

скелет_мышцы.ppt
- Количество слайдов: 34
 Скелетная мышца 1. Строение мышцы 2. Структура и состав саркомера 3. Механизм мышечного сокращения 4. Типы мышечных волокон 5. Механика мышечной активности 6. Двигательные единицы 7. Утомление в нервно-мышечном аппарате
Скелетная мышца 1. Строение мышцы 2. Структура и состав саркомера 3. Механизм мышечного сокращения 4. Типы мышечных волокон 5. Механика мышечной активности 6. Двигательные единицы 7. Утомление в нервно-мышечном аппарате
 полоски сарколемма") надкостница, покрывающая кость сухожилие фасция скелетная мышца пучок волокон мышечное волокно (клетка) полоски сарколемма саркоплазма филаменты ядро миофибрилла
надкостница, покрывающая кость сухожилие фасция скелетная мышца пучок волокон мышечное волокно (клетка) полоски сарколемма саркоплазма филаменты ядро миофибрилла
 ядро сарколемма саркоплазма миофиламенты миофибрилы полоски") мышечное волокно (клетка) ядро сарколемма саркоплазма миофиламенты миофибрилы полоски
мышечное волокно (клетка) ядро сарколемма саркоплазма миофиламенты миофибрилы полоски
 (анизотропная, более темная) толстый филамент тонкие филаменты") Структура саркомера (изотропная, светлая) (анизотропная, более темная) толстый филамент тонкие филаменты
Структура саркомера (изотропная, светлая) (анизотропная, более темная) толстый филамент тонкие филаменты
 Молекула миозина 6 полипептидных цепей: 4 легкие + 2 тяжелые головки тяжелых цепей миозина регуляторные легкая цепь существенная легкая цепь Шарнирный регион Хвостовой регион тяжелых цепей Полимеризация молекул миозина в толстый филамент Биполярный филамент
Молекула миозина 6 полипептидных цепей: 4 легкие + 2 тяжелые головки тяжелых цепей миозина регуляторные легкая цепь существенная легкая цепь Шарнирный регион Хвостовой регион тяжелых цепей Полимеризация молекул миозина в толстый филамент Биполярный филамент
 нить F-актина (тонкий филамент)") актин-мономер (G) нить F-актина (тонкий филамент)
актин-мономер (G) нить F-актина (тонкий филамент)
 Головка миозина -от 7 до 50 гребков в секунду в зависимости от типа миозина
Головка миозина -от 7 до 50 гребков в секунду в зависимости от типа миозина
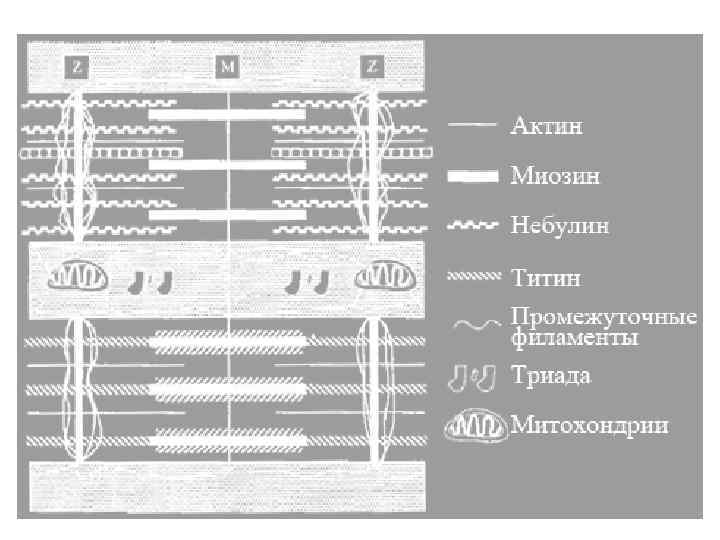
 α-актинин Миозин Актин Титин линия зона полоса
α-актинин Миозин Актин Титин линия зона полоса
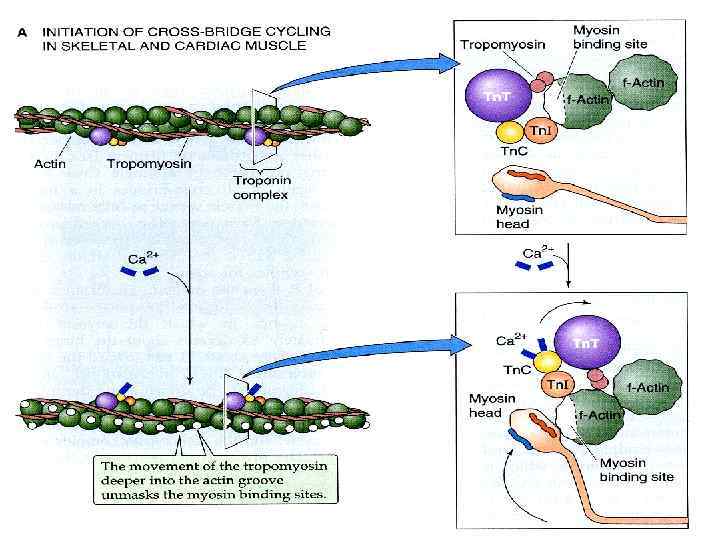
 F-актин Тропониновый комплекс (Tn)") Актин, тропонин и тропомиозин Са 2+ (связан с Тn. C) F-актин Тропониновый комплекс (Tn) тропомиозин сайт (участок), связывающий миозин
Актин, тропонин и тропомиозин Са 2+ (связан с Тn. C) F-актин Тропониновый комплекс (Tn) тропомиозин сайт (участок), связывающий миозин
 Участки, взаимодействия с миозином, закрыты тропомиозином Участки, взаимодействия с миозином, доступны для головки миозина
