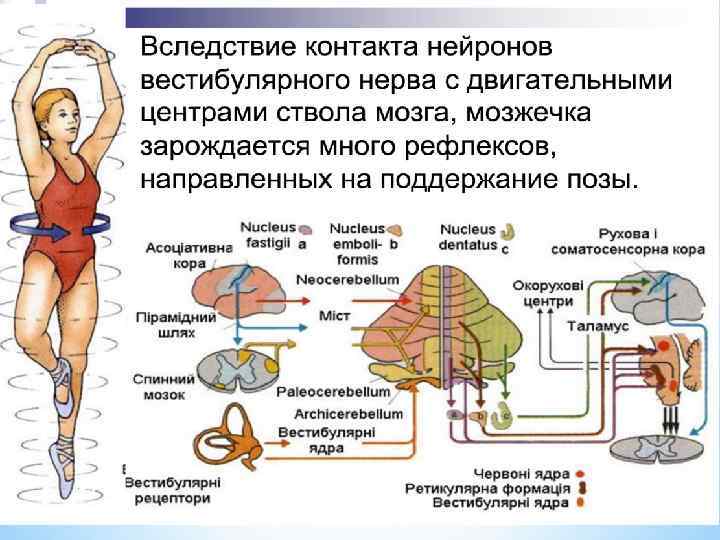
* Слух и равновесие
* Слух и равновесие
 * Регистрация двух сенсорных модальностей слуха и равновесия - происходит в ухе. * Оба органа (слуха и равновесия) формируют в толще височной кости преддверие и улитку преддверно-улитковый орган. * Рецепторные (волосковые) клетки органа слуха расположены в перепончатом канале улитки (кортиев орган), а органа равновесия (вестибулярный аппарат) в структурах преддверия - полукружных каналах, маточке и мешочке. *
* Регистрация двух сенсорных модальностей слуха и равновесия - происходит в ухе. * Оба органа (слуха и равновесия) формируют в толще височной кости преддверие и улитку преддверно-улитковый орган. * Рецепторные (волосковые) клетки органа слуха расположены в перепончатом канале улитки (кортиев орган), а органа равновесия (вестибулярный аппарат) в структурах преддверия - полукружных каналах, маточке и мешочке. *
 Преддверно-улитковый орган и рецепторные области органов слуха и равновесия. Движение перилимфы от овального к круглому окну обозначено стрелками.
Преддверно-улитковый орган и рецепторные области органов слуха и равновесия. Движение перилимфы от овального к круглому окну обозначено стрелками.
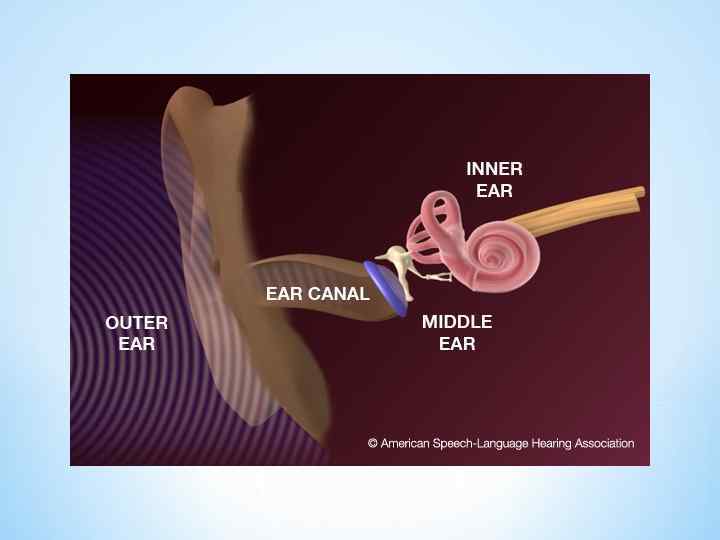
 А - общая схема наружного, среднего и внутреннего уха. Б - схема барабанной перепонки и цепочки слуховых косточек. В - схема поясняет, каким образом при смещении овальной пластинки стремени происходит движение жидкости в улитке и выгибается круглое окно
А - общая схема наружного, среднего и внутреннего уха. Б - схема барабанной перепонки и цепочки слуховых косточек. В - схема поясняет, каким образом при смещении овальной пластинки стремени происходит движение жидкости в улитке и выгибается круглое окно
 * Косточковая рычажная система уменьшает амплитуду, увеличивает силу движения примерно в 1, 3 раза. но * Площадь поверхности барабанной перепонки составляет 55 мм 2, площадь поверхности стремечко - 3, 2 мм 2. * Произведение этой 17 -кратной разницы и 1, 3 -кратного усиления рычажной системы дает примерно 22 -кратное увеличение общего давления на жидкость улитки по сравнению с силой давления звуковых волн на барабанную перепонку. * Поскольку инерция жидкости гораздо выше инерции воздуха, необходимо увеличить силу, чтобы вызвать вибрацию жидкости * Следовательно, барабанная перепонка и косточковая система обеспечивают согласование импедансов между звуковыми волнами в воздухе и звуковыми вибрациями жидкости в улитке. для звуковых частот между 300 и 3000 Гц согласование импедансов составляет 50 -75% (от идеального согласования), что обеспечивает использование большей части энергии входящих звуковых волн. *
* Косточковая рычажная система уменьшает амплитуду, увеличивает силу движения примерно в 1, 3 раза. но * Площадь поверхности барабанной перепонки составляет 55 мм 2, площадь поверхности стремечко - 3, 2 мм 2. * Произведение этой 17 -кратной разницы и 1, 3 -кратного усиления рычажной системы дает примерно 22 -кратное увеличение общего давления на жидкость улитки по сравнению с силой давления звуковых волн на барабанную перепонку. * Поскольку инерция жидкости гораздо выше инерции воздуха, необходимо увеличить силу, чтобы вызвать вибрацию жидкости * Следовательно, барабанная перепонка и косточковая система обеспечивают согласование импедансов между звуковыми волнами в воздухе и звуковыми вибрациями жидкости в улитке. для звуковых частот между 300 и 3000 Гц согласование импедансов составляет 50 -75% (от идеального согласования), что обеспечивает использование большей части энергии входящих звуковых волн. *
 *Ослабление звука путем сокращения мышцы, напрягающей барабанную перепонку, и стременной мышцы - увеличение ригидность системы, что снижает проведение низкочастотных волн на 30 -40 д. Б. * защищает улитку от повреждающих вибраций, связанных с чрезмерно громким звуком * маскирует низкочастотные звуки в шумной среде (звуками частотой выше 1000 Гц передается большинство значимой информации при речевом общении) *
*Ослабление звука путем сокращения мышцы, напрягающей барабанную перепонку, и стременной мышцы - увеличение ригидность системы, что снижает проведение низкочастотных волн на 30 -40 д. Б. * защищает улитку от повреждающих вибраций, связанных с чрезмерно громким звуком * маскирует низкочастотные звуки в шумной среде (звуками частотой выше 1000 Гц передается большинство значимой информации при речевом общении) *
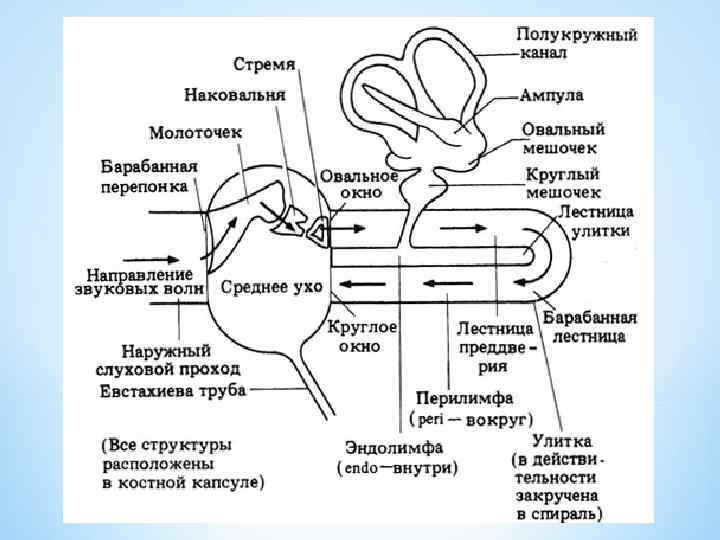
 * Наружное ухо * ушная раковина и наружный слуховой проход. * Среднее ухо * Полость сообщается с носоглоткой при помощи евстахиевой (слуховой) трубы и отделена от наружного слухового прохода барабанной перепонкой диаметром 9 мм, а от преддверия и барабанной лестницы улитки - овальным и круглым окнами соответственно. * Слуховая труба поддерживает давление воздуха в полости среднего уха на уровне атмосферного. * Барабанная * * перепонка передаёт звуковые колебания на три маленькие взаимосвязанные слуховые косточки: молоточек (прикреплён к барабанной перепонке), наковальню, стремечко (прикреплено к овальному окну). Слуховые косточки вибрируют в унисон и усиливают звук в двадцать раз. Мышцы среднего уха: стременная и напрягающая барабанную перепонку - в ответ на громкий звук рефлекторно сокращаются и уменьшают передачу звука к внутреннему уху. * Поскольку время акустического рефлекса составляет 150 мс, он не может предотвратить повреждающее действие резкого взрывного звука. * Внутреннее ухо * полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а полукружные каналы, маточка, мешочек и улитковый проток (перепончатый канал улитки) эндолимфой. * Между эндолимфой и перилимфой существует электрический потенциал - около 80 м. В (внутриулитковый, или эндокохлеарный потенциал) *
* Наружное ухо * ушная раковина и наружный слуховой проход. * Среднее ухо * Полость сообщается с носоглоткой при помощи евстахиевой (слуховой) трубы и отделена от наружного слухового прохода барабанной перепонкой диаметром 9 мм, а от преддверия и барабанной лестницы улитки - овальным и круглым окнами соответственно. * Слуховая труба поддерживает давление воздуха в полости среднего уха на уровне атмосферного. * Барабанная * * перепонка передаёт звуковые колебания на три маленькие взаимосвязанные слуховые косточки: молоточек (прикреплён к барабанной перепонке), наковальню, стремечко (прикреплено к овальному окну). Слуховые косточки вибрируют в унисон и усиливают звук в двадцать раз. Мышцы среднего уха: стременная и напрягающая барабанную перепонку - в ответ на громкий звук рефлекторно сокращаются и уменьшают передачу звука к внутреннему уху. * Поскольку время акустического рефлекса составляет 150 мс, он не может предотвратить повреждающее действие резкого взрывного звука. * Внутреннее ухо * полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а полукружные каналы, маточка, мешочек и улитковый проток (перепончатый канал улитки) эндолимфой. * Между эндолимфой и перилимфой существует электрический потенциал - около 80 м. В (внутриулитковый, или эндокохлеарный потенциал) *
 Строение улитки. А - относительное расположение улитки и вестибулярного аппарата среднего и наружного уха человека. Б - соотношение между пространствами улитки
Строение улитки. А - относительное расположение улитки и вестибулярного аппарата среднего и наружного уха человека. Б - соотношение между пространствами улитки
") *спирально закрученный костный канал, образующий 2, 5 завитка длиной около 35 мм. *Базилярная (основная) и вестибулярная мембраны, расположенные внутри канала улитки, делят полость канала на три части: на барабанную лестницу, вестибулярную лестницу и перепончатый канал улитки (средняя лестница, улитковый ход). *Эндолимфа заполняет перепончатый канал улитки, перилимфа - вестибулярную и барабанную лестницы. *В а перепончатом канале улитки на базилярной мембране расположен рецепторный аппарат улитки - кортиев (спиральный) орган. *
*спирально закрученный костный канал, образующий 2, 5 завитка длиной около 35 мм. *Базилярная (основная) и вестибулярная мембраны, расположенные внутри канала улитки, делят полость канала на три части: на барабанную лестницу, вестибулярную лестницу и перепончатый канал улитки (средняя лестница, улитковый ход). *Эндолимфа заполняет перепончатый канал улитки, перилимфа - вестибулярную и барабанную лестницы. *В а перепончатом канале улитки на базилярной мембране расположен рецепторный аппарат улитки - кортиев (спиральный) орган. *
