

регуляция активности генов 2015.ppt
- Количество слайдов: 49
 Синтез и. РНК у прокариот
Синтез и. РНК у прокариот
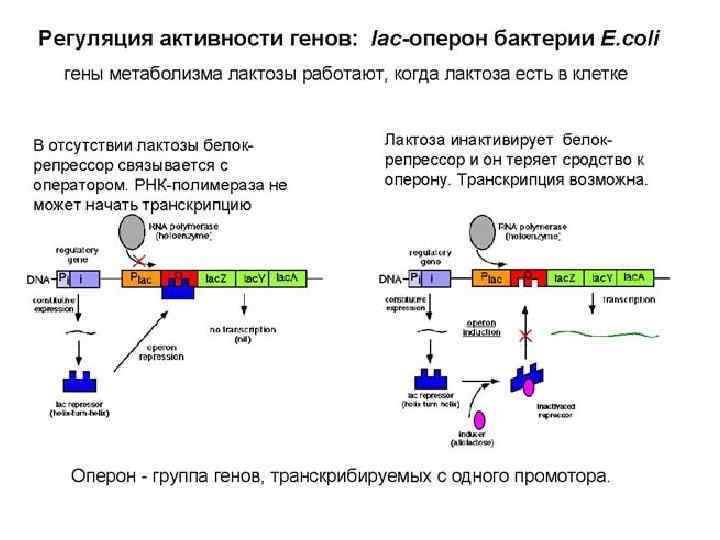
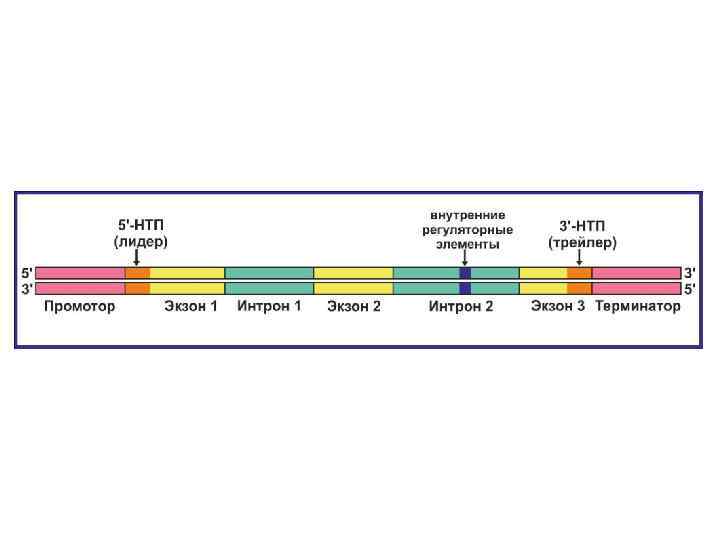
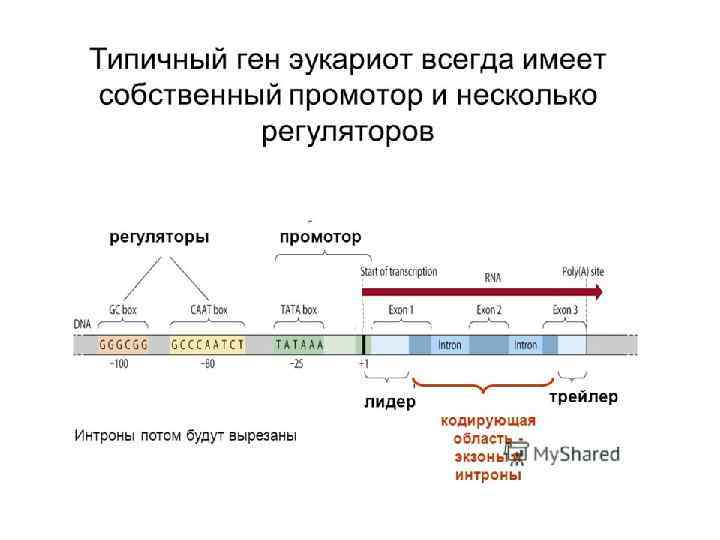
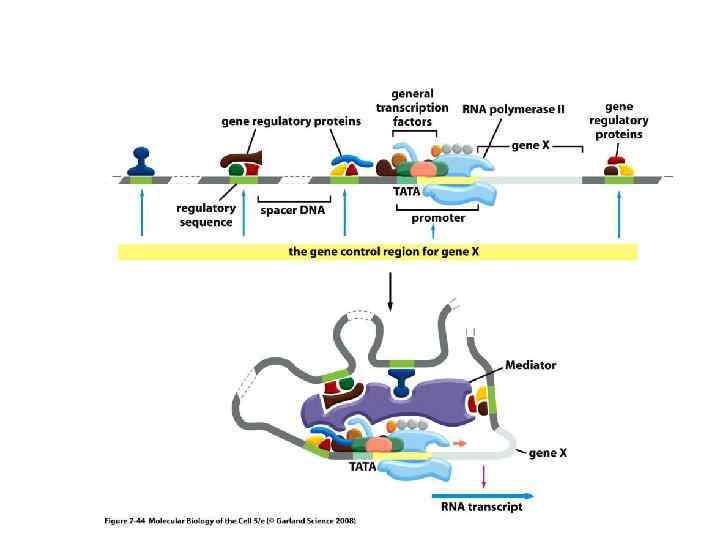
 использует S-аденозиметионин (S-adenosyl methionine, SAM) как донор") ДНК метилтрансфераза (DNA methyltransferase, DNA Mtase, DNMT) использует S-аденозиметионин (S-adenosyl methionine, SAM) как донор метильной группы.
