

Лекция 2. Строение нуклеиновых кислот ДНК.ppt
- Количество слайдов: 57
 Строение нуклеиновых кислот Лекция 2. ДНК
Строение нуклеиновых кислот Лекция 2. ДНК
 Вся информация о строении и функционировании любого живого организма находится в закодированном виде в его генетическом материале, основу которого составляет ДНК. ИСТОРИЧЕСКАЯ СПРАВКА. В 1868 швейцарский патологоанатом Иоган Фридрих Мишер (Johan Friedrich Miescher, 1811– 1887) выделил из клеточного ядра новое вещество, которое он назвал нуклеином (лат. nucleus – ядро), о чем сообщил в 1869 (подробные данные были опубликованы в 1890, уже после смерти Мишера).
Вся информация о строении и функционировании любого живого организма находится в закодированном виде в его генетическом материале, основу которого составляет ДНК. ИСТОРИЧЕСКАЯ СПРАВКА. В 1868 швейцарский патологоанатом Иоган Фридрих Мишер (Johan Friedrich Miescher, 1811– 1887) выделил из клеточного ядра новое вещество, которое он назвал нуклеином (лат. nucleus – ядро), о чем сообщил в 1869 (подробные данные были опубликованы в 1890, уже после смерти Мишера).
 Нуклеиновые кислоты – нерегулярные полимеры, с мономерной единицей – нуклеотидом, состоящим из азотистого основания, пятиуглеродного сахара (пентозы) и фосфатной группы. Строение нуклеотида Фосфатная группа присоединена к 5’-атому углерода моносахаридного остатка, а основание – к 1’-атому
Нуклеиновые кислоты – нерегулярные полимеры, с мономерной единицей – нуклеотидом, состоящим из азотистого основания, пятиуглеродного сахара (пентозы) и фосфатной группы. Строение нуклеотида Фосфатная группа присоединена к 5’-атому углерода моносахаридного остатка, а основание – к 1’-атому
 – содержат два гетероцикла и пиримидиновые (тимин и") Азотистые основания. пуриновые (аденин и гуанин) – содержат два гетероцикла и пиримидиновые (тимин и урацил) – содержат один гетероци
Азотистые основания. пуриновые (аденин и гуанин) – содержат два гетероцикла и пиримидиновые (тимин и урацил) – содержат один гетероци
 Фосфодиэфирная связь Связи в одноцепочечной ДНК 5’-углеродная группа атома одного нуклеотида связана с 3’-ОН-группой дезоксирибозы соседнего нуклеотида. На одном конце полинуклеотидной цепи находится 3’-ОН-группа (3’конец), а на другом – 5’фосфатная группа (5’конец). Азотистые основания не принимают участия в соединении нуклеотидов одной цепи.
Фосфодиэфирная связь Связи в одноцепочечной ДНК 5’-углеродная группа атома одного нуклеотида связана с 3’-ОН-группой дезоксирибозы соседнего нуклеотида. На одном конце полинуклеотидной цепи находится 3’-ОН-группа (3’конец), а на другом – 5’фосфатная группа (5’конец). Азотистые основания не принимают участия в соединении нуклеотидов одной цепи.
 водородная связь Образуется между комплементрными парами оснований противоположный цепей. Пара А-Т стабилизируется двумя водородными связями, а Г-Ц – тремя. Длина ДНК измеряется числом комплементарных пар нуклеотидов. Структура ДНК Например, двойная спираль ДНК, соответствующая 1 хромосоме, составляет 263 м. п. н.
водородная связь Образуется между комплементрными парами оснований противоположный цепей. Пара А-Т стабилизируется двумя водородными связями, а Г-Ц – тремя. Длина ДНК измеряется числом комплементарных пар нуклеотидов. Структура ДНК Например, двойная спираль ДНК, соответствующая 1 хромосоме, составляет 263 м. п. н.
 Характеристики ДНК 1. Нерегулярность Существует регулярный сахарофосфатный остов, к которому присоединены азотистые основания. Их чередование нерегулярно.
