Лекция 13-14 биологи.pptx
- Количество слайдов: 131


")
СООБЩЕСТВО многовидовое (community)
 консорция Трофическая группировка (гильдия, ассоциация простое сообщество) синонимы: биотоп, косная среда,")
Биотическая компонента редуцированный) консорция Трофическая группировка (гильдия, ассоциация простое сообщество) синонимы: биотоп, косная среда, окружающая среда С о о б щ е с т в о Абиотическая компонента Биоценоз (полный или синоним: живое вещество Видовая популяция биогеоценоз (по В. Н. Сукачеву), экосистема по А. Тэнсли (1935) экосистема по Ф. Эвансу (1956)
, консументов (животные) и редуцентов (детритофаги,")
Функциональное определение Биоценоз – группа популяций продуцентов (растений, бактерий), консументов (животные) и редуцентов (детритофаги, грибы и бактерии), приспособившихся к совместному обитанию в общем гомогенном пространстве
 и выровненности (evenness)")
Видовое разнообразие оценивается по видовому богатству (species richness) и выровненности (evenness)
 (Whittaker, Pianka) 2. Индексы разнообразия")
Способы оценки видового разнообразия 1. Кривые доминированияразнообразия (значимости) (Whittaker, Pianka) 2. Индексы разнообразия

/ln. N S/1000 S – числов видов Индекс видового разнообразия")
Индексы видового богатства S/N (S-1)/ln. N S/1000 S – числов видов Индекс видового разнообразия Симпсона (Simpson): D = 1/ Σ(Pi 2) i = 1, S Pi- доля i-го вида в общей численности или D = (1 – Σ(Pi 2)) E = D/ Dmax = D/ 1/(S(1/S)2 = D/S
")
Индекс разнообразия Шеннона (Shannon)
")
Показатели видового разнообразия тем выше: 1. Чем больше число видов – видовое богатство (S) и 2. Чем более равномерно соотношение численностей разных видов – т. е. выше выровненность (менее выражено доминирование немногих видов)

ВЫВОД: ЧИСЛО И СКОРОСТЬ ПРИРОСТА ВИДОВ В СООБЩЕСТВЕ ЗАВИСЯТ ОТ: • ХАРАКТЕРА СРЕДЫ (ИЗОЛИРОВАННОСТЬ, НЕОДНОРОДНОСТЬ) • МАСШТАБА ПРОСТРАНСТВА • РАЗМЕРОВ И ПОДВИЖНОСТИ ОРГАНИЗМОВ

Осмысление проблемы организации сообществ началось в 20 -е годы ХХ века Тогда были сформулированы ДВА ПОДХОДА:
 Концепция «сверхорганизма» Ф. Клеменса (Clements, 1916) (2) «индивидуалистическая» концепция Г. Глизона (Gleason, 1926)")
(1) Концепция «сверхорганизма» Ф. Клеменса (Clements, 1916) (2) «индивидуалистическая» концепция Г. Глизона (Gleason, 1926)
 Сообщество – совокупность разных организмов, обитающих в одном месте и образующих высокоинтегрированное целое,")
(1) Сообщество – совокупность разных организмов, обитающих в одном месте и образующих высокоинтегрированное целое, характеризующееся специфическим развитием, поведением, а возможно, и эволюцией
 Сообщество – совокупность организмов разных видов, попавших в одно место и нашедших там")
(2) Сообщество – совокупность организмов разных видов, попавших в одно место и нашедших там подходящие условия для обитания. «Целостность» сообщества – артефакт подхода исследователей
 и Э. Уилсон (Edward Wilson)")
Теория островной биогеографии Р. Мак Артура (Robert Mac. Arthur) и Э. Уилсон (Edward Wilson) (1967): Объяснение того, почему на каждом из островов, входящих в состав архипелага, обитает меньше видов, чем на равной по площади территории на материке

Для каждого острова существует точка равновесия, при которой скорость заселения и скорость вымирания совпадают (но видовой состав в равновесной точке может меняться)

Чем меньше остров, тем интенсивнее вымирание, и тем меньше равновесное количество видов

Чем ближе остров к материку, тем интенсивнее заселение, и тем больше равновесное количество видов


В теории островной биогеографии Макартура – Уилсона не предполагается расхождения видов по разным нишам. Сообщество формируется процессом расселения организмов, а не конкуренцией между уже прибывшими на остров видами Такие сообщества называют “dispersal assembled” в отличие от “niche assembled”