 *Эндолимфа - вязкая жидкость, которая заполняет перепончатый канал улитки и соединяется через специальный канал с эндолимфой вестибулярного аппарата. * Концентрация K+ в эндолимфе в 100 раз больше, чем в спинномозговой жидкости (ликворе) и перилимфе; концентрация Na+ в эндолимфе в 10 раз меньше, чем в перилимфе. *Перилимфа по химическому составу близка к плазме крови и ликвору и занимает промежуточное положение между ними по содержанию белка. *
*Эндолимфа - вязкая жидкость, которая заполняет перепончатый канал улитки и соединяется через специальный канал с эндолимфой вестибулярного аппарата. * Концентрация K+ в эндолимфе в 100 раз больше, чем в спинномозговой жидкости (ликворе) и перилимфе; концентрация Na+ в эндолимфе в 10 раз меньше, чем в перилимфе. *Перилимфа по химическому составу близка к плазме крови и ликвору и занимает промежуточное положение между ними по содержанию белка. *
 относительно двух других лестниц. •") *Перепончатый канал улитки заряжен положительно (60 -80 м. В) относительно двух других лестниц. • Источник эндокохлеарного потенциала сосудистая полоска. • Волосковые клетки поляризованы эндокохлеарным потенциалом до критического уровня, что повышает их чувствительность к механическому воздействию *
*Перепончатый канал улитки заряжен положительно (60 -80 м. В) относительно двух других лестниц. • Источник эндокохлеарного потенциала сосудистая полоска. • Волосковые клетки поляризованы эндокохлеарным потенциалом до критического уровня, что повышает их чувствительность к механическому воздействию *
 и ее поперечный разрез ( Б ). Для") Схематическое изображение улитки ( А ) и ее поперечный разрез ( Б ). Для удобства восприятия улитка изображена развернутой. 1 – рейснерова мембрана, 2 - базилярная мембрана, 3 - вестибулярная лестница, 4 – средняя лестница, 5 - барабанная лестница, 6 – волосковые клетки, 7 – текториальная мембрана, 8 - чувствительный нейрон, 9 спиральный ганглий, 10 – овальное окно, 11 – круглое окно, 12 – геликотрема.
Схематическое изображение улитки ( А ) и ее поперечный разрез ( Б ). Для удобства восприятия улитка изображена развернутой. 1 – рейснерова мембрана, 2 - базилярная мембрана, 3 - вестибулярная лестница, 4 – средняя лестница, 5 - барабанная лестница, 6 – волосковые клетки, 7 – текториальная мембрана, 8 - чувствительный нейрон, 9 спиральный ганглий, 10 – овальное окно, 11 – круглое окно, 12 – геликотрема.
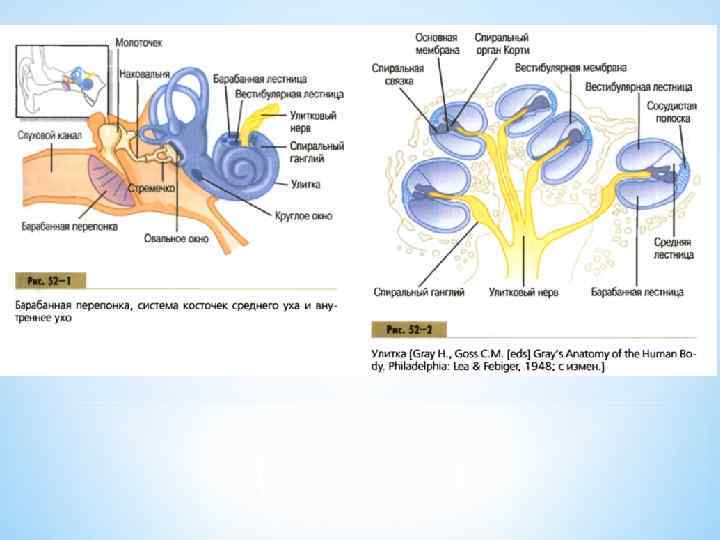
 А - схема поперечного разреза через улитку Б - строение кортиева органа
А - схема поперечного разреза через улитку Б - строение кортиева органа
 * содержит несколько рядов клеток, поддерживающих и волосковых * все клетки прикреплены к базилярной мембране, волосковые клетки своей свободной поверхностью связаны с покровной мембраной * волосковые клетки - рецепторные клетки органа Корти, образуют синаптические контакты с периферическими отростками чувствительных нейронов спирального ганглия * внутренние волосковые клетки образуют один ряд * на их свободной поверхности находится 30 -60 неподвижных микроотростков стереоцилий, проходящих через покровную мембрану * стереоцилии расположены полукругом (или в виде буквы V), открытым в сторону наружных структур кортиева органа. * общее количество клеток около 3500, они образуют примерно 95% синапсов с отростками чувствительных нейронов спирального ганглия. * наружные волосковые клетки расположены в 3 -5 рядов и также имеют стереоцилии * число клеток достигает 12 тыс. , но все вместе они образуют не более 5% синапсов с афферентными волокнами * возможно, наружные волосковые клетки как-то контролируют чувствительность внутренних волосковых клеток к различным звуковым уровням. *
* содержит несколько рядов клеток, поддерживающих и волосковых * все клетки прикреплены к базилярной мембране, волосковые клетки своей свободной поверхностью связаны с покровной мембраной * волосковые клетки - рецепторные клетки органа Корти, образуют синаптические контакты с периферическими отростками чувствительных нейронов спирального ганглия * внутренние волосковые клетки образуют один ряд * на их свободной поверхности находится 30 -60 неподвижных микроотростков стереоцилий, проходящих через покровную мембрану * стереоцилии расположены полукругом (или в виде буквы V), открытым в сторону наружных структур кортиева органа. * общее количество клеток около 3500, они образуют примерно 95% синапсов с отростками чувствительных нейронов спирального ганглия. * наружные волосковые клетки расположены в 3 -5 рядов и также имеют стереоцилии * число клеток достигает 12 тыс. , но все вместе они образуют не более 5% синапсов с афферентными волокнами * возможно, наружные волосковые клетки как-то контролируют чувствительность внутренних волосковых клеток к различным звуковым уровням. *
 Волосковые рецепторные клетки кортиева органа
Волосковые рецепторные клетки кортиева органа
 *
*
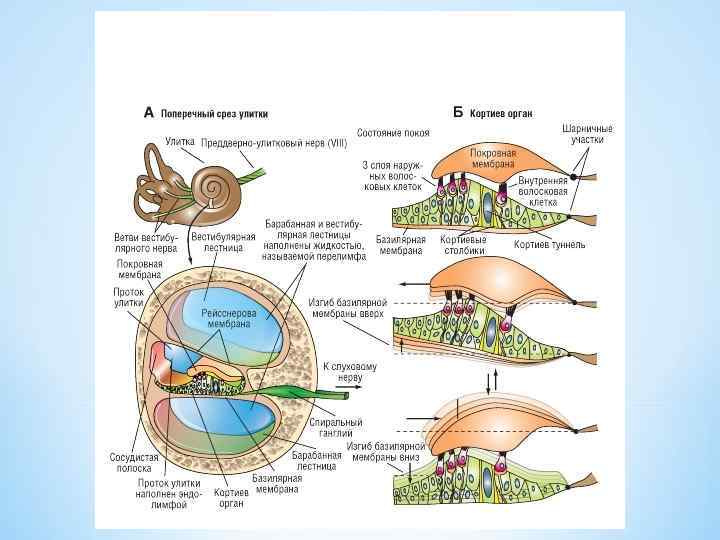
 *разделяет среднюю и барабанную лестницы, содержит до 30 тыс. базилярных волокон, идущих от костного стержня улитки по направлению к её наружной стенке *базилярные волокна тугие, эластичные, тростниковоподобные - прикреплены к стержню улитки только на одном конце, в результате могут гармонично вибрировать * длина увеличивается от основания к верхушке улитки – геликотреме * в области овального и круглого окна их длина составляет около 0, 04 мм, в области геликотремы они длиннее в 12 раз * диаметр базилярных волокон уменьшается от основания к верхушке улитки примерно в 100 раз Короткие базилярные волокна возле овального окна вибрируют лучше на высоких частотах, тогда как длинные волокна вблизи геликотремы лучше вибрируют на низких частотах. *
*разделяет среднюю и барабанную лестницы, содержит до 30 тыс. базилярных волокон, идущих от костного стержня улитки по направлению к её наружной стенке *базилярные волокна тугие, эластичные, тростниковоподобные - прикреплены к стержню улитки только на одном конце, в результате могут гармонично вибрировать * длина увеличивается от основания к верхушке улитки – геликотреме * в области овального и круглого окна их длина составляет около 0, 04 мм, в области геликотремы они длиннее в 12 раз * диаметр базилярных волокон уменьшается от основания к верхушке улитки примерно в 100 раз Короткие базилярные волокна возле овального окна вибрируют лучше на высоких частотах, тогда как длинные волокна вблизи геликотремы лучше вибрируют на низких частотах. *
 *первичным действием звуковой волны, входящей в овальное окно, должно быть выгибание базальной мембраны у основания улитки в направлении круглого окна *однако эластическое напряжение, возникающее в банальных волокнах при их выгибании в сторону круглого окна, инициирует волну жидкости, которая «бежит» вдоль базальной мембраны в направлении геликотремы *каждая волна, относительно слабая на входе, становится сильной, когда достигает части базальной мембраны, имеющей естественную резонансную частоту, равную частоте соответствующего звука *в этой точке базальная мембрана может вибрировать с такой легкостью, что энергия волны рассеивается *
*первичным действием звуковой волны, входящей в овальное окно, должно быть выгибание базальной мембраны у основания улитки в направлении круглого окна *однако эластическое напряжение, возникающее в банальных волокнах при их выгибании в сторону круглого окна, инициирует волну жидкости, которая «бежит» вдоль базальной мембраны в направлении геликотремы *каждая волна, относительно слабая на входе, становится сильной, когда достигает части базальной мембраны, имеющей естественную резонансную частоту, равную частоте соответствующего звука *в этой точке базальная мембрана может вибрировать с такой легкостью, что энергия волны рассеивается *
 и барабанная лестницы (снизу) в направлении от овального (слева вверху) через") * Вестибулярная (сверху) и барабанная лестницы (снизу) в направлении от овального (слева вверху) через геликотрему (справа) к круглому (слева внизу) окну; базилярная мембрана, разделяющая названные лестницы - горизонтальная линия. Средняя лестница в модели не учтена. * Слева: движение волн высоко- (А), средне- (Б) и низкочастотных (В) звуков вдоль базилярной мембраны. * Справа: корреляция между частотой звука и амплитудой колебаний базилярной мембраны в зависимости от расстояния от основания улитки. *
* Вестибулярная (сверху) и барабанная лестницы (снизу) в направлении от овального (слева вверху) через геликотрему (справа) к круглому (слева внизу) окну; базилярная мембрана, разделяющая названные лестницы - горизонтальная линия. Средняя лестница в модели не учтена. * Слева: движение волн высоко- (А), средне- (Б) и низкочастотных (В) звуков вдоль базилярной мембраны. * Справа: корреляция между частотой звука и амплитудой колебаний базилярной мембраны в зависимости от расстояния от основания улитки. *
