

Синапсы -1 (2008).ppt
- Количество слайдов: 168
 Синапсы Учение о синапсе.
Синапсы Учение о синапсе.
 История вопроса Термины «синапс» и «синаптическая передача» предложены в 1897 году Ч. Шеррингтоном для объяснения сообщения между нейронами в ЦНС
История вопроса Термины «синапс» и «синаптическая передача» предложены в 1897 году Ч. Шеррингтоном для объяснения сообщения между нейронами в ЦНС
 История вопроса В разработке учения о синапсах большую роль сыграли О. Леви, Г. Дейл, Д. Ленгли, Дж. Экклс, Дель Кастильо, В. Катц, А. Ф. Самойлов, А. В. Кибяков, Х. С. Коштоянц, Д. Г. Магазанник, Г. И. Полетаев, Е. Е. Никольский, А. Л. Зефиров
История вопроса В разработке учения о синапсах большую роль сыграли О. Леви, Г. Дейл, Д. Ленгли, Дж. Экклс, Дель Кастильо, В. Катц, А. Ф. Самойлов, А. В. Кибяков, Х. С. Коштоянц, Д. Г. Магазанник, Г. И. Полетаев, Е. Е. Никольский, А. Л. Зефиров
 Определение Синапс – это морфофункцнональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку).
Определение Синапс – это морфофункцнональное образование ЦНС, которое обеспечивает передачу сигнала с нейрона на другой нейрон или с нейрона на эффекторную клетку (мышечное волокно, секреторную клетку).
 Синапсы Общая физиология синаптической передачи.
Синапсы Общая физиология синаптической передачи.
 Синапсы Классификация синапсов центральной и периферической нервной системы.
Синапсы Классификация синапсов центральной и периферической нервной системы.
 Классификация синапсов Различают следующие виды синапсов ЦНС 1. По локализации – центральные (головной и спинной мозг) и периферические (нервно мышечный, нейросекреторный, синапсы вегетативной нервной системы). Виды центральных синапсов аксональные, аксо дендритические (дендритные), аксо соматические, дендро дендритические, дендро соматические и т. п.
Классификация синапсов Различают следующие виды синапсов ЦНС 1. По локализации – центральные (головной и спинной мозг) и периферические (нервно мышечный, нейросекреторный, синапсы вегетативной нервной системы). Виды центральных синапсов аксональные, аксо дендритические (дендритные), аксо соматические, дендро дендритические, дендро соматические и т. п.
 синапс.") Аксосоматический синапс. (аксосоматический) синапс.
Аксосоматический синапс. (аксосоматический) синапс.
 Подпись к рисунку из Орлова Р. С. Аксосоматический синапс А На поверхности перикариона заканчивается множество (до 10 000) пресинаптических терминалей аксонов от других нейронов. Это аксосоматические синапсы (1). Б Крупный масштаб аксосоматического синапса
Подпись к рисунку из Орлова Р. С. Аксосоматический синапс А На поверхности перикариона заканчивается множество (до 10 000) пресинаптических терминалей аксонов от других нейронов. Это аксосоматические синапсы (1). Б Крупный масштаб аксосоматического синапса
") Классификация синапсов 2. По развитию в онтогенезе – стабильные (например, синапсы дуг безусловного рефлекса) и динамичные (т. е. появляющиеся в процессе индивидуального развития) 3. По конечному эффекту – тормозные (тормозящие) и возбуждающие. 4. По механизму передачи сигнала – электрические, химические, смешанные.
Классификация синапсов 2. По развитию в онтогенезе – стабильные (например, синапсы дуг безусловного рефлекса) и динамичные (т. е. появляющиеся в процессе индивидуального развития) 3. По конечному эффекту – тормозные (тормозящие) и возбуждающие. 4. По механизму передачи сигнала – электрические, химические, смешанные.
 преходящие (варикозные") Классификация синапсов Виды химических синапсов по форме контакта – терминальные (колбообразные соединения) преходящие (варикозные расширения аксона);
Классификация синапсов Виды химических синапсов по форме контакта – терминальные (колбообразные соединения) преходящие (варикозные расширения аксона);
, 2.") Классификация синапсов Виды химических синапсов по природе медиатора 1 холинергические (медиатор – ацетилхолин), 2. адренергические (норадреналин, адреналин), 3. дофаминергические (дофамин), 4. серотонинергические (серотонин), 5. гистаминергические (гистамин)
Классификация синапсов Виды химических синапсов по природе медиатора 1 холинергические (медиатор – ацетилхолин), 2. адренергические (норадреналин, адреналин), 3. дофаминергические (дофамин), 4. серотонинергические (серотонин), 5. гистаминергические (гистамин)
, 7. ГАМК ергические (гамма аминомасляная кислота, или ГАМК), 8") Классификация синапсов 6. глутаматергические (глутамат), 7. ГАМК ергические (гамма аминомасляная кислота, или ГАМК), 8 бензодиазепинергические (бензодиазепин) 9. глицинергические (глицин), 10. пептидергические (пептиды, например, вещество Р) 11. пуринергические (АТФ, аденозин), 12. азотергические (оксид азота, или
Классификация синапсов 6. глутаматергические (глутамат), 7. ГАМК ергические (гамма аминомасляная кислота, или ГАМК), 8 бензодиазепинергические (бензодиазепин) 9. глицинергические (глицин), 10. пептидергические (пептиды, например, вещество Р) 11. пуринергические (АТФ, аденозин), 12. азотергические (оксид азота, или
.") Синапсы Электрические синапсы (эфапсы).
Синапсы Электрические синапсы (эфапсы).
. В ЦНС (в стволе мозга) имеются электрические синапсы (эфапсы) Эфапс –") Электрические синапсы (эфапсы). В ЦНС (в стволе мозга) имеются электрические синапсы (эфапсы) Эфапс – это щелевидное образование (размеры щели до 2 нм) с ионными мостиками каналами между двумя контактирующими клетками.
Электрические синапсы (эфапсы). В ЦНС (в стволе мозга) имеются электрические синапсы (эфапсы) Эфапс – это щелевидное образование (размеры щели до 2 нм) с ионными мостиками каналами между двумя контактирующими клетками.
