


Синтез белка
; - СН 3 (аланин) - СН 2")
Белки Аминокислоты R R - H (глицин); - СН 3 (аланин) - СН 2 – SH (цистеин) + Пептидная связь

Механизм, с помощью которого последовательность аминокислот в первичной структуре белка определяется последовательностью нуклеотидов в цепи нуклеиновых кислот, называется генетическим кодом. Комбинация из трех нуклеотидов, соответствующая каждой аминокислоте, составляет триплет или кодон.
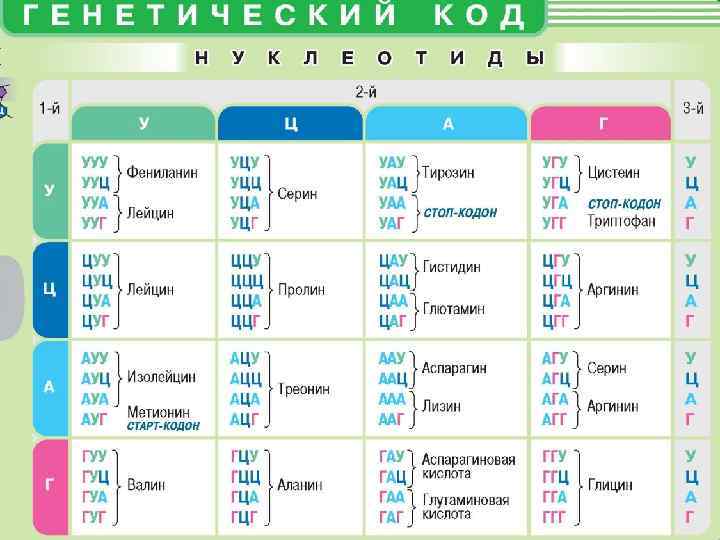
 универсален для всех организмов не перекрывается")
Свойства кода - специфичность (соответствует только одной аминокислоте) универсален для всех организмов не перекрывается является избыточным Участок ДНК, несущий информацию о структуре одной белковой молекулы, называется геном. В начале гена располагается участок ДНК, регулирующий его активность. За ним следуют промотор, инициирующий транскрипцию, и смысловая часть гена, кодирующая последовательность аминокислот в белках. У большинства прокариот белок-кодирующие последовательности (экзоны) следуют непрерывно, но у эукариотических организмов они прерываются некодирующими участками ДНК (интронами). Наследственная информация, записанная с помощью генетического кода, хранится в молекулах ДНК. Перевод этой информации в функциональную форму осуществляется с помощью РНК.
, фосфата и одного из четырех азотистых")
РНК Нуклеотиды РНК состоят из рибозы (вместо дезоксирибозы), фосфата и одного из четырех азотистых оснований - аденина, гуанина, цитозина и урацила (вместо тимина). РНК синтезируется на молекулах ДНК при помощи фермента РНК-полимеразы с соблюдением принципа комплементарности и антипараллельности, причем аденину ДНК в РНК комплементарен урацил. Из всего многообразия РНК, действующих в клетке, можно выделить три основных вида: м. РНК, т. РНК, р. РНК. А. NA

м. РНК Информация о порядке включения аминокислот в пептидную цепь заключена в нуклеотидной последовательности матричных, или информационных РНК (м. РНК, и. РНК). Процесс синтеза м. РНК называют транскрипцией. Транскрипция Синтез м. РНК начинается с обнаружения РНКполимеразой особого участка в молекуле ДНК, который указывает место начала транскрипции — промотора. После присоединения к промотору РНК-полимераза раскручивает прилежащий виток спирали ДНК. Две цепи ДНК в этом месте расходятся, и на одной из них фермент осуществляет синтез м. РНК. Сборка аминокислот в цепь происходит с соблюдением правила комплементарности между нуклеотидами м. РНК и нуклеотидам ДНК антипараллельно по отношению к матричной цепи ДНК. Таким образом матрицей для транскрипции может служить только одна из двух цепей ДНК, а именно та, которая обращена к ферменту своим 3'-концом. Такую цепь называют кодогенной.

