ген.ppt
- Количество слайдов: 20



Тұқым қуалау материалдарының дифференцировкаға әсері Орындаған: Бекишова А. Бт 31 Тексерген: Шорин С. С

тұқым қуалаушылық Тұқым қуалаушылық - ұрпақтар арасындағы материалдық және функционалдық сабақтастықты қамтамасыз ететін тірі организмдерге тән қасиет. Тұқым қуалаушылыққа байланысты тірі организмдердің морфология, физиология және биохимия құрылымы мен жеке даму ерекшеліктері ұрпақтан ұрпаққа беріледі. Организмдердегі ұқымқуалаушылық факторларының болатынын алғаш болжам жасап, тұқым қуалау заңдылықтарын ашқан Г. Мендель болды. Ол ата-аналық дарабастарды бір-бірінен бір не бірнеше белгілері бойынша ажыратылады, ал ол факторлар ата-аналарынан ұрпақтарына жыныс клеткалары арқылы беріледі деген қорытынды жасады (Мендель заңдары). 1909 жылы дат биологы В. Иогансен (1857 – 1927) бұл тұқым қуалау факторларын ген деп атады. 1911 жылы америкалық биолог Т. Морган (1866 – 1945) және оның әріптестері ұсынған “Тұқымқуалаушылықтың хромосомалық теориясы” бойынша да тұқымқуалаушылықтың бірлігі – ген деп көрсетілген. Гендер клетка ядросындағы хромосомаларда тізбектеле, бір сызықтың бойында орналасқан және әрбір геннің хромосомада нақты тұрақты орны (локусы) болады. Кез келген хромосома өзінің гендер тобымен ерекшеленеді.

Генетика ғылымының даму барысында тұқым қуалау факторлары тек ядрода ғана емес, клетка цитоплазмасының кейбір органоидтарында (митохондрияда, хлоропластарда) да кездесетіні анықталды. Осыған байланысты цитоплазмалық тұқымқуалаушылық жайлы ілім қалыптасты. Тұқымқуалаушылық материалының сақталуы, екі еселенуі және ұрпақтан ұрпаққа берілуі нуклеин қышқылдарына (ДНҚ және РНҚ) байланысты болады. [1] Тұқымқуалаушылық клеткада жүретін репликация (генетик. ақпаратты дәл көшіруді және оның ұрпақтан ұрпаққа берілуін қамтамасыз ететін нуклеин қышқылдары макромолекулаларының өздігінен жаңғыруы), транскрипция (ДНҚ-да жазылған генетик. ақпаратты жұмсаудың алғашқы кезеңі) және трансляция (ақпараттық РНҚ молекулаларындағы нуклеидтердің бірізділігі түрінде “жазып алынған” генетик. ақпаратты “есептеу”) процестерімен тығыз байланысты. Бұл кезде комплементарлық принципке сай ДНҚ және РНҚ молекулаларының айна қатесіз көшірмелері алынып, түзілетін белоктың құрамындағы амин қышқылдарының орналасу реті дәл анықталады. Мұның нәтижесінде тұқым қуалайтын нақты белгілі болады. Жер бетінде тіршіліктің пайда болуы мен дамуында тұқымқуалаушылық шешуші рөл атқарады. Өйткені эволюция барысында қалыптасқан тіршілікке қажетті жаңа белгілермен басқа да өзгерістер осы тұқымқуалаушылыққа байланысты ұрпақтан ұрпаққа беріліп, бекітіліп отырады. Тұқымқуалаушылықтың негізінде органдардың алуан түрлі топтары қалыптасты, дербес және біртұтас жүйелер (популяциялар, түрлер) құрылып, олардың тіршілік етуіне және қоршаған орта жағдайларына сай бейімділіктің сақталуына мүмкіндік туды. Сондықтан да тұқымқуалаушылық эвол. әрекеттің негізгі қозғаушы күшінің бірі болып табылады. Табиғатта тұқымқуалаушылық өзгергіштікпен қатар жүреді. Ауыл шаруашылығы мен медицина үшін тұқымқуалаушылықтың заңдылықтарын зерттеп білудің маңызы зор. Тұқымқуалаушылықпен өзгергіштіктің заңдылықтарын генетика ғылымы зерттейді
 — құрамында фосфоры бар биополемерлер. Табиғатта өте")
Нуклеин қышқылдары (лат. nucleus — ядро) — құрамында фосфоры бар биополемерлер. Табиғатта өте көп тараған. Молекулалары нуклеотидтерден тұрады, бір нуклеотидтіқ 5'-фосфор арасындағы эфирлік байланысы мен келесі нуклеотидтің углевод қалдығының 3'-гидроксилі арасы эфир байланысымен нуклеин қышқылдары углеводты-фосфатты қаққасын калайды. Нуклеин қышқылдары жоғарғы полимерлі тізбектері ондаған немесе жүздеген нуклеотидтің қалдықтарынан тұрады. Олардың м. с. 105— 1010. Нуклеин қышқылдары құрамына кіретін мономерлерінің (дезокси- немесе рибонуклеотидтер) түріне қарай ДНҚ жәңе РНҚ деп бөлінеді. [1] Нуклеин қышқылдары тірі жасуша ядросының маңызды құрам бөлігі. Нуклеин қышқылдары (НҚ) рибонуклеин қышқылы (РНҚ) және дезоксирибонуклеин қышқылы (ДНҚ) болып екі үлкен түрге бөлінеді. Тірі организмнің құрамына нуклеин қышқылдарының екі түрі де кіреді. Нуклеин қышқылдары жоғары молекулалы гетерополимерлі қосылыстар