. Петли тока, в частности при наличии потенциала действия (ПД), почти беспрепятственно") Электрические синапсы (эфапсы). Петли тока, в частности при наличии потенциала действия (ПД), почти беспрепятственно проходят через такой щелевидный контакт и возбуждают, т. е. индуцируют генерацию ПД в соседней клетке. эфапсы обеспечивают очень быструю передачу возбуждения
Электрические синапсы (эфапсы). Петли тока, в частности при наличии потенциала действия (ПД), почти беспрепятственно проходят через такой щелевидный контакт и возбуждают, т. е. индуцируют генерацию ПД в соседней клетке. эфапсы обеспечивают очень быструю передачу возбуждения
. Недостатки эфапсов С их помощью нельзя обеспечить одностороннее проведение, так как") Электрические синапсы (эфапсы). Недостатки эфапсов С их помощью нельзя обеспечить одностороннее проведение, так как большая часть таких синапсов обладает двусторонней проводимостью. С их помощью нельзя вызывать торможение эффекторной клетки.
Электрические синапсы (эфапсы). Недостатки эфапсов С их помощью нельзя обеспечить одностороннее проведение, так как большая часть таких синапсов обладает двусторонней проводимостью. С их помощью нельзя вызывать торможение эффекторной клетки.
. Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются") Электрические синапсы (эфапсы). Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются щелевые контакты типа нексуса.
Электрические синапсы (эфапсы). Аналогом электрического синапса в гладких мышцах и в сердечной мышце являются щелевые контакты типа нексуса.
 Химические синапсы
Химические синапсы
 Нервно мышечный синапс
Нервно мышечный синапс
 Химический синапс
Химический синапс
 Синапсы Основные вопросы: Общие представления о строении и механизмах функционирования химических синапсов – пресинаптическая часть, постсинаптическая часть, синаптическая щель. Медиаторы, сомедиаторы, комедиаторы, Принцип Дейла. Модуляторы синаптической передачи. Рецепторы ферменты
Синапсы Основные вопросы: Общие представления о строении и механизмах функционирования химических синапсов – пресинаптическая часть, постсинаптическая часть, синаптическая щель. Медиаторы, сомедиаторы, комедиаторы, Принцип Дейла. Модуляторы синаптической передачи. Рецепторы ферменты
") Общие представления о строении химических синапсов Химические синапсы представляют собой окончания аксона (терминальные синапсы) или его варикозную часть (проходящие синапсы). Он состоит из трех компонентов пресинаптической части постсинаптической части синаптической щели.
Общие представления о строении химических синапсов Химические синапсы представляют собой окончания аксона (терминальные синапсы) или его варикозную часть (проходящие синапсы). Он состоит из трех компонентов пресинаптической части постсинаптической части синаптической щели.
 Химический синапс
Химический синапс
 СТРУКТУРА И МЕХАНИЗМ ДЕЙСТВИЯ СИНАПСА
СТРУКТУРА И МЕХАНИЗМ ДЕЙСТВИЯ СИНАПСА
 синапс.") Аксосоматический синапс. (аксосоматический) синапс.
Аксосоматический синапс. (аксосоматический) синапс.
. Под") Общие представления о механизмах функционирования химических синапсов В пресинаптической части содержится медиатор (трансмиттер). Под влиянием нервного импульса он выделяется в синаптическую щель и, связываясь с рецепторами в постсинаптической части, вызывая изменение ионной проницаемости постинаптической мембраны , Это приводит к ее деполяризации (в возбуждающих синапсах) или гиперполяризации (в тормозных синапсах).
Общие представления о механизмах функционирования химических синапсов В пресинаптической части содержится медиатор (трансмиттер). Под влиянием нервного импульса он выделяется в синаптическую щель и, связываясь с рецепторами в постсинаптической части, вызывая изменение ионной проницаемости постинаптической мембраны , Это приводит к ее деполяризации (в возбуждающих синапсах) или гиперполяризации (в тормозных синапсах).
 В ней содержатся митохондрии, агранулярная эндоплазматическая сеть, нейрофиламенты,") Пресинаптическая часть (или пресинаптический элемент, пресинпас) В ней содержатся митохондрии, агранулярная эндоплазматическая сеть, нейрофиламенты, нейротрубочки и синаптические пузырьки диаметром 20 65 нм, в которых находится нейромедиатор.
Пресинаптическая часть (или пресинаптический элемент, пресинпас) В ней содержатся митохондрии, агранулярная эндоплазматическая сеть, нейрофиламенты, нейротрубочки и синаптические пузырьки диаметром 20 65 нм, в которых находится нейромедиатор.
 Пресинаптическая часть Форма и характер содержимого пузырьков зависят от находящихся в них нейромедиаторов. круглые светлые пузырьки содержат ацетилхолин, пузырьки с компактным плотным центром содержат норадреналин, крупные плотные пузырьки со светлым подмембранным ободком содержат пептиды.
Пресинаптическая часть Форма и характер содержимого пузырьков зависят от находящихся в них нейромедиаторов. круглые светлые пузырьки содержат ацетилхолин, пузырьки с компактным плотным центром содержат норадреналин, крупные плотные пузырьки со светлым подмембранным ободком содержат пептиды.
 Пресинаптическая часть Медиаторы вырабатываются в теле нейрона и механизмом быстрого аксонного транспорта переносятся в окончания аксона, где происходит их депонирование. Частично синаптические пузырьки образуются в самом синапсе путем отщепления от цистерн агранулярной эндоплазматической сети.
Пресинаптическая часть Медиаторы вырабатываются в теле нейрона и механизмом быстрого аксонного транспорта переносятся в окончания аксона, где происходит их депонирование. Частично синаптические пузырьки образуются в самом синапсе путем отщепления от цистерн агранулярной эндоплазматической сети.
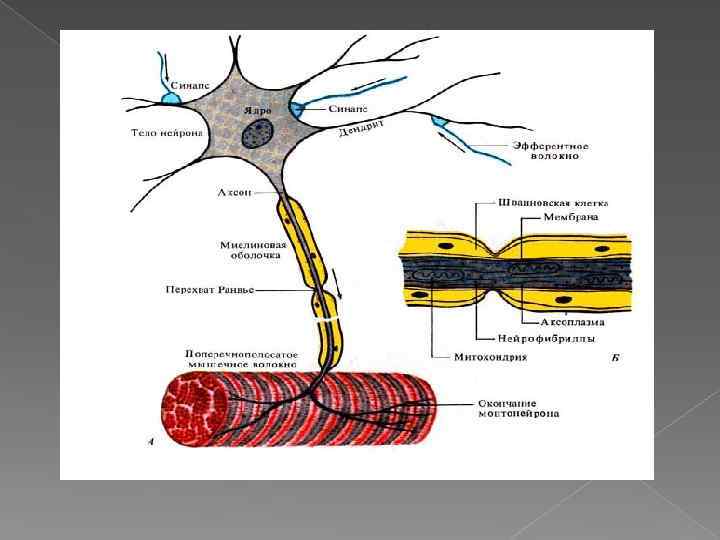
 Пресинаптическая часть На внутренней стороне. пресинаптической мембраны имеется пресинаптическое уплотнение, образованное фибриллярной гексагональной белковой сетью Ее ячейки способствуют равномерному распределению синаптических пузырьков по поверхности мембраны.