• От каких факторов зависит видовое богатство?
: ЗАКОН, ЗАГАДКА ИЛИ АРТЕФАКТ? А) Рукокрылые Нового Света Б)")
ШИРОТНЫЕ ГРАДИЕНТЫ РАЗНООБРАЗИЯ (А. Уоллес): ЗАКОН, ЗАГАДКА ИЛИ АРТЕФАКТ? А) Рукокрылые Нового Света Б) Морские беспозвоночные

 • ПЛОЩАДЬ (ГЕОМЕТРИЯ) • ЭНЕРГИЯ")
ПРИЧИНЫ И МЕХАНИЗМЫ широтного градиента видового богатства: (ОСНОВНЫЕ ГИПОТЕЗЫ) • ПЛОЩАДЬ (ГЕОМЕТРИЯ) • ЭНЕРГИЯ и ПРОДУКТИВНОСТЬ • СТАБИЛЬНОСТЬ И ПРЕДСКАЗУЕМОСТЬ КЛИМАТА • ГЕТЕРОГЕННОСТЬ СРЕДЫ • ЭВОЛЮЦИОННАЯ ЗРЕЛОСТЬ • СКОРОСТЬ ВИДООБРАЗОВАНИЯ И ВЫМИРАНИЯ • МЕЖВИДОВЫЕ ОТНОШЕНИЯ
 ГИПОТЕЗЫ: • Эффективного Эволюц. Времени: ЭЭВ = время эволюции × скорость")
БОЛЕЕ СЛОЖНЫЕ (КОМБИНИРОВАННЫЕ) ГИПОТЕЗЫ: • Эффективного Эволюц. Времени: ЭЭВ = время эволюции × скорость эволюции • Эффективной Продуктивности: ЭП = средняя продуктивность × стабильность (= продуктивность / VAR продуктивности)
 (А. И. Азовский, 2001) 1")
ДОСТОВЕРНОСТЬ ШИРОТНЫХ ГРАДИЕНТОВ РАЗНООБРАЗИЯ ДЛЯ РАЗНЫХ РАЗМЕРНЫХ ГРУПП (альфа-разнообразие) (А. И. Азовский, 2001) 1 - деревья, 2 – рептилии, 3 – рыбы, 4 - млекопитающие, 5 - амфибии, 6 - птицы, 7 - прочие сосудистые растения, 8 - макроводоросли, 9 - макрозообентос, 10 - насекомые, 11 - зоопланктон, 12 - мейобентос, 13 - простейшие, 14 – микроводоросли.
 3. Гетерогенность среды (разнообразие биотопа) 4.")
1. Эволюционное время 2. Стадия развития сообществ (сукцессии) 3. Гетерогенность среды (разнообразие биотопа) 4. Устойчивость и предсказуемость климата



5. Продуктивность сообщества


 сообщества")
Видовое разнообразие ? ? ? Продуктивность (биомасса) сообщества

Science 25 August 2000: Vol. 289. no. 5483, p. 1255 M. A. Huston, L. W. Aarssen, M. P. Austin, B. S. Cade, J. D. Fridley, E. Garnier, J. P. Grime, J. Hodgson, W. K. Lauenroth, K. Thompson, J. H. Vandermeer, D. A. Wardle No Consistent Effect of Plant Diversity on Productivity Hector et al. (1) reported on BIODEPTH, a major international experiment on the response of plant productivity to variation in the number of plant species. They found "an overall log-linear reduction of average aboveground biomass with loss of species, " leading to what the accompanying Perspective (2) described as "a rule of thumb--that each halving of diversity leads to a 10 to 20% reduction in productivity. . . We agree with Hector et al. that environmental conditions have a major effect on plant productivity and that overyielding does occur in some multispecies mixtures, particularly those containing nitrogen-fixing legumes. In light of our analysis of the data presented in their figure 2, however, we conclude that species richness per se has no statistically or biologically significant effect on plant productivity across the eight sites of the BIODEPTH experiment.




6. Конкуренция и хищничество

Схема взаимодействия механизмов организации биоценоза по Дж. Коннеллу: 1 - численность популяции; 2 - смертность, вызванная неблагоприятными абиотическими факторами среды обитания; 3 – смертность, обусловленная хищничеством; А – популяции, численность которых ограничивается неблагоприятными физическими факторами; Б - популяции, численность которых ограничивается интенсивным хищничеством

ЧТО ЗАЩИЩАЕТ РЕДКИЕ ВИДЫ ОТ ВЫМИРАНИЯ? В КАКОМ СЛУЧАЕ РЕДКИМ БЫТЬ ВЫГОДНЕЕ ЧЕМ МНОГОЧИСЛЕННЫМ?