ДНК метилтрансфераза (DNA methyltransferase, DNA Mtase, DNMT) использует S-аденозиметионин (S-adenosyl methionine, SAM) как донор метильной группы.
 • • При оплодотворении зигота наследует одну родительскую хромосому с гаметическим метилированием ДНК (красные вертикальные полосы), а также с метилированием ДНК, которое не поддерживается в клетках эмбриона (серые столбцы). Другая хромосома не метилируется на стадии гаметы. Эмбриональные клетки распределяются на клетки сомы, которые реметилируются и клетки зародышевой линии, где метилирование снимается аллель-специфическим образом. Shirley M. Tilghman The Sins of the Fathers and Mothers: Genomic Imprinting in Mammalian Development Cell, Vol. 96, 185– 193, January 22, 1999
• • При оплодотворении зигота наследует одну родительскую хромосому с гаметическим метилированием ДНК (красные вертикальные полосы), а также с метилированием ДНК, которое не поддерживается в клетках эмбриона (серые столбцы). Другая хромосома не метилируется на стадии гаметы. Эмбриональные клетки распределяются на клетки сомы, которые реметилируются и клетки зародышевой линии, где метилирование снимается аллель-специфическим образом. Shirley M. Tilghman The Sins of the Fathers and Mothers: Genomic Imprinting in Mammalian Development Cell, Vol. 96, 185– 193, January 22, 1999
 • Эпигенетическое перепрограммирование генома у мышей включает в себя две фазы деметилирования ДНК. Первая волна деметилирования ДНК происходит у самцов (синяя кривая) или самок (красная кривая) при образовании первичных (primordial)зародышевых клеток (1). Затем происходит метилирование ДНК на стадии образования гамет, причем позже в яйцеклетке (стадия овуляции), чем в сперматозоидах, это происходит по всему геному, в том числе в импринтированных генах (2). ). Вторая волна деметилирования ДНК происходит после оплодотворения зиготы в F 2 (3). Наблюдается более быстрым деметилирования в отцовском, чем в материнском геноме. Пунктирные кривые обозначают сохранение прежнего уровня метилирования, что делает возможным эффекты «материнского» и «отцовского» наследования (импринтинга). Реметилирование обоих родительских геномов происходит примерно на стадии имплантации эмбриона (4). Frésard et al. Genetics Selection Evolution 2013 45: 16 doi: 10. 1186/1297 -9686 -45 -16
• Эпигенетическое перепрограммирование генома у мышей включает в себя две фазы деметилирования ДНК. Первая волна деметилирования ДНК происходит у самцов (синяя кривая) или самок (красная кривая) при образовании первичных (primordial)зародышевых клеток (1). Затем происходит метилирование ДНК на стадии образования гамет, причем позже в яйцеклетке (стадия овуляции), чем в сперматозоидах, это происходит по всему геному, в том числе в импринтированных генах (2). ). Вторая волна деметилирования ДНК происходит после оплодотворения зиготы в F 2 (3). Наблюдается более быстрым деметилирования в отцовском, чем в материнском геноме. Пунктирные кривые обозначают сохранение прежнего уровня метилирования, что делает возможным эффекты «материнского» и «отцовского» наследования (импринтинга). Реметилирование обоих родительских геномов происходит примерно на стадии имплантации эмбриона (4). Frésard et al. Genetics Selection Evolution 2013 45: 16 doi: 10. 1186/1297 -9686 -45 -16
 • LCR – locus control region – регуляторные последовательности в геноме эукариот, которые регулируют активность генов в кластерах (например, гены глобинов).
• LCR – locus control region – регуляторные последовательности в геноме эукариот, которые регулируют активность генов в кластерах (например, гены глобинов).
 • LCR в кластере генов β-глобина
• LCR в кластере генов β-глобина
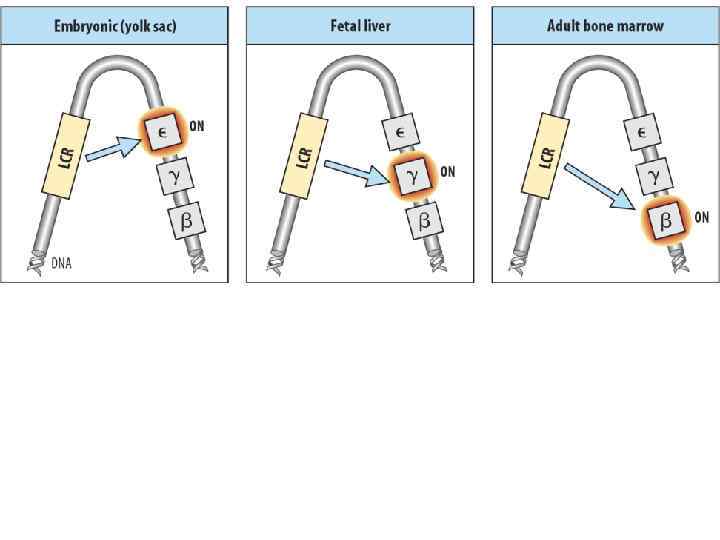
 Схема изменений в состоянии фосфорилирования большой субъединицы РНК-полимеразы -II процессе транскрипции Большая субъединица ( RPB 1) полимеразы RNAP II имеет C-терминальную область (CTD), состоящую из 52 тандемных серинбогатых повторов, которые могут быть посттрансяционно фосфорилированы. Изначально RNAP II нефосфорилирована, затем, в процессе инициации Ser 5 фосфорилируется с помощью фактора транскрипции TFIIH. В процессе элонгации наблюдается Фосфорилирование серина Ser 2 киназой CTDKI, которая является частью фактора элонгации PЌTEFb. На завершающем этапе элонгации происходит дефосфорилирование по Ser 5 с помощью протеинфосфатазы. Фосфорилирование Ser 7 необходимо для транскрипции малых ядерных РНК.