Пресинаптическая часть На внутренней стороне. пресинаптической мембраны имеется пресинаптическое уплотнение, образованное фибриллярной гексагональной белковой сетью Ее ячейки способствуют равномерному распределению синаптических пузырьков по поверхности мембраны.
 представлена постсинаптической мембраной, Она содержит особые комплексы интегральных белков") Постсинаптическая часть (или постсинаптический элемент) представлена постсинаптической мембраной, Она содержит особые комплексы интегральных белков – синаптические рецепторы (мембранные рецепторы). Эти рецепторы связываются с нейромедиатором.
Постсинаптическая часть (или постсинаптический элемент) представлена постсинаптической мембраной, Она содержит особые комплексы интегральных белков – синаптические рецепторы (мембранные рецепторы). Эти рецепторы связываются с нейромедиатором.
 СТРУКТУРА И МЕХАНИЗМ ДЕЙСТВИЯ СИНАПСА
СТРУКТУРА И МЕХАНИЗМ ДЕЙСТВИЯ СИНАПСА
 внесинаптические области, По обе стороны от постсинаптической мембраны располагаются внесинаптические области, Они играют важную роль в процессах дальнейшего проведения возбуждения от постсинаптической мембраны.
внесинаптические области, По обе стороны от постсинаптической мембраны располагаются внесинаптические области, Они играют важную роль в процессах дальнейшего проведения возбуждения от постсинаптической мембраны.
 Синаптическая щель – важный компонент синапса. Ее ширина 20 50 нм. Во многих синапсах щель содержит поперечно расположенные гликопротеиновые интрасинаптические филаменты толщиной 5 нм, которые обеспечивают адгезивные связи пре и постсинаптических частей, а также направленную диффузию медиатора.
Синаптическая щель – важный компонент синапса. Ее ширина 20 50 нм. Во многих синапсах щель содержит поперечно расположенные гликопротеиновые интрасинаптические филаменты толщиной 5 нм, которые обеспечивают адгезивные связи пре и постсинаптических частей, а также направленную диффузию медиатора.
 . Этапы синаптической передачи
. Этапы синаптической передачи
, распространяющегося") Механизм функционирования химического синапса. 1. Активация синапса происходит под влиянием потенциала действия (ПД), распространяющегося к пресинапсу от тела нейрона. 2. Под влиянием ПД происходит деполяризация пресинаптической мембраны
Механизм функционирования химического синапса. 1. Активация синапса происходит под влиянием потенциала действия (ПД), распространяющегося к пресинапсу от тела нейрона. 2. Под влиянием ПД происходит деполяризация пресинаптической мембраны
 Механизм функционирования химического синапса. 3. Это повышает проницаемость кальциевых каналов пресинаптической мембраны и приводит к увеличению входа в пресинапс ионов Са 2+, что вызывает освобождение (выход из пресинапса) 100 200 порций, или квантов, медиатора в синаптическую щель (путем экзоцитоза)
Механизм функционирования химического синапса. 3. Это повышает проницаемость кальциевых каналов пресинаптической мембраны и приводит к увеличению входа в пресинапс ионов Са 2+, что вызывает освобождение (выход из пресинапса) 100 200 порций, или квантов, медиатора в синаптическую щель (путем экзоцитоза)
 Механизм функционирования химического синапса. 4. В синаптической щели медиатор взаимодействует со специфическими рецепторами постсинаптической мембраны 4 а. Во многих синапсах основная масса рецепторов – это ионотропные рецепторы. Под влиянием медиатора они изменяют проницаемость ионных каналов постсинаптической мембраны.
Механизм функционирования химического синапса. 4. В синаптической щели медиатор взаимодействует со специфическими рецепторами постсинаптической мембраны 4 а. Во многих синапсах основная масса рецепторов – это ионотропные рецепторы. Под влиянием медиатора они изменяют проницаемость ионных каналов постсинаптической мембраны.
 Механизм функционирования химического синапса. 4 б. В ряде синапсов имеются метаботропные рецепторы. При их активации (за счет вторичных посредников и протеинкиназ) меняется активность внутриклеточных белков эффекторов, в том числе ионных каналов и ионных насосов.
Механизм функционирования химического синапса. 4 б. В ряде синапсов имеются метаботропные рецепторы. При их активации (за счет вторичных посредников и протеинкиназ) меняется активность внутриклеточных белков эффекторов, в том числе ионных каналов и ионных насосов.
 Механизм функционирования химического синапса. 5. В целом, взаимодействие медиатора с постсинаптическими рецепторами изменяет ионную проницаемость. 6. В синапсах, в которых осуществляется возбуждение постсинаптической структуры, обычно происходит повышение проницаемости для ионов Na+ или Cа 2+, что вызывает деполяризацию постсинаптической мембраны.
Механизм функционирования химического синапса. 5. В целом, взаимодействие медиатора с постсинаптическими рецепторами изменяет ионную проницаемость. 6. В синапсах, в которых осуществляется возбуждение постсинаптической структуры, обычно происходит повышение проницаемости для ионов Na+ или Cа 2+, что вызывает деполяризацию постсинаптической мембраны.
 Механизм функционирования химического синапса. 7. Эта деполяризация получила название – возбуждающий постсинаптический потенциал, или ВПСП (в нервно мышечном синапсе – потенциал концевой пластинки, или ПКП). 8. Если величина ВПСП достигает критического уровня деполяризации, то во внесинаптических областях генерируется ПД
Механизм функционирования химического синапса. 7. Эта деполяризация получила название – возбуждающий постсинаптический потенциал, или ВПСП (в нервно мышечном синапсе – потенциал концевой пластинки, или ПКП). 8. Если величина ВПСП достигает критического уровня деполяризации, то во внесинаптических областях генерируется ПД
 мв ДЕПОЛЯРИЗАЦИЯ Na+ - 70 - 85 - 90") Возбуждающий постсинаптический потенциал ( ВПСП) мв ДЕПОЛЯРИЗАЦИЯ Na+ - 70 - 85 - 90 мс 0 4 8 12
Возбуждающий постсинаптический потенциал ( ВПСП) мв ДЕПОЛЯРИЗАЦИЯ Na+ - 70 - 85 - 90 мс 0 4 8 12
 Локальные ответы и закон силы для синапса
Локальные ответы и закон силы для синапса
 Закон силы
Закон силы
 . Механизм функционирования химического синапса 9. В тормозных синапсах в результате взаимодействия медиатора с рецепторами, наоборот, происходит гиперполяризация (за счет, например, увеличения проницаемости для ионов калия и/или хлора). 10. Этот вид изменения мембранного потенциала получил название «тормозной постсинаптический потенциал, или ТПСП» .