Гипотеза Джанзена - Коннелла Воздействие специализированных животных фитофагов и патогенных микроорганизмов резко снижает выживаемость семян и проростков деревьев, если они находятся вблизи материнского растения. Для каждого отдельного растения шанс дорасти до зрелости выше, если оно располагается дальше от взрослых растений того же вида.

Выживаемость проростков Sebastiana longicuspis при разной плотности и поражении патогенными грибами (Thomas Bell, Robert P. Freckleton and Owen T. Lewis. Plant pathogens drive densitydependent seedling mortality in a tropical tree // Ecology Letters. 2006. V. 9. P. 569 -574)

Однако лучшая выживаемость редких видов тропических деревьев в сравнении с массовыми может объясняться не только гипотезой Джанзена - Коннела. Эффект может быть связан с некоторым разделением экологических ниш, в результате чего интенсивность межвидовой конкуренции оказывается ниже внутривидовой Эффект может объясняться положительным влиянием соседних растений в том случае, когда эти растения оказываются другого вида. Это так называемая «гипотеза облегчения» (facilitation)

Возможное разделение ниш: Nathan J. B. Kraft, Renato Valencia, David D. Ackerly Functional Traits and Niche-Based Tree Community Assembly in an Amazonian Forest // Science. 2008. Vol. 322. P. 580 -582 В Yasuni Forest в восточной части Эквадора. Здесь обнаружено максимальное известное на Земле видовое разнообразие деревьев. На модельном участке леса в 25 га (Yasuni Forest Dynamics Plot) - более 1123 видов деревьев (с диаметром ствола не менее 1 см на высоте груди человека). Все эти деревья (более 150 тыс. ) определены до вида, измерены, и нанесены на карту

Возможное разделение ниш: Nathan J. B. Kraft, Renato Valencia, David D. Ackerly // Science. 2008. Vol. 322. P. 580582 В качестве характеристик жизненных стратегий фигурировали: удельная поверхность листьев (площадь листьев, отнесенная к их массе), содержание в листьях азота, средний вес семени, максимальный диаметр на уровне груди (косвенный показатель максимальной высоты дерева), плотность древесины. Некоторые получены для всех 1123 видов деревьев, другие для большей части видов. Мерило пространственной близости деревьев попадание их в одну учетную площадку 20× 20 м. Нуль-гипотеза (нейтрализм) предполагала случайную комбинацию произрастающих вместе видов.
, цветом выделены значения «удельной (на единицу")
А. Показан рельеф местности (горизонтали через 2 м), цветом выделены значения «удельной (на единицу массы) площади листьев» . На «возвышенностях» и в «понижениях» преобладают деревья, различающиеся по удельной площади листьев, что указывает на «фильтрацию» , производимую средой (в долинах преобладают деревья с более тонкими листьями). B. Реальное распределение массы семян деревьев, произрастающих по соседству, то есть попавших на одну учетную площадку 20 × 20 м (сплошная линия — observed quadrat) и распределение, ожидаемое согласно нулевой гипотезе (пунктирная линия — null expectation), предполагающее независимое случайное распределение растений друг относительно друга. Видно, что для деревьев, оказавшихся рядом, наблюдается больший разброс значений этого признака, чем ожидаемый при случайной выборке. Соседствующие деревья разных видов в большей степени различаются по размеру семян, чем это предсказывается на основе случайного их распределения.

Каковы бы ни были конкретные процессы, происходящие в сообществе, очевидно, что вероятность выживания на ранних стадиях у редких видов выше, чем у массовых Эти результаты в принципе работают против гипотезы нейтральности (хотя сам Хаббел является одним из 34 авторов данной статьи )
")
7. Площадь местообитания (размер биотопа)

Может ли сообщество быть насыщено видами?

Существует ли внутренняя регуляция в биоценозах?

Есть ли связь между видовым разнообразием и устойчивостью сообществ?