Участки, взаимодействия с миозином, закрыты тропомиозином Участки, взаимодействия с миозином, доступны для головки миозина
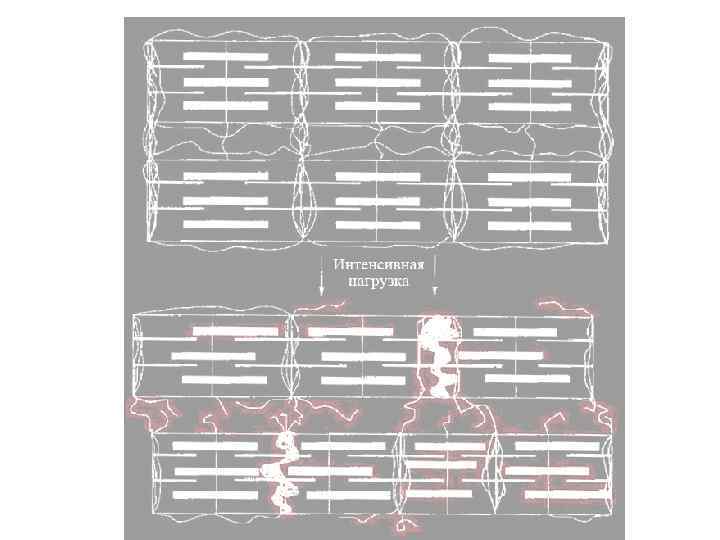
 Скольжение миофиламентов при сокращении
Скольжение миофиламентов при сокращении
 Тренировка ведет к увеличению числа ядер в мышечном волокне. Мышечное волокно становится толще. После прекращения тренировки мышца атрофируется, но число ядер остается прежним. При возобновлении тренировки мышца быстро наращивает свой объем
Тренировка ведет к увеличению числа ядер в мышечном волокне. Мышечное волокно становится толще. После прекращения тренировки мышца атрофируется, но число ядер остается прежним. При возобновлении тренировки мышца быстро наращивает свой объем
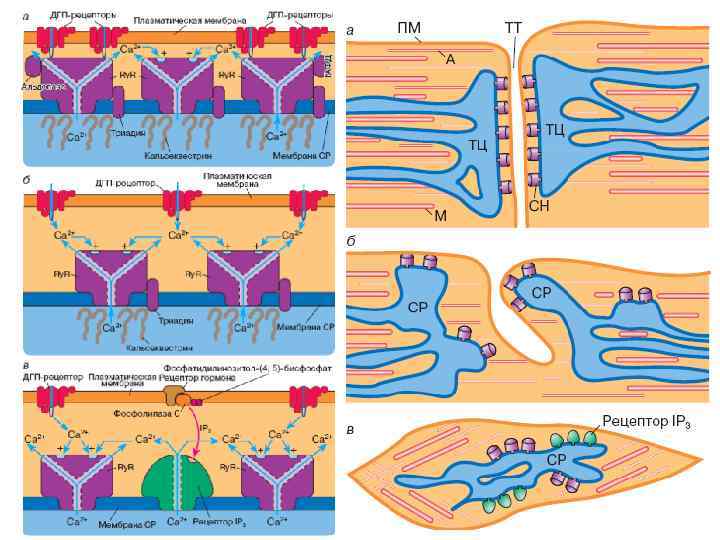
 Электро-механическое сопряжение Поперечная трубочка СПР
Электро-механическое сопряжение Поперечная трубочка СПР
 Триада: Цистерна СПР диск поперечная трубочка Инвагинации Цистерна СПР в") миофибрилла Плазматическая мембрана (сарколемма) Триада: Цистерна СПР диск поперечная трубочка Инвагинации Цистерна СПР в мембране (поперечные трубочки) диск Граница А- и I- полосок Саркоплазматический ретикулум
миофибрилла Плазматическая мембрана (сарколемма) Триада: Цистерна СПР диск поперечная трубочка Инвагинации Цистерна СПР в мембране (поперечные трубочки) диск Граница А- и I- полосок Саркоплазматический ретикулум
 Са 2+ Активаторы: Са 2+ (мк. М), АТФ, кофеин,") Рианодиновый рецептор (Cа освобождающий канал) Са 2+ Активаторы: Са 2+ (мк. М), АТФ, кофеин, полиамины, ионы тяжелых металлов Са 2+ Кальциевый канал L-типа (дигидропиридиновый рецептор) Блокаторы: Са 2+ (м. М), рианодин (10 -8 М), прокаин, . Са 2+ Блокаторы: этиловый спирт, Сd 2+, ω-конотоксин, верапамил
Рианодиновый рецептор (Cа освобождающий канал) Са 2+ Активаторы: Са 2+ (мк. М), АТФ, кофеин, полиамины, ионы тяжелых металлов Са 2+ Кальциевый канал L-типа (дигидропиридиновый рецептор) Блокаторы: Са 2+ (м. М), рианодин (10 -8 М), прокаин, . Са 2+ Блокаторы: этиловый спирт, Сd 2+, ω-конотоксин, верапамил
 Мембранная деполяризация открывает Са каналы L-типа Механическое сопряжение между Саканалом и рианодиновым рецептором, ведет к открытию последнего Са может также входить через канал L-типа и активировать рианодиновые рецепторы, но в скелетной мышце этот путь не имеет решающего значения. Са выходит из ретикулума и активирует тропонин С, что ведет к сокращению
Мембранная деполяризация открывает Са каналы L-типа Механическое сопряжение между Саканалом и рианодиновым рецептором, ведет к открытию последнего Са может также входить через канал L-типа и активировать рианодиновые рецепторы, но в скелетной мышце этот путь не имеет решающего значения. Са выходит из ретикулума и активирует тропонин С, что ведет к сокращению