. Механизм функционирования химического синапса 9. В тормозных синапсах в результате взаимодействия медиатора с рецепторами, наоборот, происходит гиперполяризация (за счет, например, увеличения проницаемости для ионов калия и/или хлора). 10. Этот вид изменения мембранного потенциала получил название «тормозной постсинаптический потенциал, или ТПСП» .
 мв 0 4 6 8 - 90 -") Тормозной постсинаптический потенциал ( ТПСП ) мв 0 4 6 8 - 90 - 94 мс ГИПЕРПОЛЯРИЗАЦИЯ К+ Cl
Тормозной постсинаптический потенциал ( ТПСП ) мв 0 4 6 8 - 90 - 94 мс ГИПЕРПОЛЯРИЗАЦИЯ К+ Cl
 Механизм функционирования химического синапса. 11. В гиперполяризованном состоянии клетка снижает свою возбудимость и благодаря этому прекращает отвечать на внешние раздражители или (если она обладала свойством автоматии) уменьшает спонтанную активность
Механизм функционирования химического синапса. 11. В гиперполяризованном состоянии клетка снижает свою возбудимость и благодаря этому прекращает отвечать на внешние раздражители или (если она обладала свойством автоматии) уменьшает спонтанную активность
 . Механизм функционирования химического синапса . 12. Выделившийся в синаптическую щель медиатор может взаимодействовать с рецепторами, расположенными на пресинаптической мембране. Тем самым он может регулировать интенсивность последующего высвобождения медиатора, т. е. процесс экзоцитоза. Это получило название антидромного эффекта или явления обратной связи.
. Механизм функционирования химического синапса . 12. Выделившийся в синаптическую щель медиатор может взаимодействовать с рецепторами, расположенными на пресинаптической мембране. Тем самым он может регулировать интенсивность последующего высвобождения медиатора, т. е. процесс экзоцитоза. Это получило название антидромного эффекта или явления обратной связи.
 Химический синапс
Химический синапс
 Механизм функционирования химического синапса. 13. После каждого цикла проведения нервного импульса медиатор разрушается и с участием специфического фермента подвергается удалению. Так, ацетилхолин (АХ) разрушается ацетилхолинэстеразой (АХЭ), норадреналин (НА) разрушается моноаминоксидазой (МАО) и/или катехол 0 метилтрансферазой (КОМТ).
Механизм функционирования химического синапса. 13. После каждого цикла проведения нервного импульса медиатор разрушается и с участием специфического фермента подвергается удалению. Так, ацетилхолин (АХ) разрушается ацетилхолинэстеразой (АХЭ), норадреналин (НА) разрушается моноаминоксидазой (МАО) и/или катехол 0 метилтрансферазой (КОМТ).
 или продуктов") Механизм функционирования химического синапса. 14. Одновременно происходит обратный захват медиатора (например, норадреналина) или продуктов расщепления медиатора (например, холина при разрушении ацетилхолина) в пресинаптическую структуру (нейрональный захват) либо в пост –и внесинаптическую структуры (экстранейрональный захват).
Механизм функционирования химического синапса. 14. Одновременно происходит обратный захват медиатора (например, норадреналина) или продуктов расщепления медиатора (например, холина при разрушении ацетилхолина) в пресинаптическую структуру (нейрональный захват) либо в пост –и внесинаптическую структуры (экстранейрональный захват).
 Химический синапс
Химический синапс
 Механизм функционирования химического синапса. 15. Снижение концентрации медиатора в синаптической щели достигается также путем его простой диффузии во внесинаптические пространства.
Механизм функционирования химического синапса. 15. Снижение концентрации медиатора в синаптической щели достигается также путем его простой диффузии во внесинаптические пространства.
 . Этапы синаптической передачи
. Этапы синаптической передачи
 Синтез медиатора происходит в пресинаптическом элементе. В него из крови или спинномозговой жидкости попадают исходные продукты (предшественники медиаторов) и ферменты, необходимые для его синтеза. Ферменты образуются в соме нейрона и по аксону со скоростью 6 мм/сутки транспортируются в пресинаптическое окончание аксона,
Синтез медиатора происходит в пресинаптическом элементе. В него из крови или спинномозговой жидкости попадают исходные продукты (предшественники медиаторов) и ферменты, необходимые для его синтеза. Ферменты образуются в соме нейрона и по аксону со скоростью 6 мм/сутки транспортируются в пресинаптическое окончание аксона,
 Синтез медиатора Угнетение активности этих ферментов фармакологическим путем может привести к истощению запасов медиатора в синапсе, т. е. к снижению его функциональной способности. Образовавшийся медиатор путем активного транспорта вводится в синаптические везикулы (мелкие везикулы).
Синтез медиатора Угнетение активности этих ферментов фармакологическим путем может привести к истощению запасов медиатора в синапсе, т. е. к снижению его функциональной способности. Образовавшийся медиатор путем активного транспорта вводится в синаптические везикулы (мелкие везикулы).
 . Этапы синаптической передачи
. Этапы синаптической передачи
 Синаптические везикулы и их типы Любой медиатор в пресинаптических нервных окончаниях содержится в везикулах. Выделяют два типа везикул 1) мелкие везикулы (диаметром около 50 нм); они однородны по размерам и содержат классический медиатор. 2) крупные везикулы (диаметром около 100 нм); они неоднородны по размерам и содержат пептиды, которые помимо медиаторной функции могут модулировать синаптическую передачу
Синаптические везикулы и их типы Любой медиатор в пресинаптических нервных окончаниях содержится в везикулах. Выделяют два типа везикул 1) мелкие везикулы (диаметром около 50 нм); они однородны по размерам и содержат классический медиатор. 2) крупные везикулы (диаметром около 100 нм); они неоднородны по размерам и содержат пептиды, которые помимо медиаторной функции могут модулировать синаптическую передачу
 Синаптические везикулы и их типы Все синаптические везикулы образуются в теле нервной клетки из эндоплазматического ретикулюма и цистерн аппарата Гольджи. Затем они транспортируются по аксону в нервные окончания. Крупные везикулы заполняются медиатором (пептидом) непосредственно в соме нейрона.