Характеристики ДНК 1. Нерегулярность Существует регулярный сахарофосфатный остов, к которому присоединены азотистые основания. Их чередование нерегулярно.
 Характеристики ДНК 2. Антипараллельность ДНК состоит из двух полинуклеотидных цепей, ориентированных антипараллельно. 3`-конец одной расположен напротив 5`-конца другой.
Характеристики ДНК 2. Антипараллельность ДНК состоит из двух полинуклеотидных цепей, ориентированных антипараллельно. 3`-конец одной расположен напротив 5`-конца другой.
 Характеристики ДНК 3. Комплементарность Каждому азотистому основанию одной цепи соответствует строго определенное азотистое основание другой цепи. Соответствие задается химией.
Характеристики ДНК 3. Комплементарность Каждому азотистому основанию одной цепи соответствует строго определенное азотистое основание другой цепи. Соответствие задается химией.
 Характеристики ДНК 4. Регулярная вторичная структура Две комплементарные, антипараллельно расположенные полинуклеотидные цепи образуют правые спирали с общей осью
Характеристики ДНК 4. Регулярная вторичная структура Две комплементарные, антипараллельно расположенные полинуклеотидные цепи образуют правые спирали с общей осью
 Формы двойной спирали ДНК В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль ДНК в живых организмах существует в разных формах. На рисунке представлены формы A, B и Z (слева направо)
Формы двойной спирали ДНК В зависимости от концентрации ионов и нуклеотидного состава молекулы, двойная спираль ДНК в живых организмах существует в разных формах. На рисунке представлены формы A, B и Z (слева направо)
 м на") правозакрученные спирали Формы двойной спирали ДНК В-форма -10 Å - Ангстрем (10) м на один виток спирали (3. 4 нм) приходится 10 комплементарных пар. Диаметр спирали 20Å, причем пуриновый нуклеотид занимает 12Å , а пиримидиновый - 8Å.
правозакрученные спирали Формы двойной спирали ДНК В-форма -10 Å - Ангстрем (10) м на один виток спирали (3. 4 нм) приходится 10 комплементарных пар. Диаметр спирали 20Å, причем пуриновый нуклеотид занимает 12Å , а пиримидиновый - 8Å.
 м 11") правозакрученные спирали Формы двойной спирали ДНК А-форма -10 Å - Ангстрем (10) м 11 пар азотистых оснований на виток. Шаг спирали равен приблизительно 2. 8 нм.
правозакрученные спирали Формы двойной спирали ДНК А-форма -10 Å - Ангстрем (10) м 11 пар азотистых оснований на виток. Шаг спирали равен приблизительно 2. 8 нм.
 м шаг") правозакрученные спирали Формы двойной спирали ДНК С-форма -10 Å - Ангстрем (10) м шаг спирали 31Å, 9. 3 пар оснований на виток.
правозакрученные спирали Формы двойной спирали ДНК С-форма -10 Å - Ангстрем (10) м шаг спирали 31Å, 9. 3 пар оснований на виток.
") левозакрученная спираль Формы двойной спирали ДНК Z –форма «зигзаг» -10 Å - Ангстрем (10) м Высота витка в Zформе -44. 5 Å, на виток приходится 12 пар нуклеотидов.
левозакрученная спираль Формы двойной спирали ДНК Z –форма «зигзаг» -10 Å - Ангстрем (10) м Высота витка в Zформе -44. 5 Å, на виток приходится 12 пар нуклеотидов.
 лишена взаимозакрученности в двойную спираль.") Формы двойной спирали ДНК Форма SBS (side by side) лишена взаимозакрученности в двойную спираль. Важна для биосинтеза ДНК
Формы двойной спирали ДНК Форма SBS (side by side) лишена взаимозакрученности в двойную спираль. Важна для биосинтеза ДНК
 Функции ДНК n ДНК является носителем генетической информации. Функция обеспечивается фактом существования генетического кода. n ДНК отвечает за воспроизведение и передачу генетической информации в поколениях клеток и организмов. Функция обеспечивается процессом репликации.