Продвигаясь вдоль кодогенной цепи ДНК, РНК-полимераза осуществляет точное переписывание информации до тех пор, пока она не встречает терминатор транскрипции. В этом участке РНКполимераза отделяется как от матрицы ДНК, так и от вновь синтезированной м. РНК. В процессе синтеза, по мере продвижения РНК-полимеразы вдоль молекулы ДНК, пройденные ею одноцепочечные участки ДНК вновь объединяются в двойную спираль. Образуемая в ходе транскрипции м. РНК содержит точную копию информации, записанной в соответствующем участке ДНК. М. 8. 7
 РНК. Формирование зрелых молекул РНК называется")
Первичным продуктом транскрипции служит так называемая гетерогенная (ядерная) РНК. Формирование зрелых молекул РНК называется процессингом. В ходе созревания молекул РНК они подвергаются сплайсингу - удалению нуклеотидных последовательностей, соответствующих интронам ДНК, и единению между собой транскрибированных участков экзонов. М. эн.
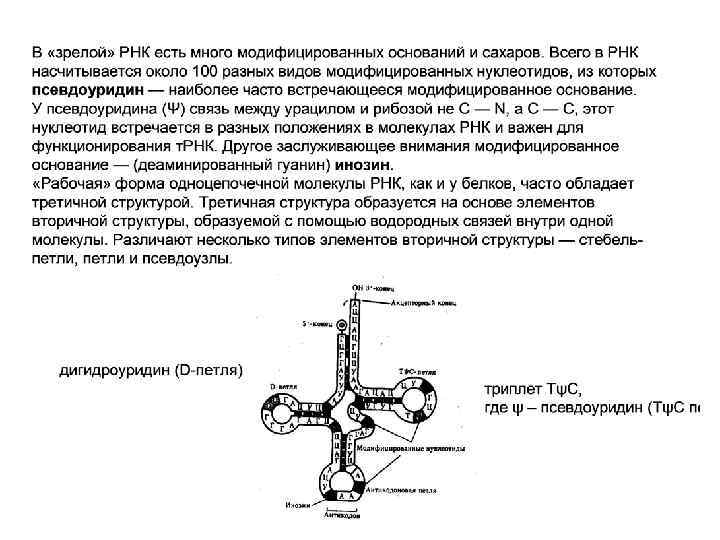

Инозин – способен соединяться с тремя разными основаниями кодона м. РНК – У, Ц и А. Благодаря этой особенности одна т. РНК узнает несколько кодонов-синонимов. В результате в клетке встречается не 61 (по количеству кодонов), а около 40 различных молекул т. РНК. дигидроуридин (D-петля) триплет TψC, где ψ – псевдоуриаин (TψC петля).

т. РНК, трансляция. Доставляя необходимые аминокислоты к месту сборки пептидных цепей, т. РНК выполняет функцию трансляции (передачи). Для т. РНК характерно образование двух перпендикулярно расположенных цепей. На конце одной из цепей располагается транспортируемая аминокислота (акцепторная ветвь), на конце другой - антикодон (антикодоновая ветвь). Эти участки максимально удалены друг от друга и стабильны за счет взаимодействия в третичной структуре двух петель, содержащих модифицированные основания – дигидроуридин (D-петля) и триплета TψC, где ψ – псевдоуридин (TψC петля).
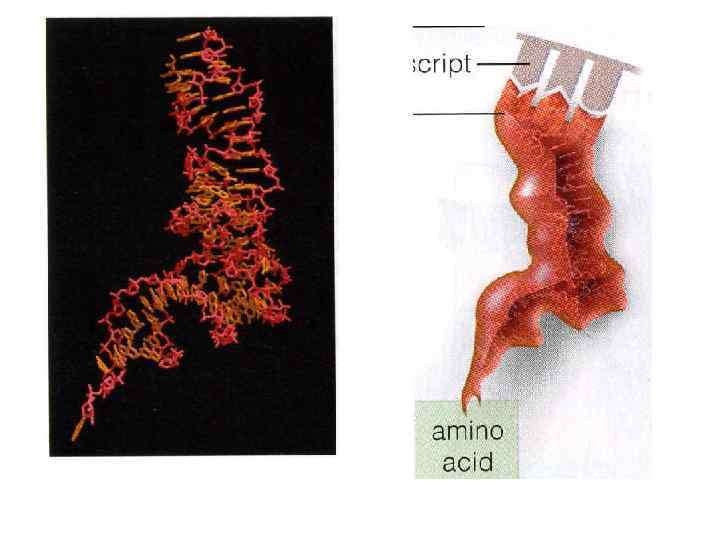