класификациясы
, фосфор қышқылы және моносахаридтер (рибоза мен")
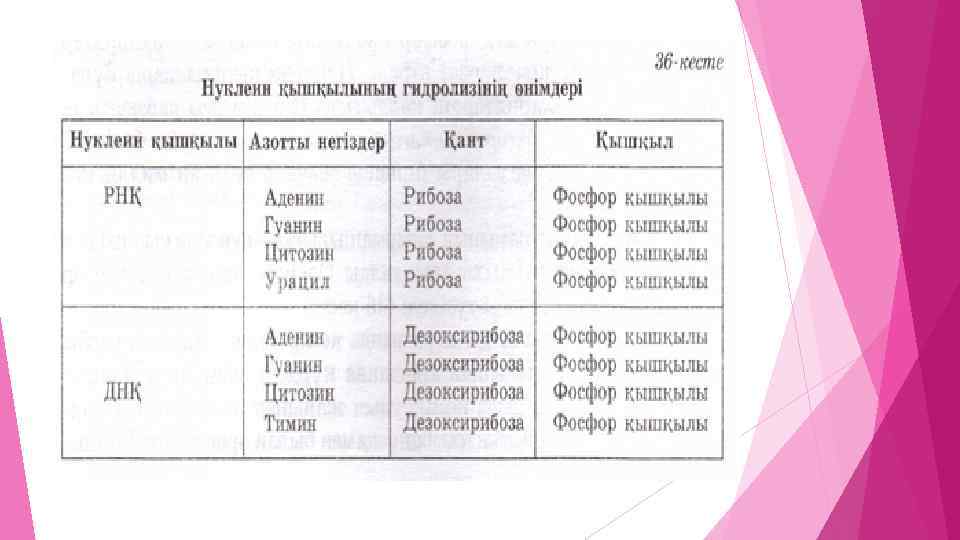
Яғни, нуклеин қышқылдарының құрамына азотты негіздер (пиримидинді, пуринді), фосфор қышқылы және моносахаридтер (рибоза мен дезоксирибоза) кіреді. Нуклеин қышқылдары құрамындағы моносахаридтердің қалдығына байланысты рибонуклеин қышқылы және дезоксирибонуклеин қышқылы болып екіге бөлінеді. ДНҚ молекулалық массалары бірнеше мыңнан ондаған миллионға жетеді. ДНҚ мен РНҚ құрамының айырмашылығы — нуклеин қышқылын толық гидролиздеу арқылы анықталды. Оларды гидролиздегенде, әр түрлі заттардың қоспасы түзіледі (36 кесте). Нуклеин қышқылдары құрамында көмірсудың гидроксил тобы мен фосфор қышқылының арасында күрделі эфирлік байланыс түзіледі, ал азотты негіз көмірсудың жанынан жалғасады. Полинуклеотидтің құрылысын сызбанұсқамен былай өрнектеуге болады:
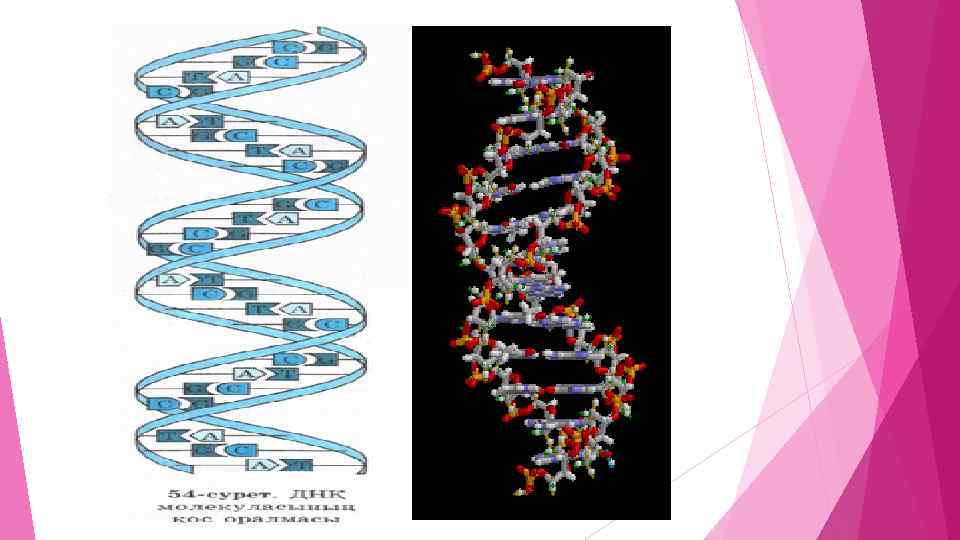