Схема изменений в состоянии фосфорилирования большой субъединицы РНК-полимеразы -II процессе транскрипции Большая субъединица ( RPB 1) полимеразы RNAP II имеет C-терминальную область (CTD), состоящую из 52 тандемных серинбогатых повторов, которые могут быть посттрансяционно фосфорилированы. Изначально RNAP II нефосфорилирована, затем, в процессе инициации Ser 5 фосфорилируется с помощью фактора транскрипции TFIIH. В процессе элонгации наблюдается Фосфорилирование серина Ser 2 киназой CTDKI, которая является частью фактора элонгации PЌTEFb. На завершающем этапе элонгации происходит дефосфорилирование по Ser 5 с помощью протеинфосфатазы. Фосфорилирование Ser 7 необходимо для транскрипции малых ядерных РНК.
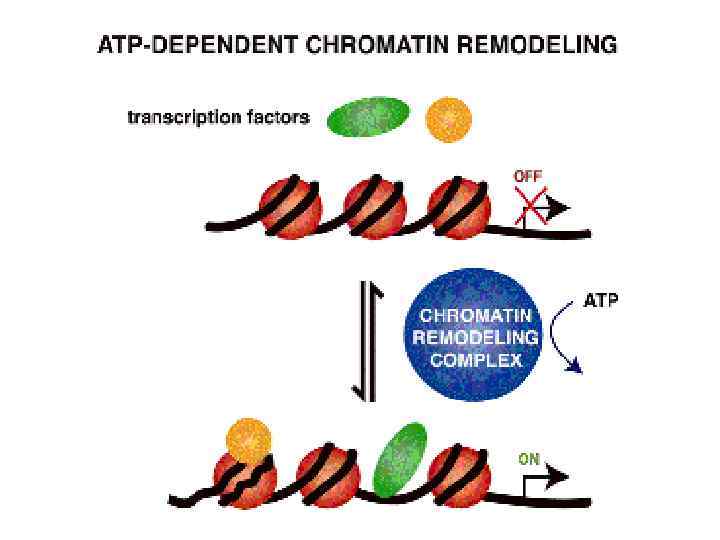
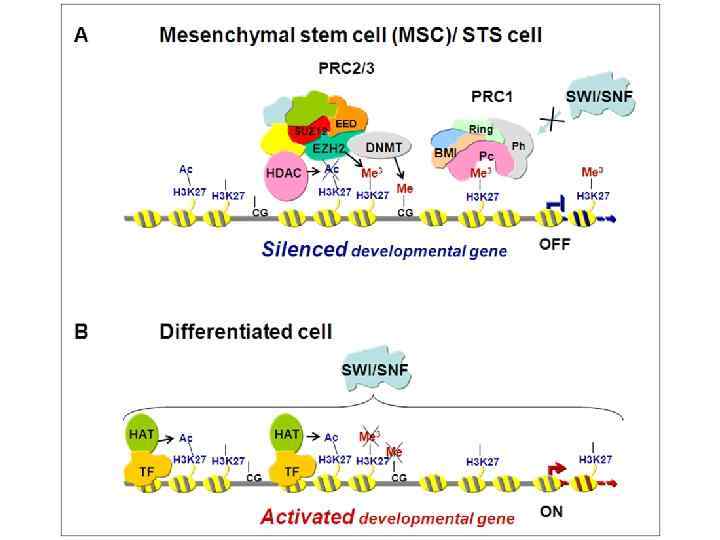
 ковалентная модификация гистонов специфическими ферментами: гистон ацетилтрансферазами (HAT), деацетилазами, метилтрансферазами,") Ремоделирование хроматина • 1) ковалентная модификация гистонов специфическими ферментами: гистон ацетилтрансферазами (HAT), деацетилазами, метилтрансферазами, киназами и 2) действие АТФ-зависимых комплексов, которые сдвигают, удаляют или реструктурируют нуклеосомы
Ремоделирование хроматина • 1) ковалентная модификация гистонов специфическими ферментами: гистон ацетилтрансферазами (HAT), деацетилазами, метилтрансферазами, киназами и 2) действие АТФ-зависимых комплексов, которые сдвигают, удаляют или реструктурируют нуклеосомы
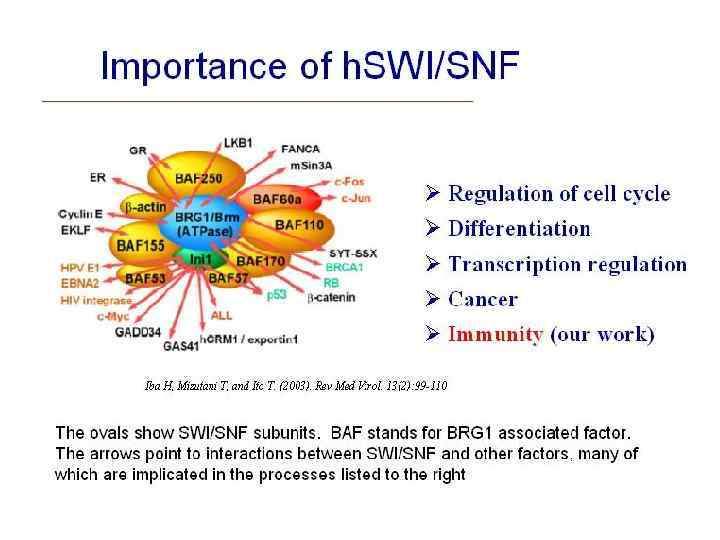
 – комплекс ремоделирования нуклеосом • Содержит продукты SWI") • SWI/SNF (SWItch/Sucrose Non. Fermentable) – комплекс ремоделирования нуклеосом • Содержит продукты SWI и SNF генов (SWI 1, SWI 2 / SNF 2, SWI 3, SWI 5, SWI 6), а также другие полипептиды. Он обладает АТФазной активностью и может дестабилизировать взаимодействие гистонов-ДНК в реконструированных нуклеосомах.
