T-лимфоциты.ppt
- Количество слайдов: 54



Т-лимфоциты Лекция № 6 МБФ

Иммунологические исследования ллимфоцитов n показали гетерогенность популяций, включающих в себя 3 типа зрелых Т-л и 3 типа В-л. В ходе дифференцировки и созревания, л-ты заселяют вначале ЦО ИС, где проходят дальнейшую дифференцировку, а затем периферические органы ИС. Благодаря непрерывной циркуляции л-тов с кровью и лимфой, в организме происходит постоянный иммунологический контроль как антигенного состава собственных клеток, так и поступления чужеродного материала извне.

Иммунокомпетентные клетки Антигенпрезентирующие клетки n Т – лимфоциты (Т – хелперы, Т – киллеры, γδТлимфоциты) n В -лимфоциты n

Характеристика лимфоцитов Лимфобласт – круглая крупная клетка. Ядро круглое или овальное, светло-фиолетовое с мелкозернистым хроматином, содержит 1 или 2 ядрышка. Цитоплазма базофильная. ЧЕТКО ОЧЕРЧЕННАЯ С УСИЛИВАЮЩЕЙСЯ базофилией к краям. n Лимфоцит – клетка размером 5 -15 мкм, ядро круглое или овальное, окрашено интенсивно в синий цвет. Цитоплазма базофильная с перинуклеарной зоной, если содержится зернистость, то она носит гранулярный характер. В цитоплазме находится сложная вакуоль, ограниченная мембраной и содержит несколько плотных крошечных пузырьков. Различают малые, средние и большие лимфоциты по размеру, по происхождению Т и В-л. n

Характеристика лимфоцитов Т-л по морфологии ядро четкое, компактное. Рецепторов немного. Резоткообразование дает с эритроцитами барана (3 -4). РБТЛ с ФГА. n В-л по морфологии ядро зернистое, диффузное. Рецепторов много. Резеткообразование с эритроцитами мыши (много). РБТЛ с ГР(-)бактериями. n Плазматическая клетка – диаметр 8 -12 мкм, ядро расположено эксцентрично, округлой или овальной формы, хроматин имеет колесовидную исчерченность. В интенсивной окраске цитоплазмы отмечается светлая околоядерная зона. n

Характеристика лимфоцитов Лимфоцитограмма - имеют округлый вид в состоянии покоя или вытянутых клеток в состоянии активного передвижения. Величина л-тов 5 -13 мкм. Форма и размер л-тов зависит от того, находятся ли они во взвешенном состоянии или прикреплены к какомулибо субстрату. Морфологически различают малые неактивные л-ты, средние и большие. В зависимости от ядерно-цитоплазматического соотношения различают узкоплазменные и широкоплазменные лты. n В крови у здоровых людей лимфоцитограмма состоит из малых лимфоцитов 62 -72%, средних 17 -22%, больших до 7%. n

Иммунологические показатели крови n n n n Общее количество лимфоцитов 1, 52, 5 х109 кл/л В-л 0, 2 -0, 5 х109 (10 -15%) Т-л 1 -2 х109 (55 -75%) Тh CD 4 0, 4 -0, 8 х109 Tk. CD 8 0, 1 -0, 6 х109 Ig. G 7 -24 г/л; Ig. M 0, 7 -1, 8 г/л Ig. A 1, 3 -5 г/л Ig. D 0, 01 Ig. E 100 к. Ед-л

Дифференцировка Т-л Сущность дифференцировки Т-л состоит в экспрессии антигенраспознающего рецептора и необходимых сервисных молекул – это мембрнные или секретируемые цитокины. Они обеспечивают взаимодействие Т-л с другими клетками организма. n Антигенраспознающие рецепторы Т-л обозначаются TCR (T-cell recеptor) состоят из цепей, принадлежпщих к суперсемейству иммунолобулинов, т. е. TCR кодируется генами родственниками генов иммуноглобулинов. Молекула TCR представляет из себя как бы 1 -4 молекулы иммуноглобулина – аналог одного Fab фрагмента. TCR являются гетеродимером – состоит из двух разновеликих полипептидных цепей. n