Наследственная информация, «записанная» в молекулах ДНК и «переписанная» на м. РНК, расшифровывается в ходе трансляции благодаря двум процессам специфического узнавания молекулярных поверхностей. Сначала фермент аминоацил-т. РНК-синтетаза обеспечивает соединение т. РНК с аминокислотой. Затем аминоацил-т. РНК комплементарно спаривается с м. РНК благодаря взаимодействию антикодона с кодоном.
. Процесс взаимодействия м. РНК и т. РНК осуществляется на рибосомах. В")
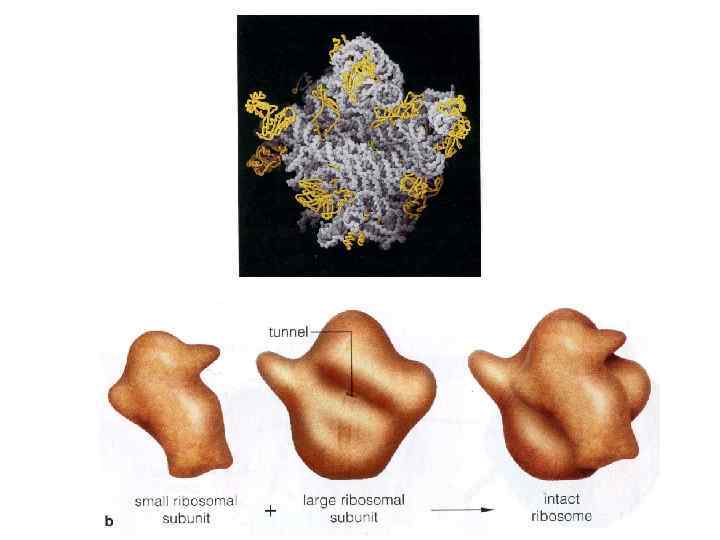
Рибосомальная РНК(р. РНК). Процесс взаимодействия м. РНК и т. РНК осуществляется на рибосомах. В рибосомах выделяют два участка. В аминоацильном (А-участке) размещается аминоацил-т. РНК, несущая определенную аминокислоту. В пептидильном (П-участке) располагается обычно т. РНК, которая нагружена цепочкой аминокислот, соединенных пептидными связями. Образование А- и П-участков обеспечивается двумя субчастицами рибосомы.

Фаза инициации. В молекуле любой м. РНК вблизи ее 5'-конца имеется участок, комплементарный р. РНК малой субчастицы рибосомы и специфически узнаваемый ею. Рядом с ним располагается инициирующий стартовый кодон АУГ, шифрующий аминокислоту метионин. Малая субчастица рибосомы соединяется с м. РНК таким образом, что стартовый кодон располагается в области, соответствующей П-участку. Затем происходит объединение большой и малой субчастиц рибосомы с образованием ее пептидильного и аминоацильного участков. Процесс инициации катализируется особыми белками — факторами инициации, которые связаны с малой субчастицей рибосомы. По завершении фазы инициации и образования комплекса рибосома — м. РНК — инициирующая аминоацил-т. РНК, эти факторы отделяются от рибосомы.

Фаза элонгации. Благодаря особенностям трехмерной организации т. РНК при соединении ее антикодона с кодоном м. РНК, транспортируемая ею аминокислота, располагается в А-участке, поблизости от ранее включенной аминокислоты, находящейся в Пучастке. Между двумя аминокислотами образуется пептидная связь, катализируемая особыми белками, входящими в состав рибосомы. В результате предыдущая аминокислота теряет связь со своей т. РНК и присоединяется к аминоацил -т. РНК, расположенной в А-участке. Находящаяся в этот момент в Пучастке т. РНК высвобождается и уходит в цитоплазму. Перемещение т. РНК, нагруженной пептидной цепочкой, из А-участка в П-участок сопровождается продвижением рибосомы по м. РНК на шаг, соответствующий одному кодону.