Синаптические везикулы и их типы Все синаптические везикулы образуются в теле нервной клетки из эндоплазматического ретикулюма и цистерн аппарата Гольджи. Затем они транспортируются по аксону в нервные окончания. Крупные везикулы заполняются медиатором (пептидом) непосредственно в соме нейрона.
 Синаптические везикулы и их типы Мелкие везикулы содержат АТФ, ионы, ферменты, а их мембрана имеет протоновые насосы. С участием этих насосов (главным образом, в пресинаптическом окончании) происходит наполнение мелких везикул медиатором, концентрация которого достигает в них высоких значений (около 100 ммоль/л).
Синаптические везикулы и их типы Мелкие везикулы содержат АТФ, ионы, ферменты, а их мембрана имеет протоновые насосы. С участием этих насосов (главным образом, в пресинаптическом окончании) происходит наполнение мелких везикул медиатором, концентрация которого достигает в них высоких значений (около 100 ммоль/л).
 Синаптические везикулы и их типы. Пулы Мелкие везикулы в пресинаптическом окончании формируют два основных пула: пул 1 пул 2.
Синаптические везикулы и их типы. Пулы Мелкие везикулы в пресинаптическом окончании формируют два основных пула: пул 1 пул 2.
 Синаптические везикулы и их типы Пул 1 – это относительно небольшой пул, готовый к немедленному экзоцитозу, т. е. это запас медиатора, способного освободиться в течение короткого промежутка времени. Пул 1 содержит 10 50 везикул и при истощении пополняется в течение 5 12 секунд.
Синаптические везикулы и их типы Пул 1 – это относительно небольшой пул, готовый к немедленному экзоцитозу, т. е. это запас медиатора, способного освободиться в течение короткого промежутка времени. Пул 1 содержит 10 50 везикул и при истощении пополняется в течение 5 12 секунд.
 Синаптические везикулы и их типы Вероятность освобождения кванта из пула 1 пропорциональна количеству везикул в пуле. Размер пула 1 является постоянной величиной и определяет эффективность работы синапса.
Синаптические везикулы и их типы Вероятность освобождения кванта из пула 1 пропорциональна количеству везикул в пуле. Размер пула 1 является постоянной величиной и определяет эффективность работы синапса.
 Синаптические везикулы и их типы Пул 2 – это относительно большой пул, везикулы которого не участвуют в секреции, но могут перемещаться по мере необходимости в пул 1, т. е. пул 2 – это мобилизационный запас медиатора, упакованного в везикулу. Везикулы этого пула с определенной скоростью могут пополнять запас доступного медиатора Переход везикул из пула 2 в пул 1 регулируется внутриклеточной концентрацией ионов кальция.
Синаптические везикулы и их типы Пул 2 – это относительно большой пул, везикулы которого не участвуют в секреции, но могут перемещаться по мере необходимости в пул 1, т. е. пул 2 – это мобилизационный запас медиатора, упакованного в везикулу. Везикулы этого пула с определенной скоростью могут пополнять запас доступного медиатора Переход везикул из пула 2 в пул 1 регулируется внутриклеточной концентрацией ионов кальция.
 Пути освобождения медиатора из синапса. В основе освобождения медиатора лежит процесс экзоцитоза Существуют два механизма освобождения медиатора и всего содержимого везикулы в синаптическую щель.
Пути освобождения медиатора из синапса. В основе освобождения медиатора лежит процесс экзоцитоза Существуют два механизма освобождения медиатора и всего содержимого везикулы в синаптическую щель.
 Пути освобождения медиатора из синапса Первый механизм – это классический экзоцитоз, при котором происходит полное слияние везикулы с пресинаптической мембраной. В этом случае все содержимое везикулы (медиатор, АТФ, ионы, ассоциированные белки и ферменты) оказывается в синаптической щели.
Пути освобождения медиатора из синапса Первый механизм – это классический экзоцитоз, при котором происходит полное слияние везикулы с пресинаптической мембраной. В этом случае все содержимое везикулы (медиатор, АТФ, ионы, ассоциированные белки и ферменты) оказывается в синаптической щели.
 Пути освобождения медиатора из синапса Второй механизм – это экзоцитоз, протекающий без полного слияния везикулы с пресинаптической мембраной и с частичным освобождением медиатора. Он характеризуется формированием временной поры (канала) в пресинаптической мембране, .
Пути освобождения медиатора из синапса Второй механизм – это экзоцитоз, протекающий без полного слияния везикулы с пресинаптической мембраной и с частичным освобождением медиатора. Он характеризуется формированием временной поры (канала) в пресинаптической мембране, .
 Пути освобождения медиатора из синапса За счет этой поры полость везикулы сообщается с синаптической щелью: при открытии поры (в момент контакта с пресинаптической мембраной). При этом везикула при каждом контакте с пресинаптической мембраной теряет только часть своего содержимого. При таком способе функционирования везикула может участвовать в экзоцитозе многократно.
Пути освобождения медиатора из синапса За счет этой поры полость везикулы сообщается с синаптической щелью: при открытии поры (в момент контакта с пресинаптической мембраной). При этом везикула при каждом контакте с пресинаптической мембраной теряет только часть своего содержимого. При таком способе функционирования везикула может участвовать в экзоцитозе многократно.
 Образование, транспорт и экзоцитоз синаптических пузырьков
Образование, транспорт и экзоцитоз синаптических пузырьков
 Организация секреции медиатора. Активная зона. Процессы экзоцитоза в синапсе осуществляются в специализированных пресинаптических структурах, которые получили название активных зон. В каждом синапсе число таких зон достигает 1 5. Активная зона имеет длину 1 3 мкм, ширину – 0, 1 мкм; от соседней активной зоны она отдалена на расстоянии 1 2 мкм.
Организация секреции медиатора. Активная зона. Процессы экзоцитоза в синапсе осуществляются в специализированных пресинаптических структурах, которые получили название активных зон. В каждом синапсе число таких зон достигает 1 5. Активная зона имеет длину 1 3 мкм, ширину – 0, 1 мкм; от соседней активной зоны она отдалена на расстоянии 1 2 мкм.