Строение молекулы Ig

Дифференцировка Т-л n Известно 2 варианта TCR. В одной паре цепи обозначаются – Т α и β (альфа и бета), вторая – T γ и δ (гамма и дельта). Эти варианты различаются составом полипептидных цепей антигенраспознающего участка. Каждый индивидуальный лимфоцит несет только один вариант рецептора α-β либо γ-δ.

Антигенраспознающий рецептор Т-л строение антигенраспознающего рецептора Т-л на примере α и β TCR. n Состоит из 8 или 10 полипептидных цепей (одна или две пары α альфа, β бета и комплекс из двух ε эпсилон и ξ дзета по одной δ дельта и γ гамма цепей). n

Антигенраспознающий рецептор Т-л n n Трансмембранные цепи альфа и бета TCR. Это 2 примерно одинаковые по размеру полипептидные цепи – альфа- кислый гликопротеин, бета-нейтральный или основной гликопротеин. Каждая из этих цепей содержит по 2 гликолизированных домена во внеклеточной части рецептора, гидрофобную трансмембранную часть и короткий цитоплазматический участок. Внеклеточные части обеих цепей соединены одной дисульфидной связью. V-область, наружные внеклеточные домены, обеих цепей, имеют вариабельный аминокислотный состав. Они гомологичны V- области молекул иммуноглобулинов и составляют V-область TCR. Именно Vобласть альфа и бета цепей вступают в связь с комплексом МНСпептид. С-область, Проксимальные домены обеих цепей гомологичны константным областям иммуноглобулинов, это С-области TCR. Короткий цитоплазматический участок (как альфа так и бета-цепи) не может самостоятельно обеспечить проведение сигнала внутрь клетки. Для этого служат 6 дополнительных полипептидных цепей γ, δ, 2ε и 2ξ.

Антигенраспознающий рецептор Т-л Комплекс CD 3 Цепи γ, δ, ε (вместе их называют комплексом СD 3) необходимы для экспрессии α и β-цепей, их стабилизации и проведения сигнала внутрь клетки. Комплекс CD 3 состоит из внеклеточной трансмембранной (отрицательно заряженной и поэтому электростатически связанной с трансмембранными участками α и β-цепей) и цитоплазматической частей. n ξ-цепи соединены между собой дисульфидным мостиком и , будучи по большей части расположенными в цитоплазме, осуществляют проведение сигнала внутрь клетки. n
. Цитоплазматические участки полипептидных цепей γ, δ,")
Антигенраспознающий рецептор Т-л ITAM (иммунорецептор тирозинсодержащий активационные последовательности). Цитоплазматические участки полипептидных цепей γ, δ, ε, ξ содержат последовательности ITAM (1 последовательность γ и δ-цепях, 2 - в ε-цепях, 3 - в каждой ξ-цепи), взаимодействующие с тирозинкиназами цитозоля (активация этих ферментов и составляет начало биохимических реакций по проведению сигнала). n В связывании антигена участвуют ионные, водородные, Ван-дер -ваальсовы и гидрофобные силы, конформация рецептора при этом существенно изменяется. Каждый TCR потенциально способен связывать порядка 105 разных антигенов, причем не только родственные по строению, но и негомологичные по структуре. n