 Схема, иллюстрирующая колебания основной мембраны при действии звуков разной частоты: А – бегущая волна в базилярной мембране, Б – амплитуда колебаний базилярной мембраны (бегущей волны) при действии звуков разных частот. 1 - проксимальный конец базилярной мембраны (обращен к овальному окну) чувствителен к колебаниям высоких частот, 2 - дистальный конец базилярной мембраны (обращен к геликотреме) чувствителен к колебаниям низких частот базальные волна, столбы Корти и ретикулярная мембрана двигаются, как единая ригидная структура при движении основной мембраны вверх ретикулярная мембрана сдвигается вверх и внутрь по направлению к центральному костному стержню улитки
Схема, иллюстрирующая колебания основной мембраны при действии звуков разной частоты: А – бегущая волна в базилярной мембране, Б – амплитуда колебаний базилярной мембраны (бегущей волны) при действии звуков разных частот. 1 - проксимальный конец базилярной мембраны (обращен к овальному окну) чувствителен к колебаниям высоких частот, 2 - дистальный конец базилярной мембраны (обращен к геликотреме) чувствителен к колебаниям низких частот базальные волна, столбы Корти и ретикулярная мембрана двигаются, как единая ригидная структура при движении основной мембраны вверх ретикулярная мембрана сдвигается вверх и внутрь по направлению к центральному костному стержню улитки
 * Цепочка передачи звукового давления: * барабанная перепонка - молоточек - наковальня - стремя - мембрана овального окна - перилимфа - базилярная и текториальная мембраны - мембрана круглого окна. * При смещении стремени перилимфа перемещается по вестибулярной лестнице и затем через геликотрему по барабанной лестнице к круглому окну. * Жидкость, сдвинутая смещением мембраны овального окна, создаёт избыточное давление в вестибулярном канале. Под действием этого давления базилярная мембрана смещается в сторону барабанной лестницы. * Колебательная реакция в виде волны распространяется от базилярной мембраны к геликотреме. * Смещение текториальной мембраны относительно волосковых клеток при действии звука вызывает их возбуждение. * Возникающая электрическая реакция (микрофонный эффект) повторяет форму звукового сигнала. *
* Цепочка передачи звукового давления: * барабанная перепонка - молоточек - наковальня - стремя - мембрана овального окна - перилимфа - базилярная и текториальная мембраны - мембрана круглого окна. * При смещении стремени перилимфа перемещается по вестибулярной лестнице и затем через геликотрему по барабанной лестнице к круглому окну. * Жидкость, сдвинутая смещением мембраны овального окна, создаёт избыточное давление в вестибулярном канале. Под действием этого давления базилярная мембрана смещается в сторону барабанной лестницы. * Колебательная реакция в виде волны распространяется от базилярной мембраны к геликотреме. * Смещение текториальной мембраны относительно волосковых клеток при действии звука вызывает их возбуждение. * Возникающая электрическая реакция (микрофонный эффект) повторяет форму звукового сигнала. *
. а - распространение звуковой волны по наружному и среднему уху;") Воздушное проведение звука (схема). а - распространение звуковой волны по наружному и среднему уху; б - распространение звуковой волны по жидкостям улитки (показано стрелками). Благодаря смещаемости мембраны круглого окна получается выгибание основной перепонки в сторону барабанной лестницы. вибрация черепа может вызвать вибрацию жидкости в самой улитке Костное проведение звука. 1 - движение головы, 2 - движение перилимфы; а - инерционный механизм костной проводимости (стрелкой показано движение головы); б - компрессионный механизм костной проводимости.
Воздушное проведение звука (схема). а - распространение звуковой волны по наружному и среднему уху; б - распространение звуковой волны по жидкостям улитки (показано стрелками). Благодаря смещаемости мембраны круглого окна получается выгибание основной перепонки в сторону барабанной лестницы. вибрация черепа может вызвать вибрацию жидкости в самой улитке Костное проведение звука. 1 - движение головы, 2 - движение перилимфы; а - инерционный механизм костной проводимости (стрелкой показано движение головы); б - компрессионный механизм костной проводимости.
 * Когда подошва стремени движется внутрь против овального окна, круглое окно выпячивается наружу, потому что улитка со всех сторон окружена костной тканью. * Начальный эффект звуковой волны, входящей в овальное окно, проявляется в прогибании базилярной мембраны в области основания улитки в направлении круглого окна. Однако эластическое напряжение базилярных волокон создаёт волну жидкости, которая пробегает вдоль базилярной мембраны в направлении геликотремы. * Каждая волна сначала относительно слаба, но она становится более сильной, когда достигает той части базилярной мембраны, где собственный резонанс мембраны становится равным частоте звуковой волны. * В этой точке базилярная мембрана может свободно вибрировать вперёд и назад, т. е. энергия звуковой волны рассеивается, волна прерывается в этой точке и теряет способность продвигаться вдоль базилярной мембраны. * Таким образом, * звуковая волна высокой частоты проходит короткое расстояние вдоль базилярной мембраны, прежде чем достичь своей резонансной точки и исчезнуть; * звуковые волны средней частоты проходят примерно половину пути и затем прекращаются; * наконец, звуковые волны очень низкой частоты проходят вдоль мембраны почти до геликотремы. *
* Когда подошва стремени движется внутрь против овального окна, круглое окно выпячивается наружу, потому что улитка со всех сторон окружена костной тканью. * Начальный эффект звуковой волны, входящей в овальное окно, проявляется в прогибании базилярной мембраны в области основания улитки в направлении круглого окна. Однако эластическое напряжение базилярных волокон создаёт волну жидкости, которая пробегает вдоль базилярной мембраны в направлении геликотремы. * Каждая волна сначала относительно слаба, но она становится более сильной, когда достигает той части базилярной мембраны, где собственный резонанс мембраны становится равным частоте звуковой волны. * В этой точке базилярная мембрана может свободно вибрировать вперёд и назад, т. е. энергия звуковой волны рассеивается, волна прерывается в этой точке и теряет способность продвигаться вдоль базилярной мембраны. * Таким образом, * звуковая волна высокой частоты проходит короткое расстояние вдоль базилярной мембраны, прежде чем достичь своей резонансной точки и исчезнуть; * звуковые волны средней частоты проходят примерно половину пути и затем прекращаются; * наконец, звуковые волны очень низкой частоты проходят вдоль мембраны почти до геликотремы. *
 А - общая схема наружного, среднего и внутреннего уха. Б - схема барабанной перепонки и цепочки слуховых косточек. В - схема поясняет, каким образом при смещении овальной пластинки стремени происходит движение жидкости в улитке и выгибается круглое окно
А - общая схема наружного, среднего и внутреннего уха. Б - схема барабанной перепонки и цепочки слуховых косточек. В - схема поясняет, каким образом при смещении овальной пластинки стремени происходит движение жидкости в улитке и выгибается круглое окно
 * Неподвижные и упругие стереоцилии направлены кверху от апикальной поверхности волосковых клеток и проникают в покровную мембрану. * Базальная часть волосковых рецепторных клеток прикреплена к содержащей базилярные волокна мембране. * Волосковые клетки возбуждаются, как только базилярная мембрана начинает вибрировать вместе с прикреплёнными к ней клетками и покровной мембраной. * Возбуждение волосковых клеток (генерация рецепторного потенциала) начинается в стереоцилиях. * Рецепторный потенциал * Возникшее натяжение стереоцилий вызывает механические преобразования, открывающие 200 -300 катионных каналов. * Ионы *В K+ из эндолимфы поступают внутрь стереоцилии, вызывая деполяризацию мембраны волосковой клетки. синапсах между рецепторной клеткой и афферентным нервным окончанием выделяется быстродействующий нейромедиатор – глутамат. * Происходит его взаимодействие с глутаматными рецепторами, деполяризация постсинаптической мембраны и генерация ПД. *