 Организация секреции медиатора. Активная зона. Каждая активная зона состоит из пресинаптической плотной полоски, около которой рядами расположены кальциевые каналы (белки), два ряда синаптических везикул, а также специализированные белки экзоцитоза (в том числе соединяющие везикулу с пресинаптической мембраной) и элементы цитоскелета, т. е. сеть актиновых филаментов и микротрубочек.
Организация секреции медиатора. Активная зона. Каждая активная зона состоит из пресинаптической плотной полоски, около которой рядами расположены кальциевые каналы (белки), два ряда синаптических везикул, а также специализированные белки экзоцитоза (в том числе соединяющие везикулу с пресинаптической мембраной) и элементы цитоскелета, т. е. сеть актиновых филаментов и микротрубочек.
 Активные зоны нервно мышечного синапса
Активные зоны нервно мышечного синапса
 Организация секреции медиатора. Активная зона. Везикулы связываются с элементами цитоскелета и между собой с участием белка (синапсин I). Элементы цитоскелета удерживают синаптические везикулы в пуле 1 (и пуле 2, т. е. в резервном пуле) и ограничивают их избыточную мобилизацию, регулируют процессы освобождения, обеспечивают характерную пространственную организацию активных зон и рециркулирование мембраны синаптической везикулы после экзоцитоза.
Организация секреции медиатора. Активная зона. Везикулы связываются с элементами цитоскелета и между собой с участием белка (синапсин I). Элементы цитоскелета удерживают синаптические везикулы в пуле 1 (и пуле 2, т. е. в резервном пуле) и ограничивают их избыточную мобилизацию, регулируют процессы освобождения, обеспечивают характерную пространственную организацию активных зон и рециркулирование мембраны синаптической везикулы после экзоцитоза.
 везикулы") Функционирование активной зоны. В активной зоне происходят последовательно 5 процессов. 1. Транспорт (мобилизация) везикулы из пула 2 в пул 1 за счет элементов цитоскелета. 2. Стыковка (докирование) везикулы с местом освобождения в активной зоне.
Функционирование активной зоны. В активной зоне происходят последовательно 5 процессов. 1. Транспорт (мобилизация) везикулы из пула 2 в пул 1 за счет элементов цитоскелета. 2. Стыковка (докирование) везикулы с местом освобождения в активной зоне.
 Активные зоны нервно мышечного синапса
Активные зоны нервно мышечного синапса
 везикулы к экзоцитозу. Она заключается в трансформации белкового") Функционирование активной зоны. 3. Подготовка (прайминг) везикулы к экзоцитозу. Она заключается в трансформации белкового комплекса экзоцитоза, в результате чего везикула готова к выбросу своего содержимого и ожидает лишь входа ионов кальция.
Функционирование активной зоны. 3. Подготовка (прайминг) везикулы к экзоцитозу. Она заключается в трансформации белкового комплекса экзоцитоза, в результате чего везикула готова к выбросу своего содержимого и ожидает лишь входа ионов кальция.
 Функционирование активной зоны. 4. Экзоцитоз – слияние мембраны везикул с плазматической мембраной. Для осуществления этого процесса необходимо открытие потенциалозависимых Са каналов и увеличение концентрации кальция в области везикулы. 5. Рециклизация везикулы посредством эндоцитоза с последующим заполнением медиатором и включением ее в пулы 1 или 2.
Функционирование активной зоны. 4. Экзоцитоз – слияние мембраны везикул с плазматической мембраной. Для осуществления этого процесса необходимо открытие потенциалозависимых Са каналов и увеличение концентрации кальция в области везикулы. 5. Рециклизация везикулы посредством эндоцитоза с последующим заполнением медиатором и включением ее в пулы 1 или 2.
 Образование, транспорт и экзоцитоз синаптических пузырьков
Образование, транспорт и экзоцитоз синаптических пузырьков
 Секретосома – это синаптическая везикула, докированная в области активной зоны, а также кальциевый канал (каналы) и белки, ответственные за экзоцитоз и эндоцитоз, т. е. белковый комплекс. Секретосомы представляют собой функциональную единицу активной зоны, или пула
Секретосома – это синаптическая везикула, докированная в области активной зоны, а также кальциевый канал (каналы) и белки, ответственные за экзоцитоз и эндоцитоз, т. е. белковый комплекс. Секретосомы представляют собой функциональную единицу активной зоны, или пула
 Секретосома Количество секретосом в отдельной активной зоне определяется размерами активной зоны (обычно оно равно нескольким десяткам). Каждая секретосома может участвовать в вызванном и спонтанном высвобождении медиатора. Область, в которой повышена концентрация ионов кальция, и где происходит экзоцитоз, называется кальциевым микродоменом.
Секретосома Количество секретосом в отдельной активной зоне определяется размерами активной зоны (обычно оно равно нескольким десяткам). Каждая секретосома может участвовать в вызванном и спонтанном высвобождении медиатора. Область, в которой повышена концентрация ионов кальция, и где происходит экзоцитоз, называется кальциевым микродоменом.
 Белковый аппарат экзоцитоза и эндоцитоза. Различают 8 различных белков, имеющих отношение к процессам экзоцитоза медиатора и эндоцитоза везикулы. 1. Синаптотагмин (65 к. Д) – белок, связанный с везикулой и предназначенный для связывания ионов кальция (кальциевый сенсор). 2. Синаптобревин (18 к. Д) – белок, связанный с везикулой и предназначенный для образования комплекса (везикула + пресинаптическая мембрана).
Белковый аппарат экзоцитоза и эндоцитоза. Различают 8 различных белков, имеющих отношение к процессам экзоцитоза медиатора и эндоцитоза везикулы. 1. Синаптотагмин (65 к. Д) – белок, связанный с везикулой и предназначенный для связывания ионов кальция (кальциевый сенсор). 2. Синаптобревин (18 к. Д) – белок, связанный с везикулой и предназначенный для образования комплекса (везикула + пресинаптическая мембрана).
 – белок, связывающий") Белковый аппарат экзоцитоза и эндоцитоза. 3. Синапсин I (86 к. Д) – белок, связывающий везикулы с цитоскелетом и тем самым участвующий в процессах мобилизации везикул. 4. Синтаксин (35 к. Д) – белок пресинаптической мембраны, связанный с кальциевыми каналами; он способен связываться с синаптотагмином.
Белковый аппарат экзоцитоза и эндоцитоза. 3. Синапсин I (86 к. Д) – белок, связывающий везикулы с цитоскелетом и тем самым участвующий в процессах мобилизации везикул. 4. Синтаксин (35 к. Д) – белок пресинаптической мембраны, связанный с кальциевыми каналами; он способен связываться с синаптотагмином.