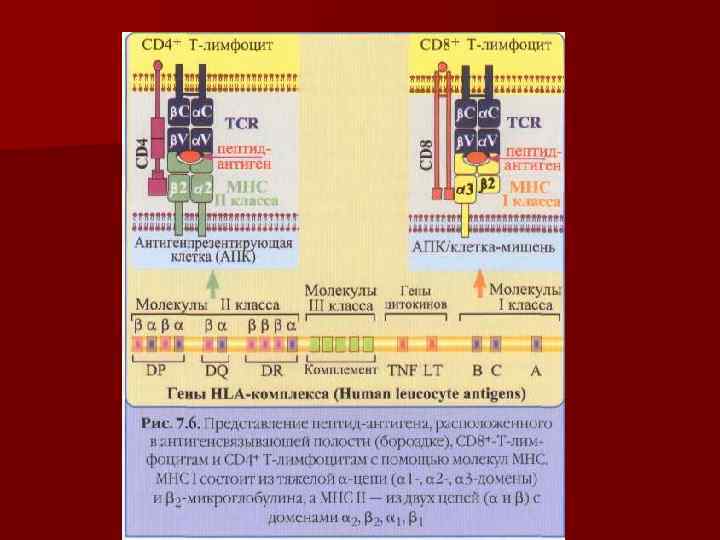
Корецепторные молекулы CD 4 и CD 8 Помимо самого TCR, каждый зрелый Т-л экспрессирует одну корецепторную молекулу – CD 4 или CD 8, которые также взаимодействуют с молекулами МНС на АПК или клеткахмишенях. Каждая из них имеет цитоплазматический участок, связанный с тирозинкиназой Lck, и , вероятно вносит свой вклад в проведение сигнала внутрь клетки при распознании антигена. n CD 4 связывается с инвариантной частью (β 2 -доменом) молекулы МНС 2. CD 4 имеет молекулярную массу 55 к. Да и 4 домена во внеклеточной части. При активации Т-л одну молекулу TCR обслуживают 2 молекулы CD 4. n CD 8 cвязывается с инвариантной частью (α 3 -доменом) молекулы МНС 1. СD 8 гетеродимер цепей α и β соединенных дисульфидной связью. Во внеклеточной части каждая из цепей имеет по одному иммуноглобулиноподобному домену. n
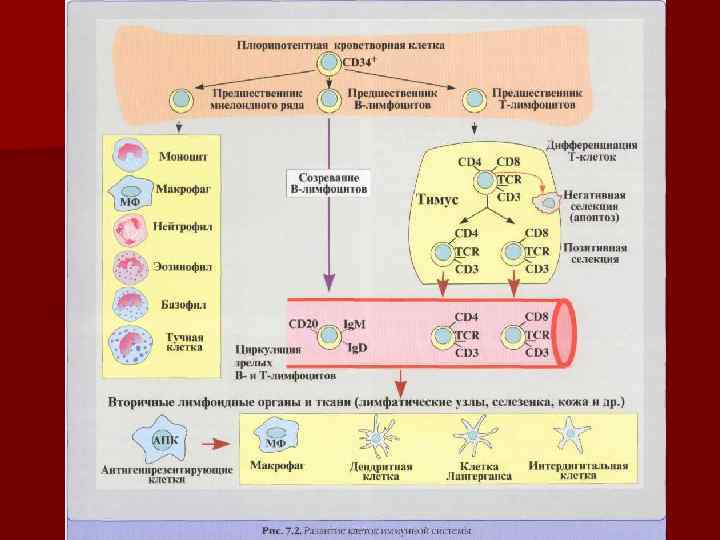
 РАЗВИВАЮТСЯ В ЭМБРИОНАЛЬНОЙ ПЕЧЕНИ И КОСТНОМ МОЗГЕ. Мигрируя")
Дифференцировка Т-л n Предшественники Т-л (пре-тимоциты) РАЗВИВАЮТСЯ В ЭМБРИОНАЛЬНОЙ ПЕЧЕНИ И КОСТНОМ МОЗГЕ. Мигрируя в тимус, претимоциты дифференцируются в ранние тимоциты (1 -я стадия), имеющие на своей поверхности мембранные маркеры СD 2, 7, 34. Происходит транскрипция генов гамма и перестройка (реаранжировка) генов бетацепи Т-клеточного рецептора. Эти клетки названы двойными (дважды) негативными, т. к. не содержат ни CD 4 и CD 8 (CD 4 минус и CD 8 минус).

