опухоли. TP 53 и экспрессируется во всех клетках организма. При отсутствии повреждений генетического аппарата белок р53 находится в неактивном состоянии, а при появлении повреждений ДНК активируется. Активация состоит в приобретении способности связываться с ДНК и активировать транскрипцию генов, которые содержат в регуляторной области нуклеотидную последовательность, которая называется p 53 -response element (участок ДНК, с которым связывается белок р53). Таким образом, р53 — фактор, который запускает транскрипцию группы генов и который активируется при накоплении повреждений ДНК. Результатом активации р53 является остановкаклеточного цикла и репликации ДНК; при сильном стрессовом сигнале — запуск апоптоза.

Белок р53 активируется при повреждениях генетического аппарата, а также при стимулах, которые могут привести к подобным повреждениям, или являются сигналом о неблагоприятном состоянии клетки (стрессовом состоянии). Функция белка р53 состоит в удалении из пула реплицирующихся клеток тех клеток, которые являются потенциально онкогенными (отсюда образное название белка р53 — англ. guardian of the genome — хранитель генома). Данное представление подтверждается тем фактом, что потеря функции белка р53 может быть установлена в ~50 % случаев злокачественных опухолей человека. В регуляции активности белка р53 ведущая роль принадлежит посттрансляционным модификациям белка и его взаимодействиям с другими белками.

Активация белка р53 происходит в ответ на многочисленные стрессовые стимулы: непосредственные повреждения ДНК (классический стимул); повреждения аппарата сегрегации генетического материала (например, митотического веретена); уменьшение концентрации свободных рибонуклеотидов; гипоксия; тепловой шок; высокая концентрация NO (монооксида азота); ионизирующее излучение.
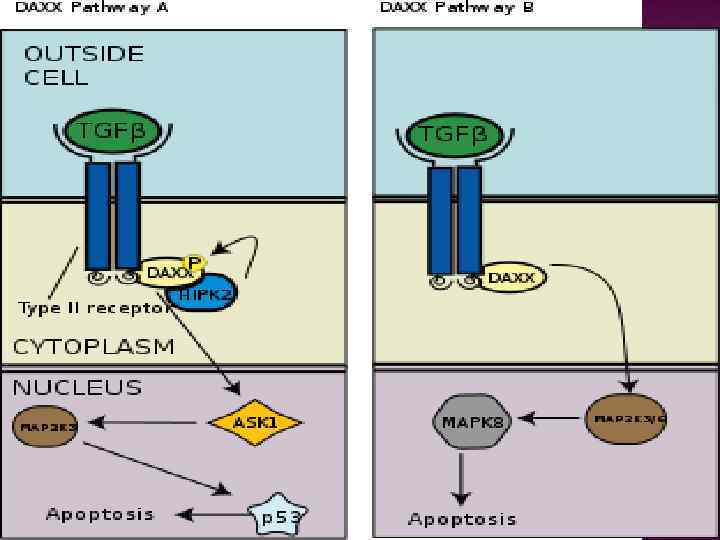
 клетках было обнаружено увеличение концентрации белка р53 по сравнению с")
В быстро делящихся(пролиферирующих) клетках было обнаружено увеличение концентрации белка р53 по сравнению с делящимися медленно. Значение увеличения концентрации р53 в данном случае в том, что клетки, которые быстро реплицируют ДНК, более подвержены возникновению повреждений генетического аппарата, чем, например, неделящиеся клетки в фазе G 0. Следовательно, увеличение концентрации р53 — это подготовка клетки для быстрой реакции на возможное возникновение повреждений ДНК. Очевидно, что для остановки клеточного цикла в условиях стимуляции пролиферации внеклеточными ростовыми факторами требуется более высокая концентрация р53, чем в условиях фазы G 0. Вследствие строгого посттрансляционного контроля активации белка р53, высокая концентрация белка р53 сама по себе не ведёт к его активации.
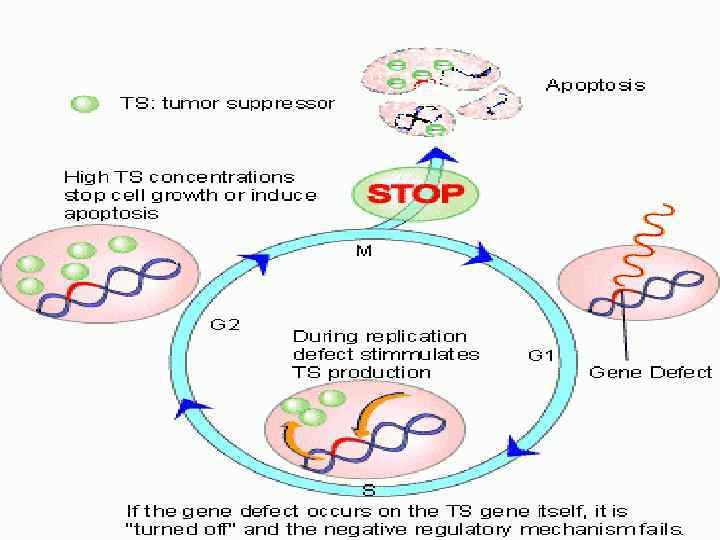