 –") Белковый аппарат экзоцитоза и эндоцитоза. 5. Rab 3 A (20 25 к. Д) – белок, препятствующий освобождению медиатора из активной зоны одним квантом. 6. NSF, или N этил малеимид чувствительный фактор; это цитозольный белок, который состоит из 4 субъединиц (76 к. Д); он обладает АТФ азной активностью и обеспечивает энергией различные внутриклеточные процессы, во время которых происходит слияние мембран и процессы экзо и эндоцитоза.
Белковый аппарат экзоцитоза и эндоцитоза. 5. Rab 3 A (20 25 к. Д) – белок, препятствующий освобождению медиатора из активной зоны одним квантом. 6. NSF, или N этил малеимид чувствительный фактор; это цитозольный белок, который состоит из 4 субъединиц (76 к. Д); он обладает АТФ азной активностью и обеспечивает энергией различные внутриклеточные процессы, во время которых происходит слияние мембран и процессы экзо и эндоцитоза.
 Белковый аппарат экзоцитоза и эндоцитоза. 7. SNAP – растворимый цитозольный белок, связывающий NSF; он участвует в синаптической передаче после докирования везикулы. 8. SNAP 25 – это белок, связанный с секретосомой. Он расположен на пресинаптической мембране. Вместе с синаптобревином и синтаксином этот белок формирует комплекс, необходимый для экзоцитоза.
Белковый аппарат экзоцитоза и эндоцитоза. 7. SNAP – растворимый цитозольный белок, связывающий NSF; он участвует в синаптической передаче после докирования везикулы. 8. SNAP 25 – это белок, связанный с секретосомой. Он расположен на пресинаптической мембране. Вместе с синаптобревином и синтаксином этот белок формирует комплекс, необходимый для экзоцитоза.
. Для объяснения экзоцитоза была предложена SNARE") Функция белкового аппарата, ответственного за экзоцитоз (SNARE гипотеза). Для объяснения экзоцитоза была предложена SNARE гипотеза, или гипотеза об универсальной единице докирования и слияния. Гипотеза предполагает, что в процессе докирования и слияния участвуют такие белки как v SNARE (или везикулярный SNAP рецептор), t SNARE, или мембранный рецептор для SNAP (он является мишенью для v SNARE), цитозольные белки NSF и SNAP белок (адаптор к NSF).
Функция белкового аппарата, ответственного за экзоцитоз (SNARE гипотеза). Для объяснения экзоцитоза была предложена SNARE гипотеза, или гипотеза об универсальной единице докирования и слияния. Гипотеза предполагает, что в процессе докирования и слияния участвуют такие белки как v SNARE (или везикулярный SNAP рецептор), t SNARE, или мембранный рецептор для SNAP (он является мишенью для v SNARE), цитозольные белки NSF и SNAP белок (адаптор к NSF).
. Докирование везикулы осуществляется благодаря комплементарности v") Функция белкового аппарата, ответственного за экзоцитоз (SNARE гипотеза). Докирование везикулы осуществляется благодаря комплементарности v SNARE и t SNARE. Эти два белка рецептора взаимодействуют друг с другом по принципу «антиген антитело» .
Функция белкового аппарата, ответственного за экзоцитоз (SNARE гипотеза). Докирование везикулы осуществляется благодаря комплементарности v SNARE и t SNARE. Эти два белка рецептора взаимодействуют друг с другом по принципу «антиген антитело» .
. После контакта, т. е. когда они") Функция белкового аппарата, ответственного за экзоцитоз (SNARE гипотеза). После контакта, т. е. когда они свяжутся между собой, оба этих белка становятся рецепторами для SNAP, который в свою очередь присоединяет NSF. При этом комплекс v SNARE + t SNARE + Са канал является осью, на которую нанизываются другие молекулы, участвующие в экзоцитозе.
Функция белкового аппарата, ответственного за экзоцитоз (SNARE гипотеза). После контакта, т. е. когда они свяжутся между собой, оба этих белка становятся рецепторами для SNAP, который в свою очередь присоединяет NSF. При этом комплекс v SNARE + t SNARE + Са канал является осью, на которую нанизываются другие молекулы, участвующие в экзоцитозе.
. Гидролиз АТФ с участием NSF ведет") Функция белкового аппарата, ответственного за экзоцитоз (SNARE гипотеза). Гидролиз АТФ с участием NSF ведет к пространственной реорганизации SNARE комплекса, что приводит синаптическую везикулу в состояние прайминга. Вход кальция дестабилизирует SNARE комплекс везикулы и приводит к экзоцитозу
Функция белкового аппарата, ответственного за экзоцитоз (SNARE гипотеза). Гидролиз АТФ с участием NSF ведет к пространственной реорганизации SNARE комплекса, что приводит синаптическую везикулу в состояние прайминга. Вход кальция дестабилизирует SNARE комплекс везикулы и приводит к экзоцитозу
. В целом, последовательность событий от момента") Функция белкового аппарата, ответственного за экзоцитоз (SNARE гипотеза). В целом, последовательность событий от момента прихода везикулы в пул 1 до момента ее прайминга можно представить следующим образом: v SNARE + t SNARE → докирование везикулы → v SNARE + t SNARE + SNAP → прайминг везикулы.
Функция белкового аппарата, ответственного за экзоцитоз (SNARE гипотеза). В целом, последовательность событий от момента прихода везикулы в пул 1 до момента ее прайминга можно представить следующим образом: v SNARE + t SNARE → докирование везикулы → v SNARE + t SNARE + SNAP → прайминг везикулы.
 Роль ионов кальция в экзоцитозе Нейрональные Са каналы являются гетерометрическими белками, которые состоят из трех субъединиц. В условиях покоя с участием Са каналов осуществляется докирование синаптической везикулы в активной зоне, что реализуется совместно с белковым аппаратом экзоцитоза, т. е. синтаксином и SNAP 25. Тем самым предотвращается случайный экзоцитоз везикул.
Роль ионов кальция в экзоцитозе Нейрональные Са каналы являются гетерометрическими белками, которые состоят из трех субъединиц. В условиях покоя с участием Са каналов осуществляется докирование синаптической везикулы в активной зоне, что реализуется совместно с белковым аппаратом экзоцитоза, т. е. синтаксином и SNAP 25. Тем самым предотвращается случайный экзоцитоз везикул.