Дифференцировка Т-л n. В следующей – 2 -ой стадии созревания обычный тимоцит содержит как CD 2, CD 7, так и одновременно (СD 4 плюс и CD 8 плюс) – двойные позитивные. Начинается перестройка генов β, γ, δцепей ТCR и формируется цитоплазматическая форма CD 3.

Дифференцировка Т-л n На короткое время с мембраны тимоцитов выпадают обе корецепторные молекулы, а затем экспрессируется одна из двух и образуются две популяции клеток, CD 4 или CD 8. CD 4 характерен для Т-х, а CD 8 – для цитотоксичных Т-клеток. Эти клетки обозначаются как однопозитивные. Более 95% тимоцитов удаляются в результате их положительной и отрицательной клональной селекции. Сначала происходит положительная селекция: отбираются тимоциты, несущие низкоаффинные TCR и взаимодействующие с поверхностными молекулами кортикальных эпителиальных клеток (с их МНС 2). Затем происходит отрицательная селекция: тимоциты с высокоаффинным TCR взаимодействуют с поверхностными комплексами МНС 1 плюс пептид антигенпрезентирующих клеток.

Дифференцировка Т-л Завершая антигеннезависимую дифференцировку, зрелые наивные Т -л покидают тимус через кровь и расселяются в тимусзависимых зонах вторичных лимфоидных органов и в барьерных тканях. При встрече Т-л с чужеродным Аг, происходит его активация, отбор (селекция клона одной специфичности) и размножение с увеличением популяции однородных клеток (экспансия клона). n CD 8 Tкл играют роль цитотоксических Т-л- они распознают и непосредственно убивают клетки модифицированные вирусом, опухолевые и др. n CD 4 Ткл -функция их разнообразна. Значительная часть становится Тх и взаимодействует с В-л и др клетками прямом контакте или через цитокины. Некоторые клетки могут развиваться CD 4 ЦТЛ при синдроме Лайелла. n

Маркеры Т-лимфоцитов n n n n CD 1 - МНС I класса, представление Аг CD 2 – адгезия Т- киллеров на клетки-мишени, к эндотелию CD 3 – TCR – проведение сигнала от рецептора путем активации тирозинкиназы CD 4 – маркер Тh, ко-рецептор TCR CD 8 – маркер Tc лимфоцитов, ко-рецептор TCR CD 25 – маркер активированных лимфоцитов, рецептор для ИЛ 2 CD 28 – рецептор Th взаимодействует с костимулирующим факторм АПК (CD 80/86) CD 30 – усиливает пролиферацию Т- и В – клеток, после связывания с лигандом

Поверхностные маркеры Т-л

Т-лимфоциты
")
Субпопуляции CD 4+Т-л (Тh)
 С концо 80 -х гг ХХв. Было принято выделять 2")
Субпопуляции CD 4+Т-л (Тh) С концо 80 -х гг ХХв. Было принято выделять 2 субпопуляции Тх( в зависимости от того , какой набор цитокинов они продуцируют)Th 1 Th 2. В последние годы спектр расширяется – Тh 17. n Трегуляторы, Тr 1, Тh 3, Тfh и др. n Тh экспрессирующие мембранные маркеры CD 4, подразделяют на Т-л с эффекторными функциями (Th 1 Th 2 Th 17) и Т-регуляторные клетки – естественные Treg и индуцированные - Th 3 или Tr 1. n

CD 4+Т-л СD 4 Т-клетки n Тh 0 – CD 4 Т-л на ранних стадиях развития иммунного ответа, продуцируют Ил-2 (митоген для всех лимфоцитов) n Тh 1 - дифференцированная субпопуляция СD 4 Т-л , продуцируют ИНФ гамма, ФНО бета, ИЛ-2; регулирует реакции клеточного иммунитета, ГЗТ, активирует ЦТЛ; стимулирует продукцию В-л –ми Ig. G, которые активируют систему комплемента. Развитие избыточного воспаления с последующим повреждением тканей связано с активностью Th 1. n