Активированный белок р53 является специфическим транскрипционным фактором. Гены, транскрипцию которых стимулирует белок р53, кодируют белкикомпоненты апоптотической программы (проапоптотические компоненты) и белки, которые регулируют клеточный цикл.

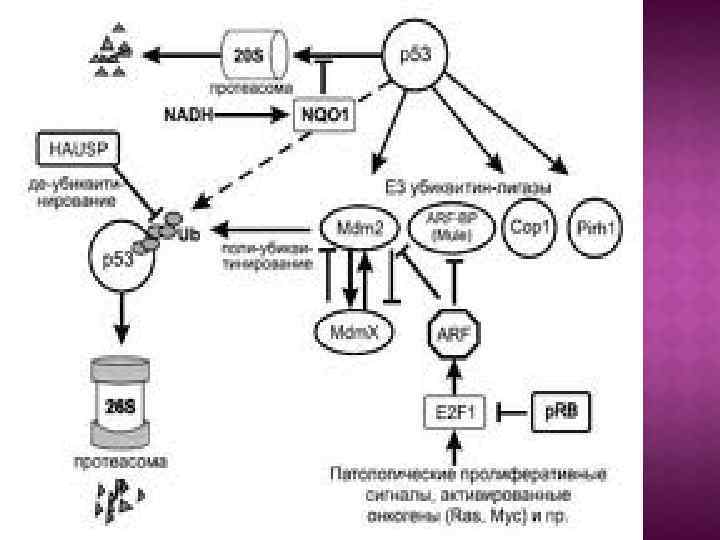
Активированный белок р53 супрессирует транскрипцию ряда генов. Этот супрессирующий эффект не связан с супрессорной функцией комплекса Mdm 2: p 53, так как данный комплекс супрессирует транскрипцию тех генов, которые активируются белком р53. В то же время супрессорный эффект белка р53 касается другого набора генов. Репрессия транскрипции по крайней мере частично объясняется тем, что белок р53 формирует комплексы с неспецифическими транскрипционными факторами, среди них белок TBP белок CBF и белок SP-1

При нормальном увеличении активности протоонкогенов (например, в результате передачи пролиферативного сигнала от рецепторов цитоплазматической мембраны) активация протоонкогенов носит временный характер и значимого увеличения концентрации p 19 ARF не происходит.

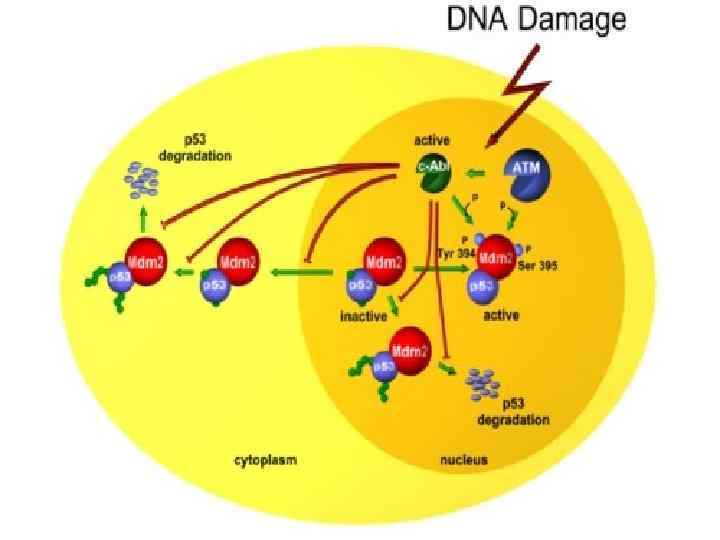

срс (2).pptx