Устойчивость сообщества не является прямой функцией числа видов. Число видов может расти, пока это не снижает эффективность использования сообществом потока вещества и энергии в данной точке

“Волчок жизни” по Н. Ф. Реймерсу: слева - для упрощённой модельной экосистемы Ю. Одума: люцерна - телятина - мальчик (с включением других консументов и редуцентов); справа - для биоценоза Земли в целом

Видовое разнообразие и устойчивость
 successions")
Сукцессии (развитие сообществ) successions
 (Т. А. Работнов, 1983)")
Сукцессии- необратимые, направленные и закономерные смены растительного покрова (фитоценозов) (Т. А. Работнов, 1983)
 конечные аутогенные конечные Гетеротрофные (деградационные) аллогенные циклические дигрессии")
Классификация сукцессий Автотрофные первичные вторичные (демутационные) конечные аутогенные конечные Гетеротрофные (деградационные) аллогенные циклические дигрессии конечные циклические длительные (циклические)
")
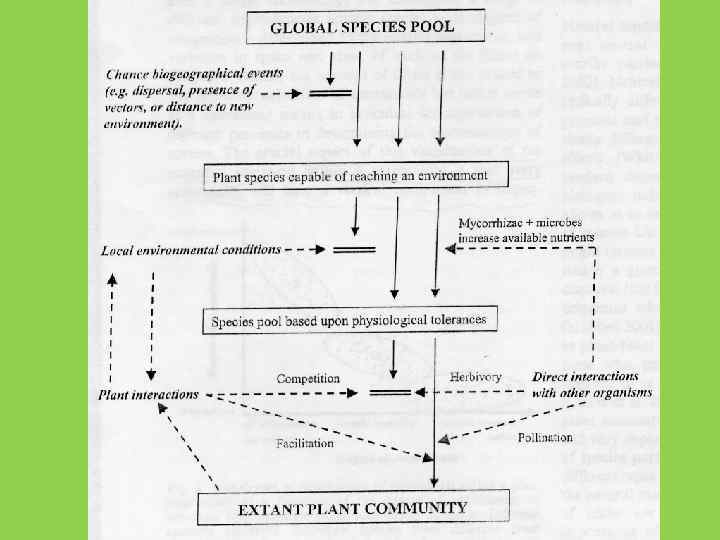
• Автотрофные сукцессии – процесс закономерного изменения видового состава сообществ (прежде всего, фитоценозов) в данном местообитании, вызванный однократным или постоянным действием абиотических или биотических факторов

Пример автотрофной демутационной аутогенной сукцессии-зарастание пашни 0 лет 3 лет 17 лет
 170 лет")
45 лет 100 лет Люри и др. , 2010 (ин-т географии РАН) 170 лет
 сукцессии")
Послепожарные (пирогенные) сукцессии

Нет ни одного сообщества на суше, которое не испытывало бы на себе действие пожаров! Пожары – один из важнейших факторов определяющих облик наземных сообществ.

ca brevifolia или дерево Джошуа


В ходе автотрофной сукцессии закономерно меняется как видовой состав, так и пространственная структура сообществ

При гетеротрофной сукцессии разлагается само местообитание – мертвое органическое вещество
 сукцессия")
Гетеротрофная (деградационная) сукцессия

Первичная сукцессия: отступление горного ледника
.")
Первичная сукцессия на склонах вулкана Этна (2 года спустя после извержения 2001).
, сложноцветных и бобовых (g. Astragalus).")
Зарастание вулканического шлака видами-пионерами семейств гвоздичных (g. Saponaria), сложноцветных и бобовых (g. Astragalus).

 Облегчение (facilitation) Толерантность (tolerance) Ингибирование (inhibition)")
Механизмы автотрофной cукцессии (Connell, Slatyer, 1977) Облегчение (facilitation) Толерантность (tolerance) Ингибирование (inhibition)
")
Механизмы автотрофной cукцессии (Connell, Slatyer, 1977)
 Pob=0 Pab")
No Poo Pao Pbo No Na Pаа Poа Pba=0 Na Nb (1) Pob=0 Pab Pbb Nb (2)
 имеет «марковское поведение»")
Модель Хорна (Н. Horn, 1975) имеет «марковское поведение»
 – процесс, при котором вероятность наступления определенного состояния")
Марковский процесс (А. А. Марков, 1920) – процесс, при котором вероятность наступления определенного состояния зависит только от текущего состояния и не зависит от пути, которым система пришла к текущему состоянию


 – стационарный видовой состав сообщества на конечной стадии автотрофной сукцессии")
Климакс (Whitford, 1901) – стационарный видовой состав сообщества на конечной стадии автотрофной сукцессии
 – серии завершаются одним")
Развитие понятия климакса • Теория моноклимакса Ф. Клементса (Clements, 1916) – серии завершаются одним климаксом • Теория поликлимакса • Теория мозаичного климакса или континуума климаксов Уиттекера (Wittaker, 1953) • Эдафический и климатический климаксы

 2. Тип почвы 3. Характер рельефа")
Основные факторы климакса 1. Климат (увлажнение и температура) 2. Тип почвы 3. Характер рельефа 4. Пожары 5. Животные-фитофаги

Пожары и гигантский лес из секвойядендронов Sequoiadendron giganteum

Жизненный цикл секвойядендронов = это цикл пожаров. Частота пожаров в лесу - от 3 до 9 лет.