Фаза терминации связана с узнаванием специфическим рибосомным белком одного из завершающих кодонов (УАА, УАГ или УГА), когда тот входит в зону Аучастка рибосомы. При этом к последней аминокислоте в пептидной цепи присоединяется вода, и ее карбоксильный конец отделяется от т. РНК. В результате завершенная пептидная цепь теряет связь с рибосомой, которая распадается на две субчастицы. M. Trl.


Продвигаясь вдоль кодогенной цепи ДНК, РНКполимераза осуществляет постепенное точное переписывание информации до тех пор, пока она не встречает специфическую нуклеотидную последовательность — терминатор транскрипции. В этом участке РНК-полимераза отделяется как от матрицы ДНК, так и от вновь синтезированной м. РНК. Фрагмент молекулы ДНК, включающий промотор, транскрибируемую последовательность и терминатор, образует единицу транскрипции — транскриптон. В процессе синтеза, по мере продвижения РНКполимеразы вдоль молекулы ДНК, пройденные ею одноцепочечные участки ДНК вновь объединяются в двойную спираль. Образуемая в ходе транскрипции м. РНК содержит точную копию информации, записанной в соответствующем участке ДНК. Тройки рядом стоящих нуклеотидов м. РНК, шифрующие аминокислоты, называют кодонами.

т. РНК. Трансляция. Доставляя необходимые аминокислоты к месту сборки пептидных цепей, т. РНК выполняет функцию трансляции (передачи). В ней выделяют четыре главные части: Акцепторный конец - образуется двумя комплементарно соединенными частями т. РНК. К этому участку присоединяется транспортируемая аминокислота. Остальные три ветви представляют собой комплементарно спаренные последовательности нуклеотидов, которые заканчиваются неспаренными участками, образующими петли. Средняя из этих ветвей содержит в центре своей петли антикодон. Антикодон — это три нуклеотида, комплементарные кодону м. РНК, который шифрует аминокислоту, транспортируемую данной молекулой к месту синтеза пептида. Между акцепторной и антикодоновой ветвями располагаются две боковые ветви, содержащие модифицированные основания.

Первичная структура т. РНК, определяемая последовательностью нуклеотидов, формирует вторичную структуру т. РНК, имеющую форму листа клевера. В свою очередь, вторичная структура обусловливает трехмерную третичную структуру, для которой характерно образование двух перпендикулярно расположенных двойных спиралей. Одна из них образована акцепторной и ТψС -ветвями, другая антикодоновой и D -ветвями. На конце одной из двойных спиралей располагается транспортируемая аминокислота, на конце другой - антикодон. Эти участки максимально удалены друг от друга. Стабильность такой структуры т. РНК поддерживается благодаря возникновению дополнительных водородных связей между основаниями полинуклеотидной цепи, находящимися в разных ее участках, но пространственно сближенных в третичной структуре. Различные виды т. РНК имеют сходную третичную структуру, хотя и с некоторыми вариациями.

Наследственная информация, «записанная» в молекулах ДНК и «переписанная» на м. РНК, расшифровывается в ходе трансляции благодаря двум процессам специфического узнавания молекулярных поверхностей. Сначала фермент аминоацил-т. РНК-синтетаза обеспечивает соединение т. РНК с транспортируемой ею аминокислотой. Затем аминоацилт. РНК комплементарно спаривается с м. РНК благодаря взаимодействию антикодона с кодоном.

Фаза инициации. В молекуле любой м. РНК вблизи ее 5'-конца имеется участок, комплементарный р. РНК малой субчастицы рибосомы и специфически узнаваемый ею. Рядом с ним располагается инициирующий стартовый кодон АУГ, шифрующий аминокислоту метионин. Малая субчастица рибосомы соединяется с м. РНК таким образом, что стартовый кодон располагается в области, соответствующей П-участку. При этом только инициирующая т. РНК, несущая метионин, способна занять место в недостроенном П-участке малой субчастицы и комплементарно соединиться со стартовым кодоном. После описанного события происходит объединение большой и малой субчастиц рибосомы с образованием ее пептидильного и аминоацильного участков. Процесс инициации катализируются особыми белками — факторами инициации, которые связаны с малой субчастицей рибосомы. По завершении фазы инициации и образования комплекса рибосома — м. РНК — инициирующая аминоацил-т. РНК, эти факторы отделяются от рибосомы.