 / SERCA 80% всех белков в СПР мкс мс АДФ АТФ") Са АТФаза (насос) / SERCA 80% всех белков в СПР мкс мс АДФ АТФ P Са 2+ 500 -1000 н. М P Кальмодулин (САМ) Cа Са 2+ / кальмодулин -зависимая протеинкиназа 2+ ФОСФОЛАМБАН
Са АТФаза (насос) / SERCA 80% всех белков в СПР мкс мс АДФ АТФ P Са 2+ 500 -1000 н. М P Кальмодулин (САМ) Cа Са 2+ / кальмодулин -зависимая протеинкиназа 2+ ФОСФОЛАМБАН
") Механизмы удаления Са 2+ из цитоплазмы Na-Cа обменник и Са-насос плазматической мембраны (в кардиомиоцитах) Са –насос саркоплазматического ретикулума Са связывающие белки в саркоплазматическом ретикулуме (буферы ионов Са) кальретикулин кальсеквестрин
Механизмы удаления Са 2+ из цитоплазмы Na-Cа обменник и Са-насос плазматической мембраны (в кардиомиоцитах) Са –насос саркоплазматического ретикулума Са связывающие белки в саркоплазматическом ретикулуме (буферы ионов Са) кальретикулин кальсеквестрин
 Сокращения мышечных волокон Тонические Фазные В ответ на раздражение – максимальное сокращение «слабый» стимул Амплитуда сокращений зависит от силы стимуляции «сильный» стимул сила «слабый» стимул «сильный» стимул сила длительность фазные волокна быстрые медленные характерно для скелетных мышц длительность контрактура -обратимое медленное устойчивое сокращение при отсутствии распространяющегося ПД характерно для гладкомышечных клеток, а также глазных мышц (мультитерминальная иннервация)
Сокращения мышечных волокон Тонические Фазные В ответ на раздражение – максимальное сокращение «слабый» стимул Амплитуда сокращений зависит от силы стимуляции «сильный» стимул сила «слабый» стимул «сильный» стимул сила длительность фазные волокна быстрые медленные характерно для скелетных мышц длительность контрактура -обратимое медленное устойчивое сокращение при отсутствии распространяющегося ПД характерно для гладкомышечных клеток, а также глазных мышц (мультитерминальная иннервация)
 Изотонический тип сокращения покой активность - при небольшой величине груза - длинная мышца укорачивается быстрее, чем короткая t s 1 Постоянное натяжение вес t s 2 вес V=s / t, V 1 > V 2 скорость
Изотонический тип сокращения покой активность - при небольшой величине груза - длинная мышца укорачивается быстрее, чем короткая t s 1 Постоянное натяжение вес t s 2 вес V=s / t, V 1 > V 2 скорость
 Изометрический тип сокращения покой активность нагрузка неподъемным грузом или статическая нагрузка Постоянная длинна Сила натяжения ~ начальной длинны: оптимальное перекрывание филаментов слабое натяжение слабо растянутая мышца сильное натяжение
Изометрический тип сокращения покой активность нагрузка неподъемным грузом или статическая нагрузка Постоянная длинна Сила натяжения ~ начальной длинны: оптимальное перекрывание филаментов слабое натяжение слабо растянутая мышца сильное натяжение