CD 4+Т-л Th 2 - дифференцированная субпопуляция СD 4 Т-л , продуцируют ИЛ-4, 5, 6, 10, 13. Активирует В-л и способствует секреции большого количества Ig и особенно Ig. E. Участвует в активации эозинофилов и развитии аллергических реакций. n Th 17 субпопуляция CD 4 T-л , продуцирует Ил 17, играют ключевую роль в аутоиммунных заболеваниях. n

CD 4+Т-л Т-регуляторы – CD 4 Т-л, это клетки подавляющие активность др. клеток ИС поддействием иммуносупрессорных цитокинов – ИЛ 10(ингибитор активности макрофагов и Тh 1) и трансформирующий фактор роста (TGF) бета- ингибитора пролиферации лимфоцитов. На мембране некоторых Т-регуляторов экспрессированны индукторы апоптоза Fas. L (Fas-лиганд). n Естественные Treg созревающие в тимусе ( CD 4+CD 25+ экспрессируют фактор транскрипции Foxp 3), и индуцированные Treg – CD 4+Т-л локализованы в слизистых оболочках ЖКТ, продуцируют трансформирующий фактор роста (TGF) бета (Th 3) или Ил-10 ( Tr 1). n Нормальное функционирование Т-регуляторов контролируют интенсивность иммунного ответа, подавляя активность других субпопуляций Т-л. Обеспечивает гомеостаз ИС и предотвращение аутоиммунных заболеваний. n

Другие CD 4+Т-л Tfh-фолликулярный хелпер- расположены в лимфоидных фолликулах и осуществляет хелперную функцию для В-л посредством продукции Ил-21, вызывая их созревание, и термальную дифференцировку в плазматические клетки. Продукция Ил-6 и 10 обеспечивает дифференцировку В-л, нарушение приводит к – развитию аутоиммунных заболеваний и иммунодефицитам. n Th 9 - продуцент Ил-9, способен вызывать пролиферацию Тх, при отсутствии антигенной стимуляции, а также усиливать секрецию В-ми Ig. M, G, E.
 n n Цитотоксические Т-л. (ЦТЛ или CTL) c помощью TCR")
СD 8+ Т-л (цитотоксические) n n Цитотоксические Т-л. (ЦТЛ или CTL) c помощью TCR – Т-кл рецептора и корецептора CD 8 распознают комплекс антигенный пептид плюс МНС 1 класса. Распознавание Аг-пептида усиливается дополнительным сигналом в виде ИЛ-2 от Тх1, что вызывает пролиферацию ЦТЛ с образованием антигенспецифического клона ЦТЛ. Далее ЦТЛ выбрасывают из гранул цитотоксические белки: перфорины, гранулизины и гранзимы (сериновые протеазы). Перфорины и гранулизины, встраиваясь в мембрану клетки мишени, образуют поры, способствующие проникновению гранзимов, которые запускают апоптоз клетки-мишени. Нарушение осмотического баланса с внеклеточной средой приводит к гибели клетки.
 n Кроме того, апоптоз вызывается в результате взаимодействия Fas-рецептора CD")
СD 8+ Т-л (цитотоксические) n Кроме того, апоптоз вызывается в результате взаимодействия Fas-рецептора CD 95 клетки-мишени с Fas-лигандом (Fas. L) ЦТЛ (CD 95 рецепторы находятся на СD 4 CD 8, NК, паренхиматозных клетках сетчатки и роговицы, клетках эндотелия сосудов). За цитотоксические свойства ЦТЛ отвечает также фактор некроза опухолей. После уничтожения клетки мишени ЦТЛ способны к разрушению других клеток-мишений (серийные убийцы). Ранее такие ЦТЛ называли Ткилерами, однако оказалось, что и другие клетки способны к уничтожению клеток-мишений. Так цитотоксическими свойствами могут обладать Тх1 у которых в основном активируется Fas-система. Тх1, соединяясь своими Fas-лигандом с Fas-рецептором клетки-мишени, передают им сигнал апоптоза. Возможно около 10% ЦТЛ являются CD 4 Т-лимфоцитами.