• SWI/SNF (SWItch/Sucrose Non. Fermentable) – комплекс ремоделирования нуклеосом • Содержит продукты SWI и SNF генов (SWI 1, SWI 2 / SNF 2, SWI 3, SWI 5, SWI 6), а также другие полипептиды. Он обладает АТФазной активностью и может дестабилизировать взаимодействие гистонов-ДНК в реконструированных нуклеосомах.
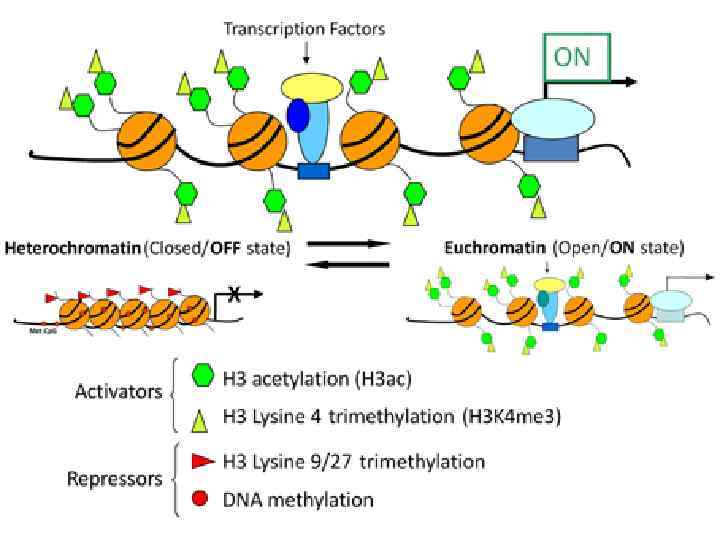
 Верхний рисунок: «включенный ген» . Хроматин находится в «открытой» конформации – ДНК не метилирована, гистоны ацетилированы. Сокращения – HAT – histone acetylase (ацетилаза гистонов). Нижний рисунок: «выключенный ген» . Хроматин находится в «закрытой» конформации – ДНК метилирована, гистоны деацетилированы. Сокращения – HDAC – histone deacetylase (деацетилаза гистонов), HMT – histone methyltransferase – гистоновая метилтрансфераза. Метилирование остатков лизина и аргинина в гистонах в различных положениях может инактивировать, либо активировать транскрипцию. Метилирование лизина 9 гистона Н 3 снижает транскрипцию.
Верхний рисунок: «включенный ген» . Хроматин находится в «открытой» конформации – ДНК не метилирована, гистоны ацетилированы. Сокращения – HAT – histone acetylase (ацетилаза гистонов). Нижний рисунок: «выключенный ген» . Хроматин находится в «закрытой» конформации – ДНК метилирована, гистоны деацетилированы. Сокращения – HDAC – histone deacetylase (деацетилаза гистонов), HMT – histone methyltransferase – гистоновая метилтрансфераза. Метилирование остатков лизина и аргинина в гистонах в различных положениях может инактивировать, либо активировать транскрипцию. Метилирование лизина 9 гистона Н 3 снижает транскрипцию.
. Sir - Silent Information Regulator") Функции Sir белков в Sirопосредованном глушении генов (Sir-mediated silencing). Sir - Silent Information Regulator – семейство генов, участвующих в деацетилировании гистонов и последующей конденсации хроматина. (A) Связывающиеся с сайленсером белки (ORC), а также Rap 1 (R) и Abf 1 (A) привлекают (recruit) Sir белки (1 - 4) к сайленсеру. Sir 2 деацетилирует ближайшие нуклеосомы, создавая сайты для прикрепления белков Sir 3 и Sir 4. Повторение цикла приводит к раширению зоны деацетилирования. (B) Показаны консервативные домены белков и названия их паралогов.
Функции Sir белков в Sirопосредованном глушении генов (Sir-mediated silencing). Sir - Silent Information Regulator – семейство генов, участвующих в деацетилировании гистонов и последующей конденсации хроматина. (A) Связывающиеся с сайленсером белки (ORC), а также Rap 1 (R) и Abf 1 (A) привлекают (recruit) Sir белки (1 - 4) к сайленсеру. Sir 2 деацетилирует ближайшие нуклеосомы, создавая сайты для прикрепления белков Sir 3 и Sir 4. Повторение цикла приводит к раширению зоны деацетилирования. (B) Показаны консервативные домены белков и названия их паралогов.
 linc. RNA, long intergenic noncoding RNA; sn. RNA, small nuclear RNA; sno. RNA, small nucleolar RNA; and misc. RNA, miscellaneous RNA.
linc. RNA, long intergenic noncoding RNA; sn. RNA, small nuclear RNA; sno. RNA, small nucleolar RNA; and misc. RNA, miscellaneous RNA.