Функции ДНК n ДНК является носителем генетической информации. Функция обеспечивается фактом существования генетического кода. n ДНК отвечает за воспроизведение и передачу генетической информации в поколениях клеток и организмов. Функция обеспечивается процессом репликации.
 Типы переноса информации в клетках Общий перенос – процесс, который обязательно осуществляется в клетках любого типа Специальный перенос – процесс осуществляется в клетках только при наличии особенных условий. Запрещенный перенос – тип переноса информации, который никогда не наблюдался ни при каких условиях. ДНК РНК БЕЛОК
Типы переноса информации в клетках Общий перенос – процесс, который обязательно осуществляется в клетках любого типа Специальный перенос – процесс осуществляется в клетках только при наличии особенных условий. Запрещенный перенос – тип переноса информации, который никогда не наблюдался ни при каких условиях. ДНК РНК БЕЛОК
 Репликация ДНК процесс, обеспечивающий перенос информации с ДНК на ДНК, то есть ДНК-копирование. Принципы репликации: 1. Комплементарность. Энергия, необходимая 2. Антипараллельность. для синтеза, высвобождается при отщеплении 3. Униполярность. пирофосфата, а катали 4. Потребность в зируют реакцию особые затравке. ферменты – ДНК 5. Прерывистость. полимеразы. 6. Полуконсервативность
Репликация ДНК процесс, обеспечивающий перенос информации с ДНК на ДНК, то есть ДНК-копирование. Принципы репликации: 1. Комплементарность. Энергия, необходимая 2. Антипараллельность. для синтеза, высвобождается при отщеплении 3. Униполярность. пирофосфата, а катали 4. Потребность в зируют реакцию особые затравке. ферменты – ДНК 5. Прерывистость. полимеразы. 6. Полуконсервативность
 Понятие о матрице и затравке Матрица для синтеза новых цепей - одноцепочечная ДНК. Затравка – 3'-гидроксильный конец двуцепочечной ДНК, причем он должен быть спарен с матрицей.
Понятие о матрице и затравке Матрица для синтеза новых цепей - одноцепочечная ДНК. Затравка – 3'-гидроксильный конец двуцепочечной ДНК, причем он должен быть спарен с матрицей.
 есть 4 шпильки, они опознаются РНКполимеразой.") прокариоты Образование РНК-затравки В ori районе (начало репликации) есть 4 шпильки, они опознаются РНКполимеразой. Вторая шпилька используется в качестве матрицы для образования РНКзатравки 3'-конец которой используется для репликации ДНК полимеразой III.
прокариоты Образование РНК-затравки В ori районе (начало репликации) есть 4 шпильки, они опознаются РНКполимеразой. Вторая шпилька используется в качестве матрицы для образования РНКзатравки 3'-конец которой используется для репликации ДНК полимеразой III.
 Факторы репликации ДНКполимеразы Топоизомеразы Геликазы Белки Альбертса
Факторы репликации ДНКполимеразы Топоизомеразы Геликазы Белки Альбертса
 ДНК-полимеразы К репликации имеют отношение ДНК-полимеразы I и III. Полимераза III является репликазой, т. е. синтезирует in vivo новые цепи ДНК. У ДНК-полимеразы I вспомогательная, репаративная функция. ДНК-полимераза II имеет отношение лишь к репарации
ДНК-полимеразы К репликации имеют отношение ДНК-полимеразы I и III. Полимераза III является репликазой, т. е. синтезирует in vivo новые цепи ДНК. У ДНК-полимеразы I вспомогательная, репаративная функция. ДНК-полимераза II имеет отношение лишь к репарации
 Топоизомеразы - ферменты, изменяющие топологию ДНК классы релаксазы гиразы уменьшают число зацеплений увеличивают число зацеплений
Топоизомеразы - ферменты, изменяющие топологию ДНК классы релаксазы гиразы уменьшают число зацеплений увеличивают число зацеплений
 Геликазы У E. coli есть четыре геликазы: геликаза I, геликаза III и rep-белок. Rep-геликаза используется при репликации одноцепочечных ДНК-содержащих фагов. Для репликации бактериальной ДНК она не нужна. Геликазы различаются требованиями к размеру посадочной площадки, на которую они садятся для начала движения.