Нуклеин қышқылдары биологиялық тұрғыдан маңызды рөл атқарады. Олар тірі организмдердегі генетикалық ақпаратты сақтайтын және тасымалдайтын жасушаның (жасушаның) маңызды кұрам бөліктері болып табылады. Нуклеин қышқылдары ақуыз биосинтезіне қатысады және тірі организмдерде тұқым қуалаушылықты сақтап, оның бір ұрпақтан екінші ұрпаққа берілуін қамтамасыз етеді. ДНҚ жасуша ядросының хромосомасында (99%), рибосомаларда және хлоропластарда, ал РНҚ ядрошықтарда, рибосомаларда, митохондрияда, пластидтер мен дитоплазмада кездеседі. Олар жасушаның қай бөлігінде шоғырланса, соған байланысты қызмет атқарады. Жоғарыда айтылғандай, ДНҚ организмдегі тұқым қуалаушылық ақпаратты сақтайтын гендердің құрылыс материалы болып табылады. Ал РНҚ үш түрлі болғандықтан: рибосомдық (р-РНҚ); тасымалдаушы (т-РНҚ) және ақпараттық (а. РНҚ) әр түрлі қызметтер атқарады. ДНҚ мен РНҚ қызметтері 1940 жылдардан бастап анықталып, түрлі биологиялық тәжірибелер арқылы дәлелденген. Осы зерттеулер нәтижесінде молекулалық генетика ғылымы жедел дами бастады. Соңғы жылдары ғалымдар жоғары организмдердің гендерін бактериялар мен ашытқы саңырауқұлақтарының организміне енгізуді іске асырды. Соңынан оларды ақуыз синтездеуге пайдаланды. Мысалы, инсулин генін осылайша "жұмыс істеткізді". Адам инсулині ең алғаш рет Е. соlі деген бактерияның көмегімен 1982 жылы алынды. Осылайша бір типтегі организмнен алынған генді басқа типтегі организмге енгізуді гендік инженерия деп атайды. Жоғарыда айтылған ипсулин, өсу гормоны — соматотропин, сондай-ақ гемофилия ауруына қолданылатын VIII фактор — гендік инженерияның өнімдері. Қазіргі кезде гендік инженерияның көмегімен түрлі жұқпалы ауруларға қарсы вакциналар өндіріле бастады. Қазақстан Ұлттық ғылым академиясының академигі М. Ә. Айтхожин жасушалық макромолекулалардың (нуклеин қышқылдары мен ақуыздың) синтезі саласында өте маңызды зерттеу жұмыстарын жүргізді