 могут репрессировать или активировать") • длинные некодирующие РНК (long noncoding RNAs, lnc. RNAs) могут репрессировать или активировать гены, действуя как «наводчики» регуляторных белков (вариант ііі) или входя в состав комплекса белков хромосомного матрикса (вариант ii). POL II - РНК полимераза II. Monya Baker Long noncoding RNAs: the search for function Nature methods | VOL. 8 NO. 5 | MAY 2011 | 379
• длинные некодирующие РНК (long noncoding RNAs, lnc. RNAs) могут репрессировать или активировать гены, действуя как «наводчики» регуляторных белков (вариант ііі) или входя в состав комплекса белков хромосомного матрикса (вариант ii). POL II - РНК полимераза II. Monya Baker Long noncoding RNAs: the search for function Nature methods | VOL. 8 NO. 5 | MAY 2011 | 379
 могут репрессировать или активировать") • длинные некодирующие РНК (long noncoding RNAs, lnc. RNAs) могут репрессировать или активировать гены, действуя как «наводчики» регуляторных белков (вариант ііі) или входя в состав комплекса белков хромосомного матрикса (вариант ii). POL II - РНК полимераза II. Monya Baker Long noncoding RNAs: the search for function Nature methods | VOL. 8 NO. 5 | MAY 2011 | 379
• длинные некодирующие РНК (long noncoding RNAs, lnc. RNAs) могут репрессировать или активировать гены, действуя как «наводчики» регуляторных белков (вариант ііі) или входя в состав комплекса белков хромосомного матрикса (вариант ii). POL II - РНК полимераза II. Monya Baker Long noncoding RNAs: the search for function Nature methods | VOL. 8 NO. 5 | MAY 2011 | 379
 человеческий ген, локализованный в хромосоме 12.") • HOTAIR (for HOX transcript antisense RNA) человеческий ген, локализованный в хромосоме 12. Это первый пример РНК, экспрессируемой геном одной хромосомы, который влияет на транскрипцию в другой хромосоме. • Ген HOTAIR содержит 6, 232 п. о. и кодирует 2. 2 kb молекулу некодирующей РНК, которая контролирует экспрессию генов. Матрица дл этой РНК находится в генном кластере HOXC. Некодирующая РНК переносится из хромосомы 12 в хромосому It is shuttled from chromosome 12 to chromosome 2 с помощью белка Suz-Twelve. • 5' конец HOTAIR взаимодействует Polycomb-group белком protein Polycomb Repressive Complex 2 (PRC 2) и регулирует состояние хроматина. Происходит глушение локуса HOXD комплексом PRC 2. 3' крнец HOTAIR взаимодействует с деметилазой гистонов LSD 1. • HOTAIR особенно экспрессирован в клетках метастазирующего рака груди и клетках карцинома пищевода.
• HOTAIR (for HOX transcript antisense RNA) человеческий ген, локализованный в хромосоме 12. Это первый пример РНК, экспрессируемой геном одной хромосомы, который влияет на транскрипцию в другой хромосоме. • Ген HOTAIR содержит 6, 232 п. о. и кодирует 2. 2 kb молекулу некодирующей РНК, которая контролирует экспрессию генов. Матрица дл этой РНК находится в генном кластере HOXC. Некодирующая РНК переносится из хромосомы 12 в хромосому It is shuttled from chromosome 12 to chromosome 2 с помощью белка Suz-Twelve. • 5' конец HOTAIR взаимодействует Polycomb-group белком protein Polycomb Repressive Complex 2 (PRC 2) и регулирует состояние хроматина. Происходит глушение локуса HOXD комплексом PRC 2. 3' крнец HOTAIR взаимодействует с деметилазой гистонов LSD 1. • HOTAIR особенно экспрессирован в клетках метастазирующего рака груди и клетках карцинома пищевода.
 RNA HOTAIR узнает специфические последовательности ДНК и направляет хроматинмодифицирующие") Длинная межгенная некодирующая РНК (linc) RNA HOTAIR узнает специфические последовательности ДНК и направляет хроматинмодифицирующие комплексы PRC 2 и LSD 1 дя глушения этих комплексов. 5’ конец HOTAIR прикрепляется к PRC 2 с помощью non-coding RNA binding domain (nc. RBD) гистоновой метилазы HMTase EZH 2, катализирующей триметилирование H 3 K 27. 3’ конец HOTAIR облегчает деметилирование H 3 K 4 me 2 с помощью лизиновой деметилазы LSD 1. H 3 K 27 me 3 и недостаток метилирования H 3 K 4 репрессируют хроматин. Deborah J. Marsh*, Jaynish S. Shah and Alexander J. Cole Histones and their modifications in ovarian cancer – drivers of disease and therapeutic targets. Frontiers in Oncology 2014 | Volume 4 | Article 144
Длинная межгенная некодирующая РНК (linc) RNA HOTAIR узнает специфические последовательности ДНК и направляет хроматинмодифицирующие комплексы PRC 2 и LSD 1 дя глушения этих комплексов. 5’ конец HOTAIR прикрепляется к PRC 2 с помощью non-coding RNA binding domain (nc. RBD) гистоновой метилазы HMTase EZH 2, катализирующей триметилирование H 3 K 27. 3’ конец HOTAIR облегчает деметилирование H 3 K 4 me 2 с помощью лизиновой деметилазы LSD 1. H 3 K 27 me 3 и недостаток метилирования H 3 K 4 репрессируют хроматин. Deborah J. Marsh*, Jaynish S. Shah and Alexander J. Cole Histones and their modifications in ovarian cancer – drivers of disease and therapeutic targets. Frontiers in Oncology 2014 | Volume 4 | Article 144
 некодирующая РНК, которая имеет размеры 17 Mb у") • Xist (X-inactive specific transcript) некодирующая РНК, которая имеет размеры 17 Mb у человека. Ген Xist локализован в q плече Х-хромосомы (Xq 13) в так называмом X inactivation center – XIC - 73. 82 – 73. 85 Mb). • Инактивированная Х-хромосома покрыта молекулами Xist-РНК, которые необходимы для процесса инактивации хромосомы.