Геликазы У E. coli есть четыре геликазы: геликаза I, геликаза III и rep-белок. Rep-геликаза используется при репликации одноцепочечных ДНК-содержащих фагов. Для репликации бактериальной ДНК она не нужна. Геликазы различаются требованиями к размеру посадочной площадки, на которую они садятся для начала движения.
 Белки Альбертса Эти белки содержат кластер положительно заряженных аминокислотных остатков,") SSB (single strand bind) Белки Альбертса Эти белки содержат кластер положительно заряженных аминокислотных остатков, но общий заряд белка отрицателен и связываются только с одноцепочечной ДНК SSB удерживают матричные цепи ДНК в репликативной вилке в одноцепочечном состоянии и защищают одноцепочечную ДНК от действия нуклеаз.
SSB (single strand bind) Белки Альбертса Эти белки содержат кластер положительно заряженных аминокислотных остатков, но общий заряд белка отрицателен и связываются только с одноцепочечной ДНК SSB удерживают матричные цепи ДНК в репликативной вилке в одноцепочечном состоянии и защищают одноцепочечную ДНК от действия нуклеаз.
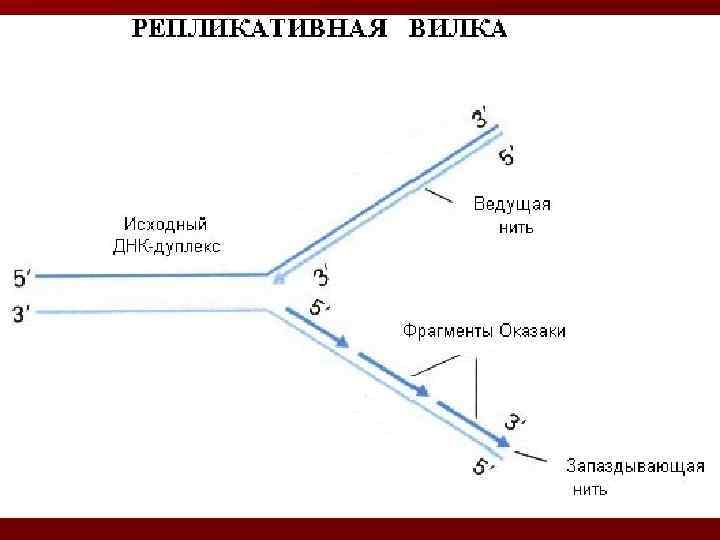
 при использовании") Этапы репликации 1. Репликация начинается с расплетания цепей ДНК ГЕЛИКАЗАМИ (или Rep-протеином) при использовании энергию АТФ. Скорость расплетания составляет около 6000 мин-1.
Этапы репликации 1. Репликация начинается с расплетания цепей ДНК ГЕЛИКАЗАМИ (или Rep-протеином) при использовании энергию АТФ. Скорость расплетания составляет около 6000 мин-1.
 2. SSB-белки присоединяются к комплементарным цепям, удерживая их от ассоциации. По мере продвижения репликационной вилки SSBпротеины передвигаются по цепи, диссоциируя с одного места и присоединяясь на другом. Этот процесс не требует затрат энергии АТФ.
2. SSB-белки присоединяются к комплементарным цепям, удерживая их от ассоциации. По мере продвижения репликационной вилки SSBпротеины передвигаются по цепи, диссоциируя с одного места и присоединяясь на другом. Этот процесс не требует затрат энергии АТФ.
 3. Синтезируется праймер-затравка. Наличие затравки является необходимым условием функционирования ДНК-полимераз (как и наличие комплементарной цепи). В качестве затравки выступают маленькие отрезки молекул РНК, которые синтезируются при помощи фермента ПРИМАЗЫ.