* Неподвижные и упругие стереоцилии направлены кверху от апикальной поверхности волосковых клеток и проникают в покровную мембрану. * Базальная часть волосковых рецепторных клеток прикреплена к содержащей базилярные волокна мембране. * Волосковые клетки возбуждаются, как только базилярная мембрана начинает вибрировать вместе с прикреплёнными к ней клетками и покровной мембраной. * Возбуждение волосковых клеток (генерация рецепторного потенциала) начинается в стереоцилиях. * Рецепторный потенциал * Возникшее натяжение стереоцилий вызывает механические преобразования, открывающие 200 -300 катионных каналов. * Ионы *В K+ из эндолимфы поступают внутрь стереоцилии, вызывая деполяризацию мембраны волосковой клетки. синапсах между рецепторной клеткой и афферентным нервным окончанием выделяется быстродействующий нейромедиатор – глутамат. * Происходит его взаимодействие с глутаматными рецепторами, деполяризация постсинаптической мембраны и генерация ПД. *
 *Когда базилярные волокна изгибаются в направлении вестибулярной лестницы, волосковые клетки деполяризуются; но при движении базилярной мембраны в противоположном направлении они гиперполяризуются (такая же дирекциональная чувствительность, определяющая электрический ответ рецепторной клетки, характерна для волосковых клеток органа равновесия). 1 – волосковая клетка, 2 – киноцилии, 3 - стереоцилия, 4 - желатинообразная купула, 5 – афферентное нервное волокно, 6 – эфферентное нервное волокно, 7 – чувствительный нейрон, 8 – изменение мембранного потенциала волосковой клетки, 9 – изменение частоты нервных импульсов в чувствительном нейроне. *
*Когда базилярные волокна изгибаются в направлении вестибулярной лестницы, волосковые клетки деполяризуются; но при движении базилярной мембраны в противоположном направлении они гиперполяризуются (такая же дирекциональная чувствительность, определяющая электрический ответ рецепторной клетки, характерна для волосковых клеток органа равновесия). 1 – волосковая клетка, 2 – киноцилии, 3 - стереоцилия, 4 - желатинообразная купула, 5 – афферентное нервное волокно, 6 – эфферентное нервное волокно, 7 – чувствительный нейрон, 8 – изменение мембранного потенциала волосковой клетки, 9 – изменение частоты нервных импульсов в чувствительном нейроне. *
 * Частота звуковой волны жёстко «привязана» к конкретному участку базилярной мембраны. * Существует пространственная организация нервных волокон на протяжении всего слухового пути - от улитки до коры больших полушарий. * Регистрация сигналов в слуховом тракте мозгового ствола и в слуховом поле коры больших полушарий показывает, что имеются специальные нейроны мозга, возбуждаемые конкретными звуковыми частотами. * Главным методом, используемым нервной системой для определения звуковых частот, является установление того участка базилярной мембраны, который наиболее стимулирован, - так называемый «принцип места» . *
* Частота звуковой волны жёстко «привязана» к конкретному участку базилярной мембраны. * Существует пространственная организация нервных волокон на протяжении всего слухового пути - от улитки до коры больших полушарий. * Регистрация сигналов в слуховом тракте мозгового ствола и в слуховом поле коры больших полушарий показывает, что имеются специальные нейроны мозга, возбуждаемые конкретными звуковыми частотами. * Главным методом, используемым нервной системой для определения звуковых частот, является установление того участка базилярной мембраны, который наиболее стимулирован, - так называемый «принцип места» . *
 * Слуховая система использует для определения громкости несколько механизмов. * Громкий звук повышает амплитуду колебаний базилярной мембраны, что увеличивает количество возбуждённых волосковых клеток, а это приводит к пространственной суммации импульсов и передаче возбуждения по многим нервным волокнам. * Наружные волосковые клетки не возбуждаются до тех пор, пока вибрация базилярной мембраны не достигнет высокой интенсивности. Стимуляция этих клеток может оцениваться нервной системой как показатель действительно громкого звука. ØОщущение возрастающей громкости звука не следует строго параллельно возрастанию силы звука (уровню звуковой мощности). ØЧтобы оценить уровень звуковой мощности, используют логарифмический показатель реальной силы звука: 10 -кратное увеличение энергии звука - 1 бел (Б). 0, 1 Б называется децибел (д. Б). 1 д. Б - увеличение звуковой энергии в 1, 26 раза - интенсивность звука по отношению к пороговой величине (2 х10 -5 дин/см 2) (1 дин = 10 -5 Н). При обычном восприятии звука во время общения человек может различать изменения интенсивности звука в 1 д. Б. *
* Слуховая система использует для определения громкости несколько механизмов. * Громкий звук повышает амплитуду колебаний базилярной мембраны, что увеличивает количество возбуждённых волосковых клеток, а это приводит к пространственной суммации импульсов и передаче возбуждения по многим нервным волокнам. * Наружные волосковые клетки не возбуждаются до тех пор, пока вибрация базилярной мембраны не достигнет высокой интенсивности. Стимуляция этих клеток может оцениваться нервной системой как показатель действительно громкого звука. ØОщущение возрастающей громкости звука не следует строго параллельно возрастанию силы звука (уровню звуковой мощности). ØЧтобы оценить уровень звуковой мощности, используют логарифмический показатель реальной силы звука: 10 -кратное увеличение энергии звука - 1 бел (Б). 0, 1 Б называется децибел (д. Б). 1 д. Б - увеличение звуковой энергии в 1, 26 раза - интенсивность звука по отношению к пороговой величине (2 х10 -5 дин/см 2) (1 дин = 10 -5 Н). При обычном восприятии звука во время общения человек может различать изменения интенсивности звука в 1 д. Б. *
 Схема, иллюстрирующая принцип распознавания частоты и громкости звуков в улитке. Цифрами на схеме обозначены частоты нервных импульсов (имп/с) пяти рецепторов, расположенных вдоль основной мембраны. Звуки А и Б будут восприниматься, как одинаковые по частоте, но разные по громкости, звуки Б и В – как одинаково громкие, но разные по частоте, а звуки А и В – как разные и по частоте и по громкости.
Схема, иллюстрирующая принцип распознавания частоты и громкости звуков в улитке. Цифрами на схеме обозначены частоты нервных импульсов (имп/с) пяти рецепторов, расположенных вдоль основной мембраны. Звуки А и Б будут восприниматься, как одинаковые по частоте, но разные по громкости, звуки Б и В – как одинаково громкие, но разные по частоте, а звуки А и В – как разные и по частоте и по громкости.
 * Афферентные нервные волокна от улитки входят * в спиральный ганглий и от него поступают *в дорсальные и вентральные улитковые ядра, расположенные в верхней части продолговатого мозга, восходящие нервные волокна образуют синапсы с нейронами второго порядка, аксоны которых частично переходят на противоположную сторону к ядрам верхней оливы, а частично оканчиваются на ядрах верхней оливы этой же стороны * далее слуховые пути поднимаются вверх через латеральный лемнисковой путь; часть волокон оканчивается в латеральных лемнисковых ядрах, а большинство аксонов минует эти ядра и * следует до нижнего двухолмия, где все или почти все слуховые волокна образуют синапсы * далее слуховой путь проходит к медиальным коленчатым телам, где все волокна заканчиваются синапсами * окончательно слуховой путь завершается в слуховой коре, располагающейся главным образом в верхней извилине височной доли Уже на уровне среднего мозга появляются нейроны, детектирующие на принципах латерального и возвратного торможения несколько признаков звука. *
* Афферентные нервные волокна от улитки входят * в спиральный ганглий и от него поступают *в дорсальные и вентральные улитковые ядра, расположенные в верхней части продолговатого мозга, восходящие нервные волокна образуют синапсы с нейронами второго порядка, аксоны которых частично переходят на противоположную сторону к ядрам верхней оливы, а частично оканчиваются на ядрах верхней оливы этой же стороны * далее слуховые пути поднимаются вверх через латеральный лемнисковой путь; часть волокон оканчивается в латеральных лемнисковых ядрах, а большинство аксонов минует эти ядра и * следует до нижнего двухолмия, где все или почти все слуховые волокна образуют синапсы * далее слуховой путь проходит к медиальным коленчатым телам, где все волокна заканчиваются синапсами * окончательно слуховой путь завершается в слуховой коре, располагающейся главным образом в верхней извилине височной доли Уже на уровне среднего мозга появляются нейроны, детектирующие на принципах латерального и возвратного торможения несколько признаков звука. *
; вид на ствол мозга сзади. Мозжечок и кора больших полушарий") Основные слуховые пути (А); вид на ствол мозга сзади. Мозжечок и кора больших полушарий удалены. Слуховая кора (Б).
Основные слуховые пути (А); вид на ствол мозга сзади. Мозжечок и кора больших полушарий удалены. Слуховая кора (Б).