Т-лимфоциты памяти n Долгоживущие потомки Т-л, Т-л памяти являются носителями рецепторов к антигенам, полученным от Т-л, ранее сенсибилизированных ими. Т-л памяти часто встречаются, обладают высокой экспрессией мембранных молекул, меньшей потребностью в провоспалительных медиаторах и корецепторных сигналах для развития вторичного иммунного ответа. Они отличаются появлением CD 45 RO изоформы тирозинфосфатазы (наивные Т-л экспрессруют изоформу CD 45 RA) и CD 29 – рецептора для фибронектина. Т-л памяти обозначаются как CD 4+СD 45 RO+ и CD 8+CD 45 RO+.

γδТ-лимфоциты

γδ Т-лимфоциты 99% Т-л, проходящих лимфопоэз в тимусе, составляют αβТ-клетки, менее 1% - γδТклетки. γδТ-клетки в эмбриогенезе появляются раньше, чем αβТ-клетки. n γδТ-клетки не экспрессируют CD 4. Молекула СD 8 экспрессирована на части γδТклеток, но не в виде αβ-гетеродимера, как на Тαβ СD 8 клетках, а в виде гомодимера из 2 -х α-цепей. n

γδ Т-лимфоциты Антигенраспознающие свойства: γδТCR в большей степени напоминают иммуноглобулины, чем αβТCR , т. е. способны связывать нативные антигены независимо от классических молекул МНС-для γδТ-клеток не обязателен или вовсе не нужен предварительный процессинг антигена АПК. n Разнообразие γδТCR меньше, чем αβТCR, или иммуноглобулинов, хотя в целом γδТклетки способны распознавать широкий спектр антигенов (микобактерии, углеводы, белки теплового шока). n

γδ Т-лимфоциты n Функции γδТ-клеток –связующее звено между врожденным и приобретенным иммунитетом. γδТклетки –первый барьер для патогенов, секретируют цитокины, иммунорегуляторыи способны дифференцироваться в ЦТЛ.

Т-лимфоциты

Взаимодействие клеток в ходе гуморального иммунного ответа
, экспрессированную")
Взаимодействие клеток в ходе гуморального иммунного ответа n TCR распознает антигенную детерминанту (эпитоп), экспрессированную на поверхности АПК вместе с молекулой МНС-2. Во взаимодействии участвует маркерная молекула Тh-СD 4. В результате подобного взаимодействия АПК секретирует ИЛ-1, стимулирующий в Т-х синтез и секрецию Ил 2, а также синтез и встраивание в плазматическую мембрану того же Т-х рецепторов для Ил-2.

Взаимодействие клеток в ходе гуморального иммунного ответа n Отбор В-л происходит при взаимодействии антигена с Fab-фрагментом антител на поверхности этих клеток. Эпитоп этого антигена в комплексе с молекулой МНС-2 распознает рецептор Т-х, после чего Т-л секретирует цитокины, стимулирующие пролиферацию В-л и их дифференцировку в плазматические клетки, синтезирующие антитела к данному антигену.

Взаимодействие АПК и Тх


Маркеры АПК

Распознавание антигенов бактерий

Распознавание Т- и Влимфоцитами

Маркеры Т-лимфоцитов

РБТЛ

Взаимодействие АПК и Тх

Субпопуляции Thлимфоцитов

Взаимодействие ТCD 4+ и TCD 8+ лимфоцитов

Гуморальный иммунный ответ

Первичный и вторичный иммунный ответ

Клеточный иммунный ответ
T-лимфоциты.ppt