3. Синтезируется праймер-затравка. Наличие затравки является необходимым условием функционирования ДНК-полимераз (как и наличие комплементарной цепи). В качестве затравки выступают маленькие отрезки молекул РНК, которые синтезируются при помощи фермента ПРИМАЗЫ.
 4. Элонгация. Когда синтез на одном из фрагментов Оказаки достигает праймера другого фрагмента, РНК-овый праймер удаляется имеющейся у полимераз 5'-3' полимеразной активностью и достраивается дезоксирибонуклеотидами. После этого сахарофосфатный остов между фрагментами сшивается ковалентной связью при помощи фермента ДНК-лигазы.
4. Элонгация. Когда синтез на одном из фрагментов Оказаки достигает праймера другого фрагмента, РНК-овый праймер удаляется имеющейся у полимераз 5'-3' полимеразной активностью и достраивается дезоксирибонуклеотидами. После этого сахарофосфатный остов между фрагментами сшивается ковалентной связью при помощи фермента ДНК-лигазы.
 Репликация концов ДНК хромосом эукариот Удаление РНК-праймеров после завершения синтеза линейных ДНК и заделывание образующихся между фрагментами брешей нуклеотидами ДНК приводит к тому, что дочерние цепи ДНК оказываются короче материнских на размер первого РНКпраймера (10 -20 нукл. ).
Репликация концов ДНК хромосом эукариот Удаление РНК-праймеров после завершения синтеза линейных ДНК и заделывание образующихся между фрагментами брешей нуклеотидами ДНК приводит к тому, что дочерние цепи ДНК оказываются короче материнских на размер первого РНКпраймера (10 -20 нукл. ).
 1. Выступающие 3'концы материнских цепей узнаются ферментом теломеразой - содержащим помимо белковой части еще и РНК, выполняющую роль матрицы для наращивания ДНК повторами. 2. Теломераза последовательно наращивает материнские цепи, используя 3'-концы в качестве затравок. 3. Образующиеся длинные одноцепочечные концы служат матрицами для синтеза дочерних цепей традиционным репликативным механизмом.
1. Выступающие 3'концы материнских цепей узнаются ферментом теломеразой - содержащим помимо белковой части еще и РНК, выполняющую роль матрицы для наращивания ДНК повторами. 2. Теломераза последовательно наращивает материнские цепи, используя 3'-концы в качестве затравок. 3. Образующиеся длинные одноцепочечные концы служат матрицами для синтеза дочерних цепей традиционным репликативным механизмом.
 Хромосомы соматических клеток фланкированы многократно повторенными гексамерами TTAГГГ, общая длина районов с повторами может достигать 10 т. п. н. В комплексе со специфическими белками такие тандемные повторы образуют теломеры. При репликации происходит укорочение теломер в среднем на 50 пар нуклеотидов.
Хромосомы соматических клеток фланкированы многократно повторенными гексамерами TTAГГГ, общая длина районов с повторами может достигать 10 т. п. н. В комплексе со специфическими белками такие тандемные повторы образуют теломеры. При репликации происходит укорочение теломер в среднем на 50 пар нуклеотидов.
 Некоторые характеристики репликации Организм Количество репликонов Средний размер репликона, тыс. п. н. Скорость движения репликативной вилки п. н. /мин. E. сoli 1 4200 50000 Дрожжи 500 40 3600 Дрозофила 3500 40 2600 15000 200 500 Ксенопус (лягушка)
Некоторые характеристики репликации Организм Количество репликонов Средний размер репликона, тыс. п. н. Скорость движения репликативной вилки п. н. /мин. E. сoli 1 4200 50000 Дрожжи 500 40 3600 Дрозофила 3500 40 2600 15000 200 500 Ксенопус (лягушка)
 Особенности репликация у эукариот У эукариот ДНК закрепляется белками в нескольких местах на ядерной мембране. На каждом отдельном участке работает топоизомераза. Сколько участков, столько и ori
Особенности репликация у эукариот У эукариот ДНК закрепляется белками в нескольких местах на ядерной мембране. На каждом отдельном участке работает топоизомераза. Сколько участков, столько и ori
 Скорость синтеза ДНК Прокариот 1000 нуклеотидов в секунду, ошибки репликации возникают с частотой 1 * 108 - 1 * 10 -10 нуклеотид. Эукариот 100 нуклеотидов в секунду, так как вновь синтезированные нуклеотиды связываются с гистоновыми белками. Ошибки репликации у эукариот на порядок ниже.