 ОМС, суммация, тетанус - регуляция силы сокращения частотой стимуляции 5 Гц одиночные сокращения 10 Гц суммация 15 -25 Гц зубчатый тетанус (неполный) 25 Гц и более гладкий тетанус
ОМС, суммация, тетанус - регуляция силы сокращения частотой стимуляции 5 Гц одиночные сокращения 10 Гц суммация 15 -25 Гц зубчатый тетанус (неполный) 25 Гц и более гладкий тетанус
 Типы скелетных мышечных волокон быстрые медленные Тип II ени тр Тип II a -красные (миоглобин) ка ров Устойчивые к утомлению Тип I Сила Утомляемые Тип II b стайер -оксидативный метаболизм - митохондрии - гликогена мало марафон - белые -оксидативный метаболизм - гликолиз - митохондрии мало - митохондрии - гликоген -красные (миоглобин) - гликоген медленное волокно быстрое волокно спринт Время, милисекунды 40% медленных – 60 % быстрых у европейца Тренировка – увеличивает только толщину волокон, но не их количество
Типы скелетных мышечных волокон быстрые медленные Тип II ени тр Тип II a -красные (миоглобин) ка ров Устойчивые к утомлению Тип I Сила Утомляемые Тип II b стайер -оксидативный метаболизм - митохондрии - гликогена мало марафон - белые -оксидативный метаболизм - гликолиз - митохондрии мало - митохондрии - гликоген -красные (миоглобин) - гликоген медленное волокно быстрое волокно спринт Время, милисекунды 40% медленных – 60 % быстрых у европейца Тренировка – увеличивает только толщину волокон, но не их количество
 Экспрессия сократительных и регуляторных белков медленные мах АТФазная активность сарколипин
Экспрессия сократительных и регуляторных белков медленные мах АТФазная активность сарколипин
 быстрое утомляемое волокно 50 g медленное волокно быстрое устойчивое волокно 10 g 2 g 25 Гц 12 Гц 20 Гц быстро утомляется практически не утомляется 6 мин 2 мин
быстрое утомляемое волокно 50 g медленное волокно быстрое устойчивое волокно 10 g 2 g 25 Гц 12 Гц 20 Гц быстро утомляется практически не утомляется 6 мин 2 мин
 двигательная единица «все или ничего» пул мотонейронов мышечные волокна чем больше") Нейро-моторная единица (двигательная) двигательная единица «все или ничего» пул мотонейронов мышечные волокна чем больше синхронность работы, тем больше сила сокращения мышцы чем больше мотонейронов активны, тем сильнее сокращение мышца, управляющая «точными» движениями: пул из 1500 -3000 нейронов, 1 МН – 8 -50 волокон. (высокая плотность иннервации) мышца, управляющая «грубыми» движениями: пул из 600 – 1600 нейронов, 1 МН – 600 -2000 волокон (низкая плотность иннервации) 85% у спортсмена, 60% у нетренированного ДЕ вовлекаются в движение
Нейро-моторная единица (двигательная) двигательная единица «все или ничего» пул мотонейронов мышечные волокна чем больше синхронность работы, тем больше сила сокращения мышцы чем больше мотонейронов активны, тем сильнее сокращение мышца, управляющая «точными» движениями: пул из 1500 -3000 нейронов, 1 МН – 8 -50 волокон. (высокая плотность иннервации) мышца, управляющая «грубыми» движениями: пул из 600 – 1600 нейронов, 1 МН – 600 -2000 волокон (низкая плотность иннервации) 85% у спортсмена, 60% у нетренированного ДЕ вовлекаются в движение