• Xist (X-inactive specific transcript) некодирующая РНК, которая имеет размеры 17 Mb у человека. Ген Xist локализован в q плече Х-хромосомы (Xq 13) в так называмом X inactivation center – XIC - 73. 82 – 73. 85 Mb). • Инактивированная Х-хромосома покрыта молекулами Xist-РНК, которые необходимы для процесса инактивации хромосомы.
 Xist RNA encompasses the X from which it is transcribed. RNA-fluorescence in situ hybridization detecting Xist RNA (red) localized on the inactive X in a preparation of condensed chromosomes from differentiated mouse cells. DNA is counterstained (blue). • Karen Ng, Dieter Pullirsch, Martin Leeb, Anton Wutz. Xist and the order of silencing. EMBO Rep. 2007 Jan; 8(1): 34– 39.
Xist RNA encompasses the X from which it is transcribed. RNA-fluorescence in situ hybridization detecting Xist RNA (red) localized on the inactive X in a preparation of condensed chromosomes from differentiated mouse cells. DNA is counterstained (blue). • Karen Ng, Dieter Pullirsch, Martin Leeb, Anton Wutz. Xist and the order of silencing. EMBO Rep. 2007 Jan; 8(1): 34– 39.
 • The X-inactivation centre regulates Xist expression to ensure that one X chromosome remains active. (A) Map of the regulatory elements implicated in counting and choice in the mouse Xic locus. The Xist gene, the antisense Tsix RNA, Xite, CCCTC-binding factor (CTCF)-binding sites at DXPas 34 and the region implicated for Xic pairing are indicated. (B) Proposed scheme of counting and choice that involves pairing of the Xic loci at the initiation of X-inactivation. Homologous X-chromosomes within one nucleus are shown and the Xic is highlighted in red. Pairing of the Xic loci could activate Xist transcription on one chromosome, therefore enforcing the commitment to become silenced, while the other chromosome remains active. Xa, active X; Xi, inactive X; Xic, X-inactivation centre.
• The X-inactivation centre regulates Xist expression to ensure that one X chromosome remains active. (A) Map of the regulatory elements implicated in counting and choice in the mouse Xic locus. The Xist gene, the antisense Tsix RNA, Xite, CCCTC-binding factor (CTCF)-binding sites at DXPas 34 and the region implicated for Xic pairing are indicated. (B) Proposed scheme of counting and choice that involves pairing of the Xic loci at the initiation of X-inactivation. Homologous X-chromosomes within one nucleus are shown and the Xic is highlighted in red. Pairing of the Xic loci could activate Xist transcription on one chromosome, therefore enforcing the commitment to become silenced, while the other chromosome remains active. Xa, active X; Xi, inactive X; Xic, X-inactivation centre.
") • Models for Xist spreading along the chromosome in cis. Xist RNA (red) and protein factors postulated to bind Xist (orange) are shown. (A) Xist spreading along the chromosome by means of ‘way stations' or ‘boosters'. (B) Xist spreading based on a cooperative binding mechanism. Xist spreads in cis from its transcription site, where the high RNA concentration is predicted to nucleate chromatin attachment. (C) Schematic representation of the Xi territory with Xist spreading inward from the Xic to the centre of the chromosome territory. LINEs, long interspersed elements; Xic, Xinactivation centre.
• Models for Xist spreading along the chromosome in cis. Xist RNA (red) and protein factors postulated to bind Xist (orange) are shown. (A) Xist spreading along the chromosome by means of ‘way stations' or ‘boosters'. (B) Xist spreading based on a cooperative binding mechanism. Xist spreads in cis from its transcription site, where the high RNA concentration is predicted to nucleate chromatin attachment. (C) Schematic representation of the Xi territory with Xist spreading inward from the Xic to the centre of the chromosome territory. LINEs, long interspersed elements; Xic, Xinactivation centre.
 Гипотетическая модель участия длинных некодирующих РНК в регуляции транскрипции, как матрицы, на которой собираются комплексы белков, регулирующих транскрипцию
Гипотетическая модель участия длинных некодирующих РНК в регуляции транскрипции, как матрицы, на которой собираются комплексы белков, регулирующих транскрипцию
 Small interfering RNA (si. RNA)") Возможные функции некодирующих РНК (нк. РНК, nc. RNA) Small interfering RNA (si. RNA)
Возможные функции некодирующих РНК (нк. РНК, nc. RNA) Small interfering RNA (si. RNA)
 Схема образования малых ядерных РНК путем разрезания нк РНК
Схема образования малых ядерных РНК путем разрезания нк РНК
 РНКаза Dicer-2 разрезает двухцепочечную ds. RNA (РНК вирусов, например у человека встречаются вирусы рода Rotavirus семейства Reoviridae, которые могут вызывать диарею) на фрагменты малых интерферирующих РНК (small interfering RNA, si. RNA), приблизительно по 20 -25 нуклеотидов, которые попадают в ферментный комплекс RISC (RNA-induced silencing complex) и подвергают деградации РНК вируса с помощью белка Argonaute-2 Argonaute, является эндонуклеазой, деградирующей м. РНК, комплементарные ведущей цепи si. RNA.
РНКаза Dicer-2 разрезает двухцепочечную ds. RNA (РНК вирусов, например у человека встречаются вирусы рода Rotavirus семейства Reoviridae, которые могут вызывать диарею) на фрагменты малых интерферирующих РНК (small interfering RNA, si. RNA), приблизительно по 20 -25 нуклеотидов, которые попадают в ферментный комплекс RISC (RNA-induced silencing complex) и подвергают деградации РНК вируса с помощью белка Argonaute-2 Argonaute, является эндонуклеазой, деградирующей м. РНК, комплементарные ведущей цепи si. RNA.