Скорость синтеза ДНК Прокариот 1000 нуклеотидов в секунду, ошибки репликации возникают с частотой 1 * 108 - 1 * 10 -10 нуклеотид. Эукариот 100 нуклеотидов в секунду, так как вновь синтезированные нуклеотиды связываются с гистоновыми белками. Ошибки репликации у эукариот на порядок ниже.
 Моно- и полирепликонные системы
Моно- и полирепликонные системы
 ДНКполимеразы типы Участие в синтезе ДНК Прокариоты 5 типов:") Репликация ДНК (отличие ферментных систем) ДНКполимеразы типы Участие в синтезе ДНК Прокариоты 5 типов: I – V тип обе цепи ДНК синтезируются ДНКполимеразой III Эукариоты 6 типов: альфа, бета, дельта, эпсилон, гамма, дзета альфа – инициация, дельта – элонгация ведущей, эпсилон – отстающей цепи
Репликация ДНК (отличие ферментных систем) ДНКполимеразы типы Участие в синтезе ДНК Прокариоты 5 типов: I – V тип обе цепи ДНК синтезируются ДНКполимеразой III Эукариоты 6 типов: альфа, бета, дельта, эпсилон, гамма, дзета альфа – инициация, дельта – элонгация ведущей, эпсилон – отстающей цепи
 ОШИБКИ РЕПЛИКАЦИИ РЕПАРАЦИЯ
ОШИБКИ РЕПЛИКАЦИИ РЕПАРАЦИЯ
 «Способность ошибаться заложена в структуре ДНК-полимеразы»
«Способность ошибаться заложена в структуре ДНК-полимеразы»
 Вероятность ошибок для ферментов вирусов, про- и эукариот Объект Вероятность замены пару оснований E. coli 2 х10 -10 Дрозофила 5 х10 -11 Фаг Т 4 2 х10 -8 Разницу связывают со скоростью работы фермента. Чем медленней, тем точнее! на
Вероятность ошибок для ферментов вирусов, про- и эукариот Объект Вероятность замены пару оснований E. coli 2 х10 -10 Дрозофила 5 х10 -11 Фаг Т 4 2 х10 -8 Разницу связывают со скоростью работы фермента. Чем медленней, тем точнее! на
 РЕПАРАЦИЯ ДНК n способность исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК или в результате воздействия физическими или химическими агентами. n Осуществляется специальными ферментными системами клетки.
РЕПАРАЦИЯ ДНК n способность исправлять химические повреждения и разрывы в молекулах ДНК, повреждённой при нормальном биосинтезе ДНК или в результате воздействия физическими или химическими агентами. n Осуществляется специальными ферментными системами клетки.