 Варианты двигательных единиц низко- быстрые утомляемые высоковозбудимый нейрон быстрые, устойчивые к утомлению Медленная ДЕ медленные Быстрая ДЕ Процент от максимальной силы - регуляция силы сокращения количеством вовлеченных волокон прыжок галлоп бег прогулка стояние Процент участвующих мотонейронов Чем меньше тело нейрона, тем больше возбудимость и включенность в рефлекторные акты
Варианты двигательных единиц низко- быстрые утомляемые высоковозбудимый нейрон быстрые, устойчивые к утомлению Медленная ДЕ медленные Быстрая ДЕ Процент от максимальной силы - регуляция силы сокращения количеством вовлеченных волокон прыжок галлоп бег прогулка стояние Процент участвующих мотонейронов Чем меньше тело нейрона, тем больше возбудимость и включенность в рефлекторные акты
 Свойства двигательных единиц IIВ IIА I Максимальная сила Тип Медленные медленные быстрые распределение Быстрые Частота работы ДЕ, Гц
Свойства двигательных единиц IIВ IIА I Максимальная сила Тип Медленные медленные быстрые распределение Быстрые Частота работы ДЕ, Гц
 синаптическая задержка синапс (100 Гц) 2 мс мышечное волокно") ЛАБИЛЬНОСТЬ мотонейрон аксон (500 Гц) синаптическая задержка синапс (100 Гц) 2 мс мышечное волокно (200 Гц) 10 мс 5 мс
ЛАБИЛЬНОСТЬ мотонейрон аксон (500 Гц) синаптическая задержка синапс (100 Гц) 2 мс мышечное волокно (200 Гц) 10 мс 5 мс
 Утомление в нервно-мышечном синапсе синаптическая депрессия механизмы пресинаптический постсинаптический уменьшение выброса медиатора снижение чувствительности к медиатору -десенситизация Ах. Р β 2 -АР - снижение активности машины слияния -уменьшение вероятности открытия Са 2+ -канала -замедление доставки синаптических везикул в активную зону Катехоловые амины -истощение запасов синаптических везикул -увеличивает активность Са-АТФазы, -ускоряет распад гликогена синтез АТФ -облегчает выброс Са 2+ из СПР расширение сосудов мышц -стимулирует синтез NO -гипертрофическое действие на мышечные волокна
Утомление в нервно-мышечном синапсе синаптическая депрессия механизмы пресинаптический постсинаптический уменьшение выброса медиатора снижение чувствительности к медиатору -десенситизация Ах. Р β 2 -АР - снижение активности машины слияния -уменьшение вероятности открытия Са 2+ -канала -замедление доставки синаптических везикул в активную зону Катехоловые амины -истощение запасов синаптических везикул -увеличивает активность Са-АТФазы, -ускоряет распад гликогена синтез АТФ -облегчает выброс Са 2+ из СПР расширение сосудов мышц -стимулирует синтез NO -гипертрофическое действие на мышечные волокна