 Предшественники mi. RNA транскрибируются и подвергаются") Регуляция трансляции с помощью микро РНК (mi. RNA) Предшественники mi. RNA транскрибируются и подвергаются процессингу РНКазами Drosha и Dicer-1 до зрелой mi. RNA. Последняя загружается в комплекс RISC, содержащий фермент Argonaute-1, который присоединяется к м. РНК и привлекает ряд других белков – ингибиторов трансляции
Регуляция трансляции с помощью микро РНК (mi. RNA) Предшественники mi. RNA транскрибируются и подвергаются процессингу РНКазами Drosha и Dicer-1 до зрелой mi. RNA. Последняя загружается в комплекс RISC, содержащий фермент Argonaute-1, который присоединяется к м. РНК и привлекает ряд других белков – ингибиторов трансляции
 РНКазы Aubergine") Участие РНКи в подавлении экспрессии подвижных генетических элементов у дрозофилы (модель пинг-понга) РНКазы Aubergine и Argonaute-3 разрезают значащую (отмечено красным) и незначащую (отмечено голубым) цепи в транскриптах с мобильных элементов (ТЕ), причем для наведения на цель служат pi. RNA – особый класс РНКи. Вырезание инактивирует транскрипт и приводит к образованию новой pi. RNA, этот предшественник pi. RNA связывается белком из семейства Piwi, при его участии образуется 30 конец молекулы, который модифицируется метилированием (Me). Активный Piwi комплекс связывает транскрипты с мобильных элементов и белковые факторы, вызывающие образование гетерохроматина.
Участие РНКи в подавлении экспрессии подвижных генетических элементов у дрозофилы (модель пинг-понга) РНКазы Aubergine и Argonaute-3 разрезают значащую (отмечено красным) и незначащую (отмечено голубым) цепи в транскриптах с мобильных элементов (ТЕ), причем для наведения на цель служат pi. RNA – особый класс РНКи. Вырезание инактивирует транскрипт и приводит к образованию новой pi. RNA, этот предшественник pi. RNA связывается белком из семейства Piwi, при его участии образуется 30 конец молекулы, который модифицируется метилированием (Me). Активный Piwi комплекс связывает транскрипты с мобильных элементов и белковые факторы, вызывающие образование гетерохроматина.
 у дрозофилы— семейство генов, кодирующих регуляторные") Piwi (PIWI, от англ. Pelement induced wimpy testis) у дрозофилы— семейство генов, кодирующих регуляторные белки, Некоторые белки PIWI связывают с особые малые некодирующие РНК — pi. РНК, и функционируют в комплексе с ними. Обычно их активность направлена на сайленсинг транспозонов в клетках зародышевой линии.
Piwi (PIWI, от англ. Pelement induced wimpy testis) у дрозофилы— семейство генов, кодирующих регуляторные белки, Некоторые белки PIWI связывают с особые малые некодирующие РНК — pi. РНК, и функционируют в комплексе с ними. Обычно их активность направлена на сайленсинг транспозонов в клетках зародышевой линии.
 или small interfering RNA (si. RNA) как средство") Малые ингибиторные РНК (20 -25 нуклеотидов) или small interfering RNA (si. RNA) как средство для избирательного ингибирования экспрессии генов
Малые ингибиторные РНК (20 -25 нуклеотидов) или small interfering RNA (si. RNA) как средство для избирательного ингибирования экспрессии генов
 • Транскрипцио нный фактор NF-κB (ядерный фактор «каппа -би» ; англ. nuclear factor kappa-light-chain-enhancer of activated B cells, NF-k. B) — универсальный фактор транскрипции, контролирующий экспрессию генов иммунного ответа, апоптоза и клеточного цикла. Нарушение регуляции NF-k. B вызывает воспаление, аутоиммунные заболевания, а также развитие вирусных инфекций и рака. Семейство NF-k. B состоит из 5 белков: NF-k. B 1 (или p 50), NF-k. B 2 (или p 52), Rel. A (или p 65), Rel. B и c-Rel, образующих 15 комбинаций димеров. Все белки семейства объединяет наличие домена гомологии Rel, который обеспечивает образование белковых димеров, связывание NF-k. B с ДНК и с цитозольным ингибиторным белком Ik. B. Фактор NF-k. B проявляет активность только в димерной форме (возможно образование как гетеро-, так и гомодимеров), причём наиболее распространённые формы — димеры субъединиц p 50 или p 52 с субъединицей p 65. • NF-k. B активируется целым рядом стимулов, включая цитокины (такие как TNF и интерлейкин 1), T- и Bклеточные митогены, бактериальные и вирусные продукты
• Транскрипцио нный фактор NF-κB (ядерный фактор «каппа -би» ; англ. nuclear factor kappa-light-chain-enhancer of activated B cells, NF-k. B) — универсальный фактор транскрипции, контролирующий экспрессию генов иммунного ответа, апоптоза и клеточного цикла. Нарушение регуляции NF-k. B вызывает воспаление, аутоиммунные заболевания, а также развитие вирусных инфекций и рака. Семейство NF-k. B состоит из 5 белков: NF-k. B 1 (или p 50), NF-k. B 2 (или p 52), Rel. A (или p 65), Rel. B и c-Rel, образующих 15 комбинаций димеров. Все белки семейства объединяет наличие домена гомологии Rel, который обеспечивает образование белковых димеров, связывание NF-k. B с ДНК и с цитозольным ингибиторным белком Ik. B. Фактор NF-k. B проявляет активность только в димерной форме (возможно образование как гетеро-, так и гомодимеров), причём наиболее распространённые формы — димеры субъединиц p 50 или p 52 с субъединицей p 65. • NF-k. B активируется целым рядом стимулов, включая цитокины (такие как TNF и интерлейкин 1), T- и Bклеточные митогены, бактериальные и вирусные продукты
 Under resting conditions, NF-κB dimers are bound to inhibitory IκB proteins, which sequester inactive NFκB complexes in the cytoplasm. Stimulus-induced degradation of IκB proteins is initiated through phosphorylation by the IκB kinase Лимфотоксин-бета (англ. Lymphotoxin-beta) — мембранный цитокин, лимфокин из суперсемейства факторов некроза опухоли. Вместе с лимфотоксином-альфа играет роль в нормальном иммунном гомеостазе.