, лигирующая") ДНК-полимераза I (кольцеобразная структура, состоящая из нескольких одинаковых молекул белка, показанных разными цветами), лигирующая повреждённую цепь ДНК
ДНК-полимераза I (кольцеобразная структура, состоящая из нескольких одинаковых молекул белка, показанных разными цветами), лигирующая повреждённую цепь ДНК
 Этапы проверки n Первичный отбор нуклеотидов идет по принципу n Редактирующий отбор. Его проводят все n Исправление ошибок в уже синтезированной ДНК. Этим занимаются ферменты репарации комплементарности. Способностью к этому виду отбора обладают все ДНК-полимеразы благодаря полимеризационной 5‘ - 3' активности. полимеразы благодаря экзонуклеазной активности 3‘ - 5‘
Этапы проверки n Первичный отбор нуклеотидов идет по принципу n Редактирующий отбор. Его проводят все n Исправление ошибок в уже синтезированной ДНК. Этим занимаются ферменты репарации комплементарности. Способностью к этому виду отбора обладают все ДНК-полимеразы благодаря полимеризационной 5‘ - 3' активности. полимеразы благодаря экзонуклеазной активности 3‘ - 5‘
 – вырезание: поврежденных нуклеотидов ( NER -") Механизмы репарации n эксцизионная репарация ДНК (NER) – вырезание: поврежденных нуклеотидов ( NER - nucleotide excision repair ) 2. азотистых оснований ( BER - base excision repair). 1.
Механизмы репарации n эксцизионная репарация ДНК (NER) – вырезание: поврежденных нуклеотидов ( NER - nucleotide excision repair ) 2. азотистых оснований ( BER - base excision repair). 1.
 – исправление ошибок спаривания, возникающих в результате") Механизмы репарации n мисмэтч-репарация (mismatch repair, MMR) – исправление ошибок спаривания, возникающих в результате нарушения комплементарности пар AT или GC в дочерней цепи при включении в них некомплементарных нуклеотидов.
Механизмы репарации n мисмэтч-репарация (mismatch repair, MMR) – исправление ошибок спаривания, возникающих в результате нарушения комплементарности пар AT или GC в дочерней цепи при включении в них некомплементарных нуклеотидов.
 Основные репарабельные повреждения в ДНК и принципы их устранения 1. Апуринизация. Каждая соматическая клетка теряет за сутки около 10000 пуринов и пиримидинов. В ДНК образуются АП-сайты. Причины апуринизации: изменение р. Н, ионизирующее излучение, повышение температуры и т. д.
Основные репарабельные повреждения в ДНК и принципы их устранения 1. Апуринизация. Каждая соматическая клетка теряет за сутки около 10000 пуринов и пиримидинов. В ДНК образуются АП-сайты. Причины апуринизации: изменение р. Н, ионизирующее излучение, повышение температуры и т. д.
 Возникновение АР-сайтов. Разрывается N-гликозидная связь между пуриновым основанием и дезоксирибозой. AP-сайты: 1. химически нестабильны и на их месте могут возникать одноцепочечные разрывы ДНК. 2. Блокируют репликацию
Возникновение АР-сайтов. Разрывается N-гликозидная связь между пуриновым основанием и дезоксирибозой. AP-сайты: 1. химически нестабильны и на их месте могут возникать одноцепочечные разрывы ДНК. 2. Блокируют репликацию
 Механизмы репарации АР-сайтов n Эксцизионный – апуриновая эндонуклеаза узнает в ДНК апуриновый участок, далее вступают экзонуклеазы ДНК, полимеразы и лигазы. n Репуринизация. Существует фермент репуриназа, катализирующий прямое включение пуриновых оснований в апуриновые участки
Механизмы репарации АР-сайтов n Эксцизионный – апуриновая эндонуклеаза узнает в ДНК апуриновый участок, далее вступают экзонуклеазы ДНК, полимеразы и лигазы. n Репуринизация. Существует фермент репуриназа, катализирующий прямое включение пуриновых оснований в апуриновые участки
 2. Дезаминирование – процесс удаления аминогруппы Аденин превращается в гипоксантин, который образует две водородные связи с цитозином. Гуанин превращается в ксантин, который образует водородные связи с тимином. При дезаминировании цитозина образуется урацил. Тимин не может быть дезаминирован (единственный в ДНК).
2. Дезаминирование – процесс удаления аминогруппы Аденин превращается в гипоксантин, который образует две водородные связи с цитозином. Гуанин превращается в ксантин, который образует водородные связи с тимином. При дезаминировании цитозина образуется урацил. Тимин не может быть дезаминирован (единственный в ДНК).