Тірі ағзалардың химиялық құрамының біркелкі болуы. Тірі ағзалардың құрамында табиғатта кездесетін 70 -тен астам химиялық элементтер болады. Тірі азғалар мен өлі табиғаттың құрамындағы химиялық элементтердің мөлшері әр түрлі. Мысалы, өлі табиғаттың құрамында оттектен басқа кремний, темір, магний, алюминий, т. б. элементтер көбірек кездеседі. Тірі ағзалардың химиялық құрамының 98%-ын, негізінен, 4 химиялық элемент құрайды. Ондай элементтерге — көміртек, оттек, азот және сутек жатады. Тірі ағзаларда бұл элементтер күрделі органикалық молекулалардың түзілуіне қатысады. Өлі табиғатта бұл элементтер басқаша мөлшерде және өзгеше сапада болады. Өлі табиғатта кездесетін органикалық қосылыстар, негізінен, тірі ағзалардың тіршілік әрекеттерінен пайда болған. Тірі азғалар құрамындағы органикалық молекулалардың өздеріне тән ерекшеліктері бар және олар тірі ағзаларда белгілі бір қызмет атқарады.
 —")
Филогенез (гр. рһуіоп — түр, туыс және гр. genesis — шығу тегі) — организмдер топтарының тарихи тұрғыдан қалыптасуы; организмнің тарихи дамуы немесе органикалық дүниенің, әр түрлі тип, класс, отряд, туыс және түрлердің эволюциясы. Жеке-дара даму — онтогенез бен тарихи даму — филогенез тірі табиғаттың біртұтас дамуының ажырамас қырлары болып саналады және бір-біріне өзара әсер етеді. Филогения — организмнің тарихи дамуы. Филогенез терминін 1866 ж. неміс биологі Э. Геккель (1834 — 1919) ұсынады. Филогенез процесін және оның заңдылықтарын филогенетика зерттейді. Филогенездік зерттеулердің мақсаты — жануарлардың, өсімдіктердің, микроорганизмдердің эволюциялық өзгерістері негізінде олардың тегін және организмдер арасындағы туыстық байланыстарды анықтау. Ағылшын эволюцияшысы В. Гарстанг филогенезді ұрпақтан-ұрпаққа өтетін олтогенез деген пікір айтты. Бұл пікірді орыс биологі И. И. Шмальгаузен (1884 -1963) дамытып, филогенезді онтогенездің белгілі қатары деп атады

Онтогенездегі бластомерлер арасындағы айырмашылықтардың пайда болуын жіне іртүрлі клеткалардың ұлпалардың пайда болуын , әртүрлі мүшелердің пайда боуын және іртүрлі мүшелердің түзілуін дифференциация процесі деп атайды. Диференциация – дамып келе жатқан организмнің құрылыстық , биохимиялық өзгнерісі.

Гендік реттелудің цитодифференцировкасы Цитодифференцировка процесінің негізінде гендік экспрессияның реттелуі жатыр деп саналады. Осы реттелудің нәтижесінде клеткалық белоктардың құрамы өзгереді. Мұндай бөлек полипептидтердің және олардың топтарының құрамынының пайда болуы , жойылуы , үлкеюін қосатын мұндай өзгерістердің алуантүрлілігі гендік экспрессияның реттелуінің актілерінің алуантүрлілігімен сипатталады. Бұларды мынадай етіп систематикалауға болады.
 А) Басқасымен байланысты емес жеке гендердің экспрессиясының инициациясы немесе тоқталуы ; Гендік")
1) А) Басқасымен байланысты емес жеке гендердің экспрессиясының инициациясы немесе тоқталуы ; Гендік экспрессияның координацияланбаған өзгерістері : Б) Жеке гендердің экспресиясын күшейту немесе әлсірету ; 2) Гендік экспрессиясының координацияланған өзгерістері : А)Гендердің топтарының біруақыттағы инициациясы немесе экспрессиясының тоқталуы ; Б) Өзге геннің немесе топтарының экспресиясының инициациясы кезінде бір немесе бірнеше гендердің экспресиясының тоқталуы ; В)Гендер топтарының экспресиясының біруақыттағы тіке пропорционалды өзгерісі ; Г) 2 геннің немесе гендік топтардың экспресиясының біруақыттағы кері пропорционалды өзгерісі ; Д) Гендік белсенділіктің пропорционалды емес координацияланған өзгерісі ; Е) Гендік өнімдердің санының қатынасын анықтайтын мықты координация ; 3)Гендік экспресиялардың « координацияланған және координацияланбаған ) уақыттағы өзгерістер ; 4)Гендік экспресияның ұзақ уақыттағы өзгерістері ( координация және координацияланбаған ); қысқа