Нет НИ ОДНОГО дерева в лесу не несущего следы пожаров

Кора взрослых деревьев достигает 1 м толщины

Без пожаров семена секвойядендронов не прорастают

WIN BIG! Какова вероятность для семечка стать деревом?




Ранне- и позднесукцессионные растения Характеристики Число семян Размер семян Дисперсия семян Ранние много мелкие большая Поздние мало крупные небольшая Механизмы дисп-ии Ветер, птицы, летучие мыши низкий низкая небольшой Гравитация, млекопитающие высокий высокая большой Индекс корни/побеги Выживаемость семян Размер взрослого растения Скорость роста высокая Устойчивость к стрессам высокая Продолжительность низкая жизни низкая высокая Huston 1994, Budowski 1965, 1970, Pianka 1970, Ricklefs 1973, Bazzaz 1979.

Смертность семян деревьев после 3 мес в условиях затенения 50% береза сосна 25 клен дуб гледичия каштан 0 1 10 1000 мг Вес семени


Сукцессии и метаболизм сообщества
 первичного органического вещества (за год)")
Климакс – равновесие скоростей потоков продукции и деструкции (разложения) первичного органического вещества (за год) P=R

1. связывается СО 2 только в процессе фотосинтеза растений, в листьях; 2. выделяется СО 2 в процессе дыхания растений (листьев, стеблей, корней), бактерий и грибов (как на поверхности земли, так и в почве)

Дыхание всей экосистемы Rtot складывается из R дыхания растений a (как надземных, так и подземных частей), Rh грибов, бактерий и животных

Соответственно возможны варианты: GPP = Rtot GPP > Rtot GPP< Rtot
 Net Ecosystem Production (NEP) NEP=GPP")
Чистая продукция экосистемы за год (г. С га-2 год-1) Net Ecosystem Production (NEP) NEP=GPP – ER = (GPP – Ra) – Rh = NPP - Rh

ER GPP – Ra – Rh = NEP = NEE NPP GPP – ER = NEE GPP > ER, +NEE – сток С-СО 2 из атмосферы GPP < ER, -NEE – источник С-CO 2 в атмосферу GPP – валовая первичная продукция Ra - дыхание автотрофов Rh – дыхание гетеротрофов NPP – чистая первичная продукция ER – валовое дыхание экосистемы NEP – чистая продукция экосистемы NEE – чистый поток (углерода)
,")
ВСЕ ЭТО ПОТОКИ (СКОРОСТЬ ПРОИЗВОДСТВА ИЛИ ПОТЕРИ МАССЫ УГЛЕРОДА на ПЛОЩАДИ или В ОБЪЕМЕ), которые измеряются одними единицами! Примеры: г С(СО 2)м-2 сут-1 Гт С(CO 2) км-2 год-1 г С(CO 2) м-3 сут-1

GPP ER = Ra +Rh Ra NPP Rh NEE Сток с водами Пожары Ветровая эрозия Депонирование в глубоких отложениях




 климакс в экосистемах цикличность – повторяемость состояний во времени, мозаичность – повторяемость")
Циклический (мозаичный) климакс в экосистемах цикличность – повторяемость состояний во времени, мозаичность – повторяемость состояний в пространстве



ER GPP – Ra – Rg = NEP = NEE NPP GPP – ER = NEE (NF) GPP > ER, +NEE – сток С(СО 2) из атмосферы; GPP < ER, -NEE – источник С (CO 2) в атмосферу; Единицы потоков: масса связанного (освобожденного) углерода в пространст за время (г на м 2 в сутки)




 развития растительности в тундрах")
Примеры цикличности (мозаичности) развития растительности в тундрах





«Ориентированные» озера в тундрах



/P = ± 3%")
Баланс 100%(P-R)/P = ± 3%
/P = -0. 5%")
Баланс 100%(P-R)/P = -0. 5%


Вывод: Климакс – состояние сообщества, жестко связанное с пространственным и временным масштабом
Лекция 13-14 биологи.pptx