 Механизи репарации дезаминирования n n N-гликозилаза - узнает дезамини-рованное основание, разрывает N-гликозидную связь и удаляет неправильное основание. После этого эндонуклеаза вносит одноцепочечный разрыв, а фосфодиэстераза отщепляет от ДНК сахарофосфатную группу, к которой не присоединено основание. Появляется брешь размером в один нуклеотид. У E. coli она заделывается ДНК-полимеразой I, а лигаза сшивает концы ДНК. У эукариот брешь заделывает ДНК-полимераза.
Механизи репарации дезаминирования n n N-гликозилаза - узнает дезамини-рованное основание, разрывает N-гликозидную связь и удаляет неправильное основание. После этого эндонуклеаза вносит одноцепочечный разрыв, а фосфодиэстераза отщепляет от ДНК сахарофосфатную группу, к которой не присоединено основание. Появляется брешь размером в один нуклеотид. У E. coli она заделывается ДНК-полимеразой I, а лигаза сшивает концы ДНК. У эукариот брешь заделывает ДНК-полимераза.
 3. Тиминовые димеры. Под действием ультрафиолетого света происходит ковалентное сшивание рядом стоящих пиримидинов. При сшивании тиминов образуется циклобутановое производное, блокирующее репликацию.
3. Тиминовые димеры. Под действием ультрафиолетого света происходит ковалентное сшивание рядом стоящих пиримидинов. При сшивании тиминов образуется циклобутановое производное, блокирующее репликацию.
 а - тиминовый димер циклобутанового типа; б - пиримидиновый димер
а - тиминовый димер циклобутанового типа; б - пиримидиновый димер
 Механизм репарации димеров у бактерий Фермент фотолиаза - узнает тиминовые димеры и образует с ними комплекс. При освещении видимым светом происходит активация фермента, циклобутановое кольцо разрывается, и вновь получаются два тимина. Этот процесс называется фотореактивацией
Механизм репарации димеров у бактерий Фермент фотолиаза - узнает тиминовые димеры и образует с ними комплекс. При освещении видимым светом происходит активация фермента, циклобутановое кольцо разрывается, и вновь получаются два тимина. Этот процесс называется фотореактивацией
 Стадии восстановления ДНК, в которой образовался циклобутановый димер. Участок с этой структурой, мешающей копированию ДНК, распознают и вырезают эндонуклеазы семейства ХР (ксеродермы). У человека работают 7 белков - продуктов генов семейства ХР (XPA-XPG). Застраивает образовавшуюся брешь в 29 нуклеотидов полимераза d (или e) при наличии фактора репликации (RFC) и белкапомощника (PCNA).
Стадии восстановления ДНК, в которой образовался циклобутановый димер. Участок с этой структурой, мешающей копированию ДНК, распознают и вырезают эндонуклеазы семейства ХР (ксеродермы). У человека работают 7 белков - продуктов генов семейства ХР (XPA-XPG). Застраивает образовавшуюся брешь в 29 нуклеотидов полимераза d (или e) при наличии фактора репликации (RFC) и белкапомощника (PCNA).
 И дезаминированные основания, и тиминовые димеры, кроме того, могут удаляться с помощью эксцизионной репарации: 1. инцизия - специфические эндонуклеазы производят одноцепочечные разрезы 2. эксцизия - затем происходит удаление нескольких нуклеотидов и заделывание бреши. У E. сoli заделыванием бреши занимается ДНК-полимераза I. Лигаза сшивает цепь. Она же ликвидирует одноцепочечные разрывы, возникающие при действии ионизирующей радиации.
И дезаминированные основания, и тиминовые димеры, кроме того, могут удаляться с помощью эксцизионной репарации: 1. инцизия - специфические эндонуклеазы производят одноцепочечные разрезы 2. эксцизия - затем происходит удаление нескольких нуклеотидов и заделывание бреши. У E. сoli заделыванием бреши занимается ДНК-полимераза I. Лигаза сшивает цепь. Она же ликвидирует одноцепочечные разрывы, возникающие при действии ионизирующей радиации.