 Кохлеарный нерв 1, вентральное кохлеарное ядро 2, дорсальное кохлеарное ядро, оливарный комплекс 4, ядро латеральной петли 6, добавочное ядро 5, нижние холмики 7, медиальное коленчатое тело 8, первичная звуковая кора 9
Кохлеарный нерв 1, вентральное кохлеарное ядро 2, дорсальное кохлеарное ядро, оливарный комплекс 4, ядро латеральной петли 6, добавочное ядро 5, нижние холмики 7, медиальное коленчатое тело 8, первичная звуковая кора 9
 *Проекционные области слуховой коры располагаются в верхней части верхней височной извилины, простираются на наружную сторону височной доли, захватывая часть островковой коры и теменной покрышки. *Первичная слуховая кора непосредственно получает сигналы: от внутреннего (медиального) коленчатого тела. *Слуховая ассоциативная область вторично возбуждается импульсами из первичной слуховой коры и таламических областей, граничащих с медиальным коленчатым телом. *
*Проекционные области слуховой коры располагаются в верхней части верхней височной извилины, простираются на наружную сторону височной доли, захватывая часть островковой коры и теменной покрышки. *Первичная слуховая кора непосредственно получает сигналы: от внутреннего (медиального) коленчатого тела. *Слуховая ассоциативная область вторично возбуждается импульсами из первичной слуховой коры и таламических областей, граничащих с медиальным коленчатым телом. *
 *В каждой из шести тонотопических карт звуки высокой частоты возбуждают нейроны в задней части карты, в то время как звуки низкой частоты возбуждают нейроны в передней её части. *Предполагают, что каждая отдельная область воспринимает свои специфические особенности звука. * Например, одна большая карта в первичной слуховой коре почти целиком дискриминирует звуки, которые субъекту кажутся высокими. * Другая карта используется, чтобы определить направление поступающего звука. * Некоторые области слуховой коры выявляют специальные качества звуковых сигналов (например, неожиданное начало звуков или их модуляции). *
*В каждой из шести тонотопических карт звуки высокой частоты возбуждают нейроны в задней части карты, в то время как звуки низкой частоты возбуждают нейроны в передней её части. *Предполагают, что каждая отдельная область воспринимает свои специфические особенности звука. * Например, одна большая карта в первичной слуховой коре почти целиком дискриминирует звуки, которые субъекту кажутся высокими. * Другая карта используется, чтобы определить направление поступающего звука. * Некоторые области слуховой коры выявляют специальные качества звуковых сигналов (например, неожиданное начало звуков или их модуляции). *
 *Диапазон звуковой частоты, на которую отвечают нейроны слуховой коры уже, чем для нейронов спирального ганглия и мозгового ствола. *Это объясняется, * высокой степенью специализации нейронов коры, * феном латерального и возвратного торможения, усиливающего разрешающую способность нейронов воспринимать необходимую частоту звука. *
*Диапазон звуковой частоты, на которую отвечают нейроны слуховой коры уже, чем для нейронов спирального ганглия и мозгового ствола. *Это объясняется, * высокой степенью специализации нейронов коры, * феном латерального и возвратного торможения, усиливающего разрешающую способность нейронов воспринимать необходимую частоту звука. *
 * Направление источника звука * два уха, работающих в унисон, могут обнаруживать источник звука по разнице в громкости и времени, которое ему требуется, чтобы достичь обеих сторон головы. * Человек определяет звук, идущий к нему, двумя путями: * Временем задержки между поступлением звука в одно противоположное ухо и в * Если источник звука находится по средней линии спереди или сзади, то даже минимальный сдвиг от средней линии воспринимается человеком. * Сравнение минимальной разницы во времени поступления звука ЦНС осуществляет в точках, где происходит конвергенция слуховых сигналов. * Этими точками конвергенции являются верхние оливы, нижнее двухолмие, первичная слуховая кора. * Различием между интенсивностью звуков в двух ушах. * При высоких частотах звука размер головы заметно превышает длину звуковой волны и волна отражается головой. Вследствие этого возникает разница в интенсивности звуков, поступающих к правому и левому уху. *
* Направление источника звука * два уха, работающих в унисон, могут обнаруживать источник звука по разнице в громкости и времени, которое ему требуется, чтобы достичь обеих сторон головы. * Человек определяет звук, идущий к нему, двумя путями: * Временем задержки между поступлением звука в одно противоположное ухо и в * Если источник звука находится по средней линии спереди или сзади, то даже минимальный сдвиг от средней линии воспринимается человеком. * Сравнение минимальной разницы во времени поступления звука ЦНС осуществляет в точках, где происходит конвергенция слуховых сигналов. * Этими точками конвергенции являются верхние оливы, нижнее двухолмие, первичная слуховая кора. * Различием между интенсивностью звуков в двух ушах. * При высоких частотах звука размер головы заметно превышает длину звуковой волны и волна отражается головой. Вследствие этого возникает разница в интенсивности звуков, поступающих к правому и левому уху. *
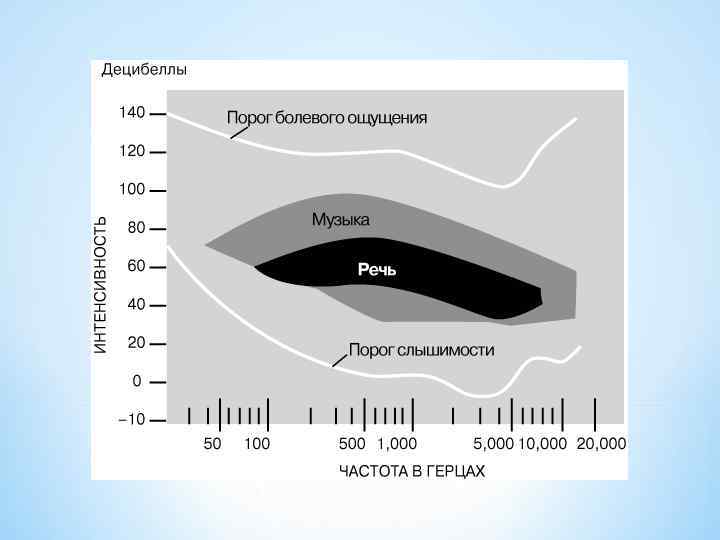
 * Диапазон частот, который воспринимает человек, включает около 10 октав музыкальной шкалы (от 16 Гц до 20 к. Гц). * Этот диапазон постепенно сужается с возрастом, так как снижается восприятие высоких частот. * Различение частоты звука характеризуется минимальным различием по частоте двух близких звуков, которое ещё улавливается человеком. * Абсолютный порог слуховой чувствительности - минимальная сила звука, которую слышит человек в 50% случаев его предъявления. * Порог слышимости зависит от частоты звуковых волн. * Максимальная чувствительность слуха у людей располагается в области от 500 до 4000 Гц. * В диапазоне этих частот располагается область звукового восприятия человеческой речи. * Чувствительность снижается. к звуковым частотам ниже 500 Гц прогрессивно * Это предохраняет человека от возможного постоянного ощущения низкочастотных колебаний и шумов, производимых собственным телом. *
* Диапазон частот, который воспринимает человек, включает около 10 октав музыкальной шкалы (от 16 Гц до 20 к. Гц). * Этот диапазон постепенно сужается с возрастом, так как снижается восприятие высоких частот. * Различение частоты звука характеризуется минимальным различием по частоте двух близких звуков, которое ещё улавливается человеком. * Абсолютный порог слуховой чувствительности - минимальная сила звука, которую слышит человек в 50% случаев его предъявления. * Порог слышимости зависит от частоты звуковых волн. * Максимальная чувствительность слуха у людей располагается в области от 500 до 4000 Гц. * В диапазоне этих частот располагается область звукового восприятия человеческой речи. * Чувствительность снижается. к звуковым частотам ниже 500 Гц прогрессивно * Это предохраняет человека от возможного постоянного ощущения низкочастотных колебаний и шумов, производимых собственным телом. *
 *
*
 и сферического (саккулус") *вестибулярный аппарат: улитковый проток, полукружные каналы, камеры - эллиптического (утрикулюс _) и сферического (саккулус I) мешочков *макулы (пятна) - сенсорные органы эллиптического и сферического мешочков для определения ориентации головы по отношению к силе тяжести *кристаллы карбоната кальция - статоконии (отолиты) * вес статоконий сгибает реснички в направлении действия силы тяжести *стереоцилии и киноцилия * мельчайшие нитевидные сцепки связывают верхушку каждой стереоцилии с соседней *
*вестибулярный аппарат: улитковый проток, полукружные каналы, камеры - эллиптического (утрикулюс _) и сферического (саккулус I) мешочков *макулы (пятна) - сенсорные органы эллиптического и сферического мешочков для определения ориентации головы по отношению к силе тяжести *кристаллы карбоната кальция - статоконии (отолиты) * вес статоконий сгибает реснички в направлении действия силы тяжести *стереоцилии и киноцилия * мельчайшие нитевидные сцепки связывают верхушку каждой стереоцилии с соседней *
 *в условиях покоя по нервным волокнам, идущим от волосковых клеток, постоянно проводятся импульсы с частотой 100 имп/с *в каждой макуле каждая из волосковых ориентирована в определенном направлении клеток * следовательно, для каждой ориентации головы в гравитационном поле в нервных волокнах, идущих от макулы, возникает разный «рисунок» возбуждения *каналы - передний, задний и латеральный (горизонтальный) *расширение – ампула * ампулярный гребешок * рыхлая студенистая тканевая масса купол (купула) *
*в условиях покоя по нервным волокнам, идущим от волосковых клеток, постоянно проводятся импульсы с частотой 100 имп/с *в каждой макуле каждая из волосковых ориентирована в определенном направлении клеток * следовательно, для каждой ориентации головы в гравитационном поле в нервных волокнах, идущих от макулы, возникает разный «рисунок» возбуждения *каналы - передний, задний и латеральный (горизонтальный) *расширение – ампула * ампулярный гребешок * рыхлая студенистая тканевая масса купол (купула) *
 *в ответ на «рисунок» вестибулярная, мозжечковая, ретикулярная двигательные системы мозга возбуждают соответствующие позные мышцы поддержания надлежащего равновесия *полукружные каналы сообщают о том, что голова начинает или прекращает вращаться в одном или в другом направлении *информация от них оповещает ЦНС о том, что примерно через долю секунды человек потеряет равновесие, если не провести некоторую упреждающую коррекцию *
*в ответ на «рисунок» вестибулярная, мозжечковая, ретикулярная двигательные системы мозга возбуждают соответствующие позные мышцы поддержания надлежащего равновесия *полукружные каналы сообщают о том, что голова начинает или прекращает вращаться в одном или в другом направлении *информация от них оповещает ЦНС о том, что примерно через долю секунды человек потеряет равновесие, если не провести некоторую упреждающую коррекцию *
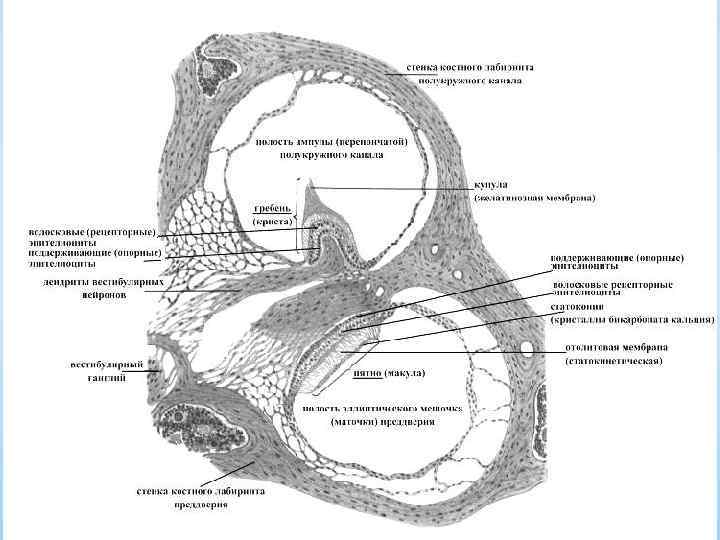
 Перепончатый лабиринт и организация ампулярного гребешка и макулы
Перепончатый лабиринт и организация ампулярного гребешка и макулы
 * = преддверный аппарат = орган равновесия * расположен в каменистой части височной кости * состоит из костного и перепончатого лабиринтов * костный лабиринт - система полукружных протоков; сообщающаяся с ними полость – преддверие * перепончатый лабиринт - система тонкостенных трубок и мешочков, расположенная внутри костного лабиринта * в костных ампулах перепончатые каналы расширяются *в * в каждом ампулярном расширении полукружного канала находятся гребешки преддверии перепончатого лабиринта образуются две сообщающиеся между собой полости: маточка, в которую открываются перепончатые полукружные каналы, и мешочек * чувствительные области полостей - пятна * Перепончатые полукружные каналы, маточка и мешочек заполнены эндолимфой и сообщаются с улиткой, а также с расположенным в полости черепа эндолимфатическим мешком. * Гребешки и пятна - воспринимающие области вестибулярного органа - содержат рецепторные волосковые клетки. * В полукружных каналах происходит регистрация вращательных движений (угловое ускорение), в маточке и мешочке - линейное ускорение. *
* = преддверный аппарат = орган равновесия * расположен в каменистой части височной кости * состоит из костного и перепончатого лабиринтов * костный лабиринт - система полукружных протоков; сообщающаяся с ними полость – преддверие * перепончатый лабиринт - система тонкостенных трубок и мешочков, расположенная внутри костного лабиринта * в костных ампулах перепончатые каналы расширяются *в * в каждом ампулярном расширении полукружного канала находятся гребешки преддверии перепончатого лабиринта образуются две сообщающиеся между собой полости: маточка, в которую открываются перепончатые полукружные каналы, и мешочек * чувствительные области полостей - пятна * Перепончатые полукружные каналы, маточка и мешочек заполнены эндолимфой и сообщаются с улиткой, а также с расположенным в полости черепа эндолимфатическим мешком. * Гребешки и пятна - воспринимающие области вестибулярного органа - содержат рецепторные волосковые клетки. * В полукружных каналах происходит регистрация вращательных движений (угловое ускорение), в маточке и мешочке - линейное ускорение. *
, линейные (Б) и угловые (В)") Схема, иллюстрирующая механизм восприятия положения головы в пространстве (А), линейные (Б) и угловые (В) ускорения.