Қазіргі таңдағы цитодифференцировканың негізгі механизмі болып дифференциальді гендердің экспресиясы саналады. Гендердің дифференциальді экспресиясының реттелуінің деңгейі ген полипептид – белгі бағытындағы информацияның реализация этаптарына байланысты және клеткаішілік процестерді ғана емес, сонымен қатар ұлпалық және ағзалық процестерді де қамтиды. Клетка дифференциациясына тән ерекшелігі ол қайтымсыз сол немесе соған ұқсас клеткалардың типтеріне алып баруымен сипатталады. Бұл процестің детерминация деген атауы бар және генетикалық қадағалануда , тирозинкиназа рецепторы арқылы пептидтік өсімділік факторларымен іске асатын сигналдардың негізінде клеткалардың дифференциация және детерминациясымен әсерлесіп , реттеледі

Гендердің дифференциальді экспресиясының реттелуінің деңгейі ген - полипептид – белгі бағытындағы информацияның реализация этаптарына байланысты және клеткаішілік процестерді ғана емес, сонымен қатар ұлпалық және ағзалық процестерді де қамтиды. Клетка дифференциациясына тән ерекшелігі ол қайтымсыз сол немесе соған ұқсас клеткалардың типтеріне алып баруымен сипатталады. Бұл процестің детерминация деген атауы бар және генетикалық қадағалануда , тирозинкиназа рецепторы арқылы пептидтік өсімділік факторларымен іске асатын сигналдардың негізінде клеткалардың дифференциация және детерминациясымен әсерлесіп , реттеледі. Мүмкін , осындай жүйелер көп шығар. Бұлшықеттің және нерв клеткаларының дифференциациясы бір немесе бірнеше тирозинкиназды рецепторлар арқлы әсер ететін өз алдына мембранды клеткалар болып келетін нейрорегуландармен реттеледі.

Эмбриональді даму периодындағы гендердің белсенділігінің регуляциясы да маңызды роль атқарады. Гендердің дифференциациясы әр түрлі уақытта әсер етеді деп есептеледі. Бұл м. РНК сы әр түрлі және әр түрлі дифференциацияланған клеткалардың транскрипциасындағы гендердің репрессиясы және дерепрессисында орын алады. Мысалы , теңіз кірпісінің бластоцистасының РНК-ғы транскиьирленетін гендердің саны – 10% ға тең , ал атжалмандардың бауырының клеткаларындағы саны да – 10 % ға тең , ірі мүйізді жануарлардың тимусының клеткаларындағы саны – 15 % ға тең.

Гендердің транскрипциасын қадағалағанда гистонды белоктар қатысады деген болжам бар. Осы болжам бойынша келесі мәліметтерге сүйенуге болады. Фазадағы клеткалардың хроматині in vitro жүйесінде транскибирленгенде гистонды м. РНК ғана синтезделеді , оның артынша гистондар да синтезделеді. Керісінше О фазасының клеткаларының хроматинін қолданса , онда ешқандай м. РНК синтезделмейді. G 1 фазасының хроматинінен гистонды емес белоктар ажырап және орнына гистонды емес хромосомды белоктар келсе ( S фазасында синтезделеді ) , онда мұндай хроматиннің транскрипцияснан кейін in vitro гистонды м. РНк синтезделеді. Гистонды емес белоктардың табиғаты G фазасының клеткаларыннан болса , онда ешқандай гистонды м. РНК синтезделмейді. Бұл нәтижелер хроматиннің құрамындағы гистонды емес белоктар гистонды кодтайтын гендердің транскрипциясының мүмкіндіктерін анықтайды. Сондықтан да гистонды емес хромасомды белоктар эукариоттардың гендерінің экспресиясында және реттелуіндегі маңызды роль атқарады деп саналады.

Қолданылған әдебиеттер : 1. Воронин Е. С. , Тихонов И. В. Биотехнология: Учебник для вузов – М. : Де Ли принт, 2006. – 521 с. 2. Биотехнология /Т. Г. Волова. – Новосибирск: Из-во СОРА, 1999 – 252 с. 3. Дебабов В. Г. Генная инженерия в производстве БАВ – М. : Де Ли принт, 2009 – 391 с. 4. Беккер М. Е. и др. Биотехнология – М. : Агропромиздат, 1990 — 420 с.
ген.ppt