Under resting conditions, NF-κB dimers are bound to inhibitory IκB proteins, which sequester inactive NFκB complexes in the cytoplasm. Stimulus-induced degradation of IκB proteins is initiated through phosphorylation by the IκB kinase Лимфотоксин-бета (англ. Lymphotoxin-beta) — мембранный цитокин, лимфокин из суперсемейства факторов некроза опухоли. Вместе с лимфотоксином-альфа играет роль в нормальном иммунном гомеостазе.
 Схема строения транскрипционной фабрики Диаметр фабрики – 56, 75 нм, по данным разных авторов В клетках He. La приблизительно 8000 фабрик RNAPII и 2000 RNAPIII
Схема строения транскрипционной фабрики Диаметр фабрики – 56, 75 нм, по данным разных авторов В клетках He. La приблизительно 8000 фабрик RNAPII и 2000 RNAPIII
 Две возможных схемы сплайсинга генов, транскрибируемых на двух транскрипционных фабриках с – транскрипция двух генов идет на общей транскрипционной фабрике в - транскрипция двух генов идет на разных транскрипционных фабриках, объединение в одну м. РНК идет за счет факторов сплайсинга (желтые кружочки)
Две возможных схемы сплайсинга генов, транскрибируемых на двух транскрипционных фабриках с – транскрипция двух генов идет на общей транскрипционной фабрике в - транскрипция двух генов идет на разных транскрипционных фабриках, объединение в одну м. РНК идет за счет факторов сплайсинга (желтые кружочки)
 Транскрипционные фабрики в клетках He. La Транскрипционные фабрики указаны стрелочками, интерхроматиновые гранулы - звездочками
Транскрипционные фабрики в клетках He. La Транскрипционные фабрики указаны стрелочками, интерхроматиновые гранулы - звездочками
 Транскрипция двух генов в эритробластах человека SC 35 – фактор сплайсинга
Транскрипция двух генов в эритробластах человека SC 35 – фактор сплайсинга
 Транскрипция глобиновых генов в эритробластах человека Красный цвет – РНК-пол II Зеленый –РНК Два глобиновых гена транскрибируются на разных полимеразных фабриках
Транскрипция глобиновых генов в эритробластах человека Красный цвет – РНК-пол II Зеленый –РНК Два глобиновых гена транскрибируются на разных полимеразных фабриках
 в клетках He. La Гранулы содержат мя. РНК") Интерхроматиновые гранулы (speckles, 20 -25 нм) в клетках He. La Гранулы содержат мя. РНК и факторы сплайсинга Размер черточки – 5 мкм
Интерхроматиновые гранулы (speckles, 20 -25 нм) в клетках He. La Гранулы содержат мя. РНК и факторы сплайсинга Размер черточки – 5 мкм
 окрасили") Роль РНК MEN b, в образовании гранул paraspeckles в ядре paraspeckles (желтые точки) окрасили антителами к белку PSPC 1, характерному для них MEN b, >20 -kb nc. RNA Содержится в ядре в гранулах paraspeckles Нижний ряд фотографий – клетки He. La обработали антисенсолигонуклеотидами к MEN b, в результате чего paraspeckles исчезли
Роль РНК MEN b, в образовании гранул paraspeckles в ядре paraspeckles (желтые точки) окрасили антителами к белку PSPC 1, характерному для них MEN b, >20 -kb nc. RNA Содержится в ядре в гранулах paraspeckles Нижний ряд фотографий – клетки He. La обработали антисенсолигонуклеотидами к MEN b, в результате чего paraspeckles исчезли
 Схема изменений в состоянии фосфорилирования большой субъединицы РНК-полимеразы -II процессе транскрипции Большая субъединица ( RPB 1) полимеразы RNAP II имеет C-терминальную область (CTD), состоящую из 52 тандемных серинбогатых повторов, которые могут быть посттрансяционно фосфорилированы. Изначально RNAP II нефосфорилирована, затем, в процессе инициации Ser 5 фосфорилируется с помощью фактора транскрипции TFIIH. В процессе элонгации наблюдается Фосфорилирование серина Ser 2 киназой CTDKI, которая является частью фактора элонгации PЌTEFb. На завершающем этапе элонгации происходит дефосфорилирование по Ser 5 с помощью протеинфосфатазы. Фосфорилирование Ser 7 необходимо для транскрипции малых ядерных РНК.
Схема изменений в состоянии фосфорилирования большой субъединицы РНК-полимеразы -II процессе транскрипции Большая субъединица ( RPB 1) полимеразы RNAP II имеет C-терминальную область (CTD), состоящую из 52 тандемных серинбогатых повторов, которые могут быть посттрансяционно фосфорилированы. Изначально RNAP II нефосфорилирована, затем, в процессе инициации Ser 5 фосфорилируется с помощью фактора транскрипции TFIIH. В процессе элонгации наблюдается Фосфорилирование серина Ser 2 киназой CTDKI, которая является частью фактора элонгации PЌTEFb. На завершающем этапе элонгации происходит дефосфорилирование по Ser 5 с помощью протеинфосфатазы. Фосфорилирование Ser 7 необходимо для транскрипции малых ядерных РНК.