Схема, иллюстрирующая механизм восприятия положения головы в пространстве (А), линейные (Б) и угловые (В) ускорения.
 и пятна (Б, В). ОМ") Рецепторная область органа равновесия. Вертикальные срезы через гребешок (А) и пятна (Б, В). ОМ - отолитовая мембрана, О - отолиты, ПК - поддерживающая клетка, РК - рецепторная клетка.
Рецепторная область органа равновесия. Вертикальные срезы через гребешок (А) и пятна (Б, В). ОМ - отолитовая мембрана, О - отолиты, ПК - поддерживающая клетка, РК - рецепторная клетка.
 А - строение вестибулярного аппарата. Б - вид сверху на основание черепа. Заметна ориентация структур внутреннего уха. В - схема разреза через ампулярный гребешок. Стереоцилии и киноцилия каждой волосковой клетки погружены в купулу. Положение купулы до поворота головы (Г) и во время поворота (Д). Е - строение отолитовых органов
А - строение вестибулярного аппарата. Б - вид сверху на основание черепа. Заметна ориентация структур внутреннего уха. В - схема разреза через ампулярный гребешок. Стереоцилии и киноцилия каждой волосковой клетки погружены в купулу. Положение купулы до поворота головы (Г) и во время поворота (Д). Е - строение отолитовых органов
 * В эпителии пятен и гребешков находятся чувствительные волосковые и поддерживающие клетки. * Эпителий пятен покрыт студенистой отолитовой мембраной, содержащей отолиты - кристаллы карбоната кальция. * Эпителий гребешков окружён желеобразным прозрачным куполом, легко смещающимся при движениях эндолимфы. * Волосковые клетки находятся в гребешках каждой ампулы полукружных каналов и в пятнах мешочков преддверия. * Волосковые рецепторные клетки в апикальной части содержат 40 -110 неподвижных волосков (стереоцилии) и одну подвижную ресничку (киноцилия), расположенную на периферии пучка стереоцилий. * Самые длинные стереоцилии находятся вблизи киноцилии, а длина остальных уменьшается по мере удаления от киноцилии. * Волосковые клетки чувствительны к направлению действия стимула (дирекционная чувствительность). * При направлении раздражающего воздействия от стереоцилий к киноцилии волосковая клетка возбуждается (происходит деполяризация). При противоположном направлении стимула происходит угнетение ответа (гиперполяризация). *
* В эпителии пятен и гребешков находятся чувствительные волосковые и поддерживающие клетки. * Эпителий пятен покрыт студенистой отолитовой мембраной, содержащей отолиты - кристаллы карбоната кальция. * Эпителий гребешков окружён желеобразным прозрачным куполом, легко смещающимся при движениях эндолимфы. * Волосковые клетки находятся в гребешках каждой ампулы полукружных каналов и в пятнах мешочков преддверия. * Волосковые рецепторные клетки в апикальной части содержат 40 -110 неподвижных волосков (стереоцилии) и одну подвижную ресничку (киноцилия), расположенную на периферии пучка стереоцилий. * Самые длинные стереоцилии находятся вблизи киноцилии, а длина остальных уменьшается по мере удаления от киноцилии. * Волосковые клетки чувствительны к направлению действия стимула (дирекционная чувствительность). * При направлении раздражающего воздействия от стереоцилий к киноцилии волосковая клетка возбуждается (происходит деполяризация). При противоположном направлении стимула происходит угнетение ответа (гиперполяризация). *
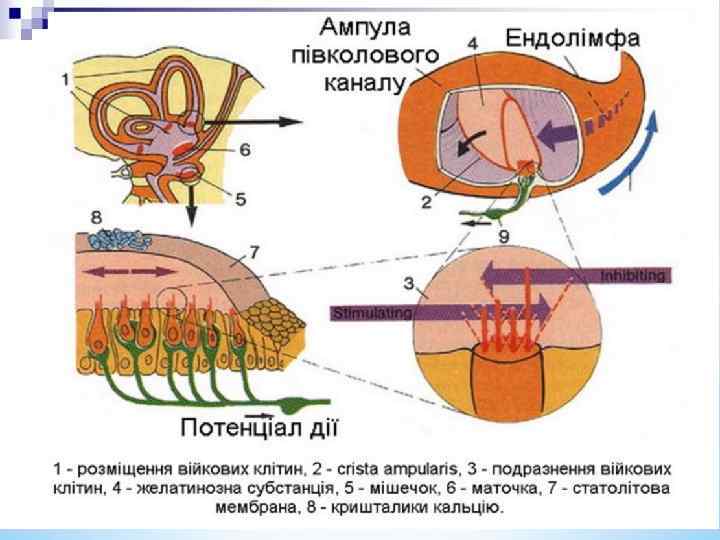
 * Рецепторы полукружных каналов воспринимают ускорение вращения, т. е. углового ускорения. * В состоянии покоя наблюдается баланс частоты нервных импульсов от ампул обеих сторон головы. * Углового ускорения порядка 0, 5 в секунду достаточно для смещения купола и сгибания ресничек. * Угловое ускорение регистрируется благодаря инерции эндолимфы. * При повороте головы эндолимфа остаётся в прежнем положении, а свободный конец купола отклоняется в сторону, противоположную повороту. * Перемещение купола сгибает киноцилию и стереоцилии, внедрённые в желеобразную структуру купола. * Наклон стереоцилий по направлению к киноцилии вызывает деполяризацию и возбуждение; противоположное направление наклона приводит к гиперполяризации и торможению. * При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит выброс ацетилхолина, который активирует афферентные окончания вестибулярного нерва. * и
* Рецепторы полукружных каналов воспринимают ускорение вращения, т. е. углового ускорения. * В состоянии покоя наблюдается баланс частоты нервных импульсов от ампул обеих сторон головы. * Углового ускорения порядка 0, 5 в секунду достаточно для смещения купола и сгибания ресничек. * Угловое ускорение регистрируется благодаря инерции эндолимфы. * При повороте головы эндолимфа остаётся в прежнем положении, а свободный конец купола отклоняется в сторону, противоположную повороту. * Перемещение купола сгибает киноцилию и стереоцилии, внедрённые в желеобразную структуру купола. * Наклон стереоцилий по направлению к киноцилии вызывает деполяризацию и возбуждение; противоположное направление наклона приводит к гиперполяризации и торможению. * При возбуждении в волосковых клетках генерируется рецепторный потенциал и происходит выброс ацетилхолина, который активирует афферентные окончания вестибулярного нерва. * и
 Физиология регистрации углового ускорения. А - различная реакция волосковых клеток в гребешках ампул левого и правого горизонтальных полукружных каналов при повороте головы; Б - последовательно увеличивающиеся изображения воспринимающих структур гребешка.
Физиология регистрации углового ускорения. А - различная реакция волосковых клеток в гребешках ампул левого и правого горизонтальных полукружных каналов при повороте головы; Б - последовательно увеличивающиеся изображения воспринимающих структур гребешка.
 А - иннервация перепончатого лабиринта. Б - вестибулярные волосковые клетки I и II типов.
А - иннервация перепончатого лабиринта. Б - вестибулярные волосковые клетки I и II типов.
 А - Волосковая клетка; Б Положительная механическая деформация; В Отрицательная механическая деформация; Г Механическая чувствительность волосковой клетки; Д Функциональная поляризация вестибулярных волосковых клеток. При сгибании стереоцилий по направлению к киноцилии волосковая клетка деполяризуется и в афферентном волокне возникает возбуждение. При сгибании стереоцилий в сторону от киноцилии волосковая клетка гиперполяризуется и афферентный разряд ослабевает или прекращается
А - Волосковая клетка; Б Положительная механическая деформация; В Отрицательная механическая деформация; Г Механическая чувствительность волосковой клетки; Д Функциональная поляризация вестибулярных волосковых клеток. При сгибании стереоцилий по направлению к киноцилии волосковая клетка деполяризуется и в афферентном волокне возникает возбуждение. При сгибании стереоцилий в сторону от киноцилии волосковая клетка гиперполяризуется и афферентный разряд ослабевает или прекращается
 * Стимуляция полукружных каналов вызывает субъективные ощущения в виде головокружения, тошноты и других реакций, связанных с возбуждением вегетативной нервной системы. *К этому добавляются объективные проявления в виде изменения тонуса глазных мышц (нистагм) и тонуса антигравитационных мышц (реакция падения). * Головокружение является ощущением вращения и может вызвать нарушение равновесия и падение. * Направление ощущения вращения зависит от того, какой полукружный канал был стимулирован. * В каждом случае головокружение ориентировано в направлении, противоположном смещению эндолимфы. * Во время вращения ощущение головокружения направлено в сторону вращения. * Ощущение, испытываемое после прекращения вращения, направлено в сторону, противоположную реальному вращению. *В результате головокружения возникают вегетативные реакции: тошнота, рвота, бледность, потоотделение, а при интенсивной стимуляции полукружных каналов возможно резкое падение АД (коллапс). *
* Стимуляция полукружных каналов вызывает субъективные ощущения в виде головокружения, тошноты и других реакций, связанных с возбуждением вегетативной нервной системы. *К этому добавляются объективные проявления в виде изменения тонуса глазных мышц (нистагм) и тонуса антигравитационных мышц (реакция падения). * Головокружение является ощущением вращения и может вызвать нарушение равновесия и падение. * Направление ощущения вращения зависит от того, какой полукружный канал был стимулирован. * В каждом случае головокружение ориентировано в направлении, противоположном смещению эндолимфы. * Во время вращения ощущение головокружения направлено в сторону вращения. * Ощущение, испытываемое после прекращения вращения, направлено в сторону, противоположную реальному вращению. *В результате головокружения возникают вегетативные реакции: тошнота, рвота, бледность, потоотделение, а при интенсивной стимуляции полукружных каналов возможно резкое падение АД (коллапс). *
 * Стимуляция полукружных каналов вызывает изменения мышечного тонуса, проявляющиеся в нистагме, нарушении координаторных проб и реакции падения. * Нистагм - ритмические подёргивания глаза, состоящие из медленных и быстрых движений. * Медленные движения всегда направлены в сторону движения эндолимфы и являются рефлекторной реакцией. * Рефлекс возникает в гребешках полукружных каналов, импульсы поступают к вестибулярным ядрам ствола мозга и оттуда переключаются к мышцам глаза. * Быстрые движения определяются направлением нистагма; они возникают в результате активности ЦНС (как часть вестибулярного рефлекса из ретикулярной формации в ствол мозга). * Вращение в горизонтальной плоскости вызывает горизонтальный нистагм, вращение в сагиттальной плоскости - вертикальный нистагм, во фронтальной плоскости - вращательный нистагм. *
* Стимуляция полукружных каналов вызывает изменения мышечного тонуса, проявляющиеся в нистагме, нарушении координаторных проб и реакции падения. * Нистагм - ритмические подёргивания глаза, состоящие из медленных и быстрых движений. * Медленные движения всегда направлены в сторону движения эндолимфы и являются рефлекторной реакцией. * Рефлекс возникает в гребешках полукружных каналов, импульсы поступают к вестибулярным ядрам ствола мозга и оттуда переключаются к мышцам глаза. * Быстрые движения определяются направлением нистагма; они возникают в результате активности ЦНС (как часть вестибулярного рефлекса из ретикулярной формации в ствол мозга). * Вращение в горизонтальной плоскости вызывает горизонтальный нистагм, вращение в сагиттальной плоскости - вертикальный нистагм, во фронтальной плоскости - вращательный нистагм. *
 Движение купола и погруженных в него волосков в начале вращения
Движение купола и погруженных в него волосков в начале вращения
 *Нарушение указательной пробы и реакция падения являются результатом изменений тонуса антигравитационных мышц. *Тонус мышц-разгибателей усиливается на стороне тела, куда направлено смещение эндолимфы, противоположной стороне. и ослабевает на * Так, если силы гравитации направлены на правую стопу, то голова и тело человека отклоняются вправо, смещая эндолимфу влево. * Возникший рефлекс немедленно вызовет разгибание правой ноги и руки и сгибание левой руки и ноги, сопровождаемое отклонением глаз влево. *Эти движения рефлексом. * являются защитным выпрямительным
*Нарушение указательной пробы и реакция падения являются результатом изменений тонуса антигравитационных мышц. *Тонус мышц-разгибателей усиливается на стороне тела, куда направлено смещение эндолимфы, противоположной стороне. и ослабевает на * Так, если силы гравитации направлены на правую стопу, то голова и тело человека отклоняются вправо, смещая эндолимфу влево. * Возникший рефлекс немедленно вызовет разгибание правой ноги и руки и сгибание левой руки и ноги, сопровождаемое отклонением глаз влево. *Эти движения рефлексом. * являются защитным выпрямительным
 * Пятно маточки, лежащее горизонтально на нижней её поверхности, реагирует на линейное ускорение в горизонтальном направлении (например, в положении лёжа); * Пятно мешочка, расположенное вертикально на боковой поверхности мешочка, определяет линейное ускорение в вертикальном направлении (например, в положении стоя). * Наклон головы смещает мешочек и маточку на какой-то угол между горизонтальным и вертикальным положением. * Сила тяжести отолитов двигает отолитовую мембрану по отношению к поверхности сенсорного эпителия. * Цилии, внедрённые в отолитовую мембрану, сгибаются под влиянием отолитовой мембраны, скользящей вдоль них. * Если цилии сгибаются в сторону киноцилии, то увеличивается импульсная активность, если в другую сторону от киноцилии, то импульсная активность уменьшается. * Таким образом, функцией мешочка и маточки является поддержание статического равновесия и ориентация головы по отношению к направлению силы тяжести. *
* Пятно маточки, лежащее горизонтально на нижней её поверхности, реагирует на линейное ускорение в горизонтальном направлении (например, в положении лёжа); * Пятно мешочка, расположенное вертикально на боковой поверхности мешочка, определяет линейное ускорение в вертикальном направлении (например, в положении стоя). * Наклон головы смещает мешочек и маточку на какой-то угол между горизонтальным и вертикальным положением. * Сила тяжести отолитов двигает отолитовую мембрану по отношению к поверхности сенсорного эпителия. * Цилии, внедрённые в отолитовую мембрану, сгибаются под влиянием отолитовой мембраны, скользящей вдоль них. * Если цилии сгибаются в сторону киноцилии, то увеличивается импульсная активность, если в другую сторону от киноцилии, то импульсная активность уменьшается. * Таким образом, функцией мешочка и маточки является поддержание статического равновесия и ориентация головы по отношению к направлению силы тяжести. *
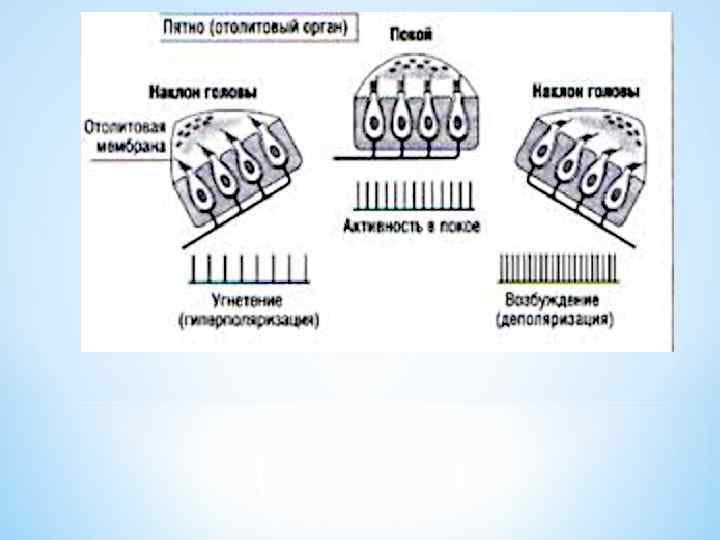
 * Пятна маточки и мешочка участвуют также в определении линейного ускорения. * Когда человек неожиданно получает толчок вперёд (ускорение), то отолитовая мембрана, имеющая инерционность намного больше, чем окружающая жидкость, смещается назад на цилии волосковой клетки. * Вследствие этого в нервную систему сигнал поступает о нарушении равновесия тела, и человек чувствует, что он падает назад. * Автоматически человек наклоняется вперёд до тех пор, пока это движение не вызовет такое же ощущение падения вперёд, потому что отолитовая мембрана под влиянием ускорения возвращается на своё место. * В этой точке нервная система определяет состояние подходящего равновесия и прекращает наклон тела вперёд. * Следовательно, пятна управляют поддержанием равновесия во время линейного ускорения. *
* Пятна маточки и мешочка участвуют также в определении линейного ускорения. * Когда человек неожиданно получает толчок вперёд (ускорение), то отолитовая мембрана, имеющая инерционность намного больше, чем окружающая жидкость, смещается назад на цилии волосковой клетки. * Вследствие этого в нервную систему сигнал поступает о нарушении равновесия тела, и человек чувствует, что он падает назад. * Автоматически человек наклоняется вперёд до тех пор, пока это движение не вызовет такое же ощущение падения вперёд, потому что отолитовая мембрана под влиянием ускорения возвращается на своё место. * В этой точке нервная система определяет состояние подходящего равновесия и прекращает наклон тела вперёд. * Следовательно, пятна управляют поддержанием равновесия во время линейного ускорения. *
 *Вестибулярная ветвь VIII черепного нерва образована отростками примерно 19 тыс. биполярных нейронов, составляющих чувствительный ганглий. *Периферические отростки нейронов подходят к волосковым клеткам каждого полукружного канала, маточки и мешочка, а центральные отростки направляются в вестибулярные ядра продолговатого мозга. *Аксоны нервных клеток второго порядка связаны со спинным мозгом (преддверно-спинномозговой путь, оливоспинномозговой путь) и поднимаются в составе медиальных продольных пучков к двигательным ядрам черепных нервов, осуществляющих контроль за движениями глаза. *Имеется также путь, проводящий импульсы от вестибулярных рецепторов через таламус к коре больших полушарий мозга. *
*Вестибулярная ветвь VIII черепного нерва образована отростками примерно 19 тыс. биполярных нейронов, составляющих чувствительный ганглий. *Периферические отростки нейронов подходят к волосковым клеткам каждого полукружного канала, маточки и мешочка, а центральные отростки направляются в вестибулярные ядра продолговатого мозга. *Аксоны нервных клеток второго порядка связаны со спинным мозгом (преддверно-спинномозговой путь, оливоспинномозговой путь) и поднимаются в составе медиальных продольных пучков к двигательным ядрам черепных нервов, осуществляющих контроль за движениями глаза. *Имеется также путь, проводящий импульсы от вестибулярных рецепторов через таламус к коре больших полушарий мозга. *
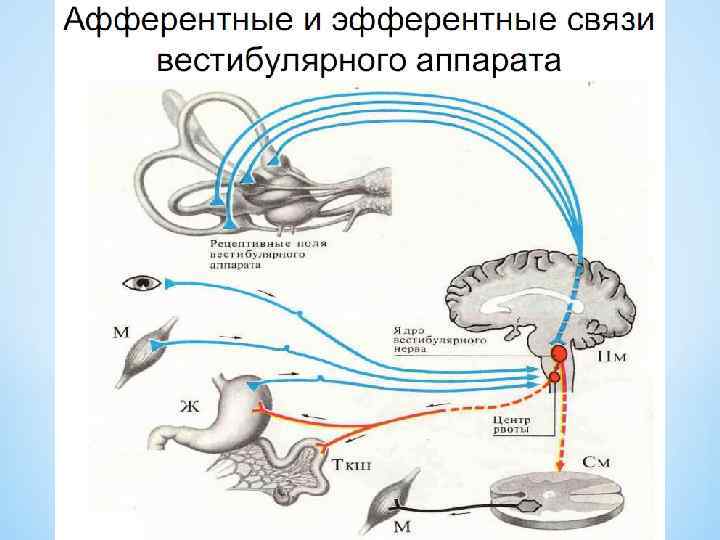
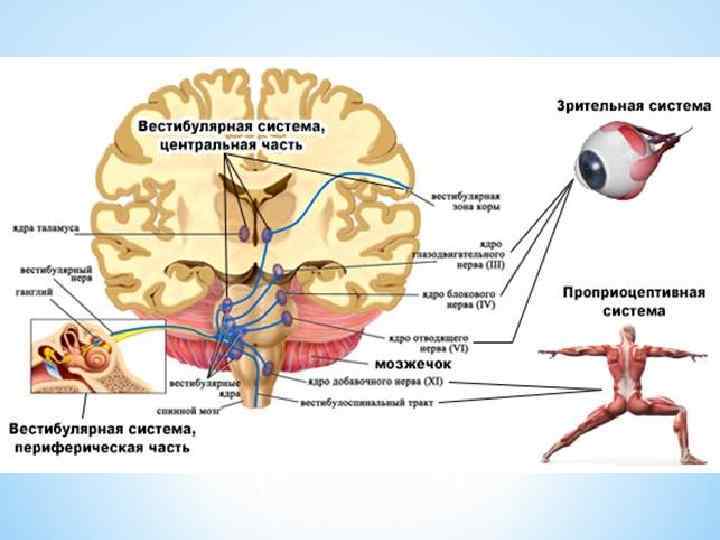
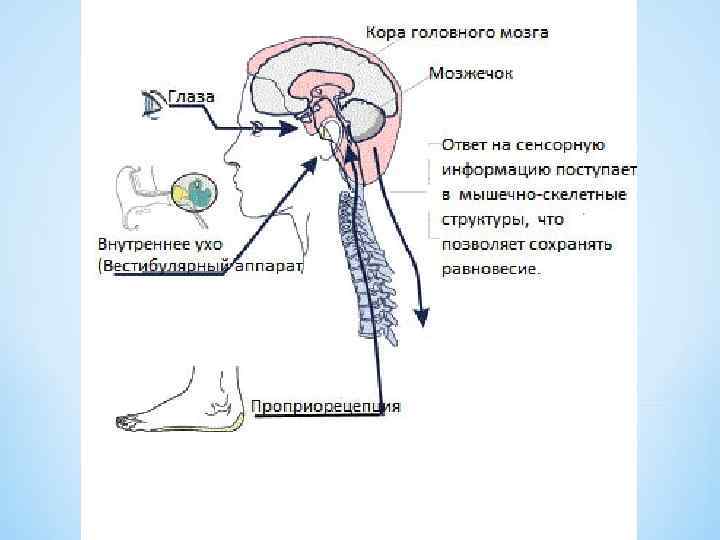
 *Мультимодальная система включает зрительные и соматические рецепторы, которые посылают сигналы к вестибулярным ядрам либо непосредственно, либо через вестибулярные ядра мозжечка или ретикулярную формацию. *Входящие сигналы интегрируются в вестибулярных ядрах, и выходящие команды воздействуют на глазодвигательные и спинальные системы моторного контроля. *Центральная и координирующая роль вестибулярных ядер соединение прямыми и обратными связями с основными рецепторными и центральными системами пространственной координации. *
*Мультимодальная система включает зрительные и соматические рецепторы, которые посылают сигналы к вестибулярным ядрам либо непосредственно, либо через вестибулярные ядра мозжечка или ретикулярную формацию. *Входящие сигналы интегрируются в вестибулярных ядрах, и выходящие команды воздействуют на глазодвигательные и спинальные системы моторного контроля. *Центральная и координирующая роль вестибулярных ядер соединение прямыми и обратными связями с основными рецепторными и центральными системами пространственной координации. *
 *Равновесие тела — состояние покоя тела относительно какойлибо системы отсчета, в частном случае — неподвижность тела относительно окружающей его среды. Равновесие тела бывает статическим и динамическим. При статическом равновесии тела проекция общего центра тяжести тела находится внутри площади опоры (рис. 1). При ходьбе, беге, катании на коньках и т. п. динамическое равновесие тела достигается путем балансирования, т. е. подведением площади опоры под сместившуюся проекцию центра тяжести тела (рис. 2). Рис. 1. Плоскость опоры тела при вольном стоянии: S — точка, соответствующая проекции общего центра тяжести. Рис. 2. Восстановление равновесия тела путем компенсирующего изменения деятельности мышц ног при наклоне туловища вперед. Проекция центра тяжести (вертикальная линия) возвращена в прежнее положение на плоскость опоры.
*Равновесие тела — состояние покоя тела относительно какойлибо системы отсчета, в частном случае — неподвижность тела относительно окружающей его среды. Равновесие тела бывает статическим и динамическим. При статическом равновесии тела проекция общего центра тяжести тела находится внутри площади опоры (рис. 1). При ходьбе, беге, катании на коньках и т. п. динамическое равновесие тела достигается путем балансирования, т. е. подведением площади опоры под сместившуюся проекцию центра тяжести тела (рис. 2). Рис. 1. Плоскость опоры тела при вольном стоянии: S — точка, соответствующая проекции общего центра тяжести. Рис. 2. Восстановление равновесия тела путем компенсирующего изменения деятельности мышц ног при наклоне туловища вперед. Проекция центра тяжести (вертикальная линия) возвращена в прежнее положение на плоскость опоры.
; мультимодальная") Восходящие пути вестибулярного аппарата, вид сзади. Мозжечок и кора больших полушарий удалены (А); мультимодальная система пространственной ориентации тела (Б).
Восходящие пути вестибулярного аппарата, вид сзади. Мозжечок и кора больших полушарий удалены (А); мультимодальная система пространственной ориентации тела (Б).
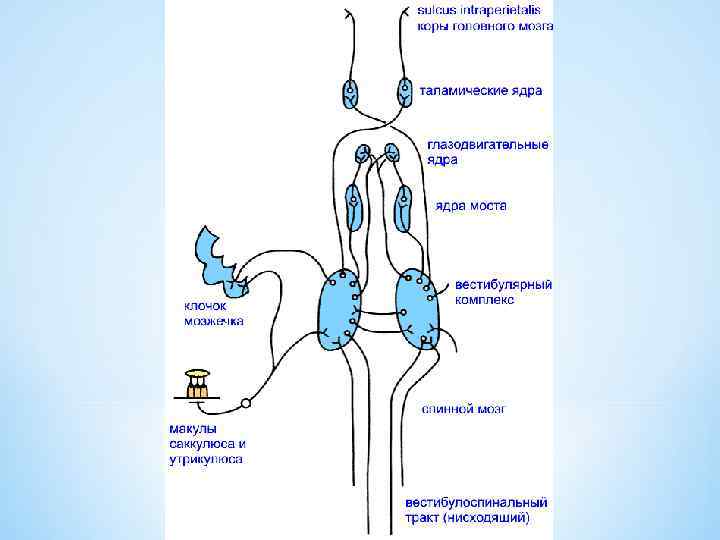
 Проводящие пути импульсов вестибулярной чувствительности. 1 — преддверно-спинномозговой путь; 2 — полукружные протоки; 3 — преддверный узел; 4 — преддверный корешок; 5 — нижнее вестибулярное ядро; 6 — медиальное вестибулярное ядро; 7 — латеральное вестибулярное ядро; 8 — верхнее вестибуляр ное ядро; 9 — ядро шатра мозжечка; 10 — зубчатое ядро мозжечка; 11 — медиальный продольный пучок; 12 — ядро отводящего нерва; 13 — ретикулярная формация; 14 — верхняя мозжечковая ножка; 15 — красное ядро; 16 — ядро глазодвигательного нерва; 17 — ядро Даркшевича; 18 — чечевицеобразное ядро; 19 — таламус; 20 — кора темен ной доли; 21 — кора височной доли большого полушария мозга.
Проводящие пути импульсов вестибулярной чувствительности. 1 — преддверно-спинномозговой путь; 2 — полукружные протоки; 3 — преддверный узел; 4 — преддверный корешок; 5 — нижнее вестибулярное ядро; 6 — медиальное вестибулярное ядро; 7 — латеральное вестибулярное ядро; 8 — верхнее вестибуляр ное ядро; 9 — ядро шатра мозжечка; 10 — зубчатое ядро мозжечка; 11 — медиальный продольный пучок; 12 — ядро отводящего нерва; 13 — ретикулярная формация; 14 — верхняя мозжечковая ножка; 15 — красное ядро; 16 — ядро глазодвигательного нерва; 17 — ядро Даркшевича; 18 — чечевицеобразное ядро; 19 — таламус; 20 — кора темен ной доли; 21 — кора височной доли большого полушария мозга.
 Связи вестибулярных нервов через вестибулярные ядра с другими областями ЦНС
Связи вестибулярных нервов через вестибулярные ядра с другими областями ЦНС