") Роль ионов кальция в экзоцитозе При деполяризации пресинаптической мембраны под влиянием потенциала действия (ПД) Са каналы, находящиеся рядом с докированной везикулой, открываются. Ионы Са 2+ входят в пресинапс. Это создает около везикулы высокую концентрацию Са 2+ (более 100 мкмоль/л), т. е. кальциевый микродомен.
Роль ионов кальция в экзоцитозе При деполяризации пресинаптической мембраны под влиянием потенциала действия (ПД) Са каналы, находящиеся рядом с докированной везикулой, открываются. Ионы Са 2+ входят в пресинапс. Это создает около везикулы высокую концентрацию Са 2+ (более 100 мкмоль/л), т. е. кальциевый микродомен.
 Роль ионов кальция в экзоцитозе Са 2+ взаимодействует с Са сенсором везикулы и тем самым инициирует ее экзоцитоз, в частности Са 2+ активирует АТФ азную активность NSF, благодаря чему совершается контакт везикулы с пресинаптической мембраной и выброс медиатора в синаптическую щель.
Роль ионов кальция в экзоцитозе Са 2+ взаимодействует с Са сенсором везикулы и тем самым инициирует ее экзоцитоз, в частности Са 2+ активирует АТФ азную активность NSF, благодаря чему совершается контакт везикулы с пресинаптической мембраной и выброс медиатора в синаптическую щель.
 Роль ионов кальция в экзоцитозе Созданию кальциевого микродомена способствуют митохондрии и эндоплазматический ретикулюм везикулы (как депо Са 2+). Чем дольше существует кальциевый микродомен, тем длительнее и эффективнее экзоцитоз везикулы
Роль ионов кальция в экзоцитозе Созданию кальциевого микродомена способствуют митохондрии и эндоплазматический ретикулюм везикулы (как депо Са 2+). Чем дольше существует кальциевый микродомен, тем длительнее и эффективнее экзоцитоз везикулы
 Синтез рецепторов постсинаптической мембраны. Все рецепторы постсинаптической мембраны интегральные белки плазматической мембраны. Они синтезируются в рибосамах эндоплазматического ретикулюма клетки После «сортировки» в аппарате Гольджи они встраиваются в постсинаптическую мембрану
Синтез рецепторов постсинаптической мембраны. Все рецепторы постсинаптической мембраны интегральные белки плазматической мембраны. Они синтезируются в рибосамах эндоплазматического ретикулюма клетки После «сортировки» в аппарате Гольджи они встраиваются в постсинаптическую мембрану
 Синтез рецепторов постсинаптической мембраны. Синтез рецепторов контролируется йодсодержащими гормонами щитовидной железы и, вероятно, другими гормонами Синтез рецепторов зависит от наличием медиатора. Например, при денервации мышцы число Н холинорецепторов в области постсинаптической мембраны снижается
Синтез рецепторов постсинаптической мембраны. Синтез рецепторов контролируется йодсодержащими гормонами щитовидной железы и, вероятно, другими гормонами Синтез рецепторов зависит от наличием медиатора. Например, при денервации мышцы число Н холинорецепторов в области постсинаптической мембраны снижается
 Синтез рецепторов постсинаптической мембраны. При нарушении синтеза рецепторов или при наличии антител к ним, реальная концентрация рецепторов на постсинаптической мембране снижается. Это уменьшает эффективность синаптической передачи.
Синтез рецепторов постсинаптической мембраны. При нарушении синтеза рецепторов или при наличии антител к ним, реальная концентрация рецепторов на постсинаптической мембране снижается. Это уменьшает эффективность синаптической передачи.
") Активация постсинаптических рецепторов Агонисты вещества, способные активировать рецептор. Медиатор частный случай агониста Антагонисты (блокаторы) вещества, способные блокировать взаимодействие агониста с рецепторами Модуляторы–сенсибилизаторы вещества, повышающие эффективность активации рецептора агонистом Лиганды вещества, способные взаимодействовать с рецептором (т. е. это агонисты, антагонисты, модуляторы сенсибилизаторы)
Активация постсинаптических рецепторов Агонисты вещества, способные активировать рецептор. Медиатор частный случай агониста Антагонисты (блокаторы) вещества, способные блокировать взаимодействие агониста с рецепторами Модуляторы–сенсибилизаторы вещества, повышающие эффективность активации рецептора агонистом Лиганды вещества, способные взаимодействовать с рецептором (т. е. это агонисты, антагонисты, модуляторы сенсибилизаторы)
 или") Активация постсинаптических рецепторов При взаимодействии медиатора с рецептором меняется ионная проницаемость (ионотропные рецепторы) или состояние внутриклеточных эффекторов, например, ионных насосов (метаботропныен рецепторы)
Активация постсинаптических рецепторов При взаимодействии медиатора с рецептором меняется ионная проницаемость (ионотропные рецепторы) или состояние внутриклеточных эффекторов, например, ионных насосов (метаботропныен рецепторы)
 для связывания медиатора (агониста).") Активация постсинаптических рецепторов Рецептор постсинаптической мембраны имеет сайт (активный центр) для связывания медиатора (агониста). Рецептор может иметь сайты для связывания модуляторов или комедиаторов, благодаря чему сродство данного рецептора к медиатору может существенно изменяться.
Активация постсинаптических рецепторов Рецептор постсинаптической мембраны имеет сайт (активный центр) для связывания медиатора (агониста). Рецептор может иметь сайты для связывания модуляторов или комедиаторов, благодаря чему сродство данного рецептора к медиатору может существенно изменяться.
 Активация постсинаптических рецепторов Так, в сыворотке крови и ликворе имеется эндогенный сенсибилизатор β адренорецепторов (его функцию выполняют гистидин, триптофан, тирозин), повышающий в 10 100 раз чувствительность этих рецепторов к норадреналину и адреналину.
Активация постсинаптических рецепторов Так, в сыворотке крови и ликворе имеется эндогенный сенсибилизатор β адренорецепторов (его функцию выполняют гистидин, триптофан, тирозин), повышающий в 10 100 раз чувствительность этих рецепторов к норадреналину и адреналину.
 Рис. –адреносенсибилизирующая активность 100 - кратного разведения сыворотки беременных женщин. Механограмма продольной полоски рога матки небеременной крысы. Горизонтальные линии под механограммой обозначают момент воздействия адреналина (10 -8 г/мл) и сыворотки крови.
Рис. –адреносенсибилизирующая активность 100 - кратного разведения сыворотки беременных женщин. Механограмма продольной полоски рога матки небеременной крысы. Горизонтальные линии под механограммой обозначают момент воздействия адреналина (10 -8 г/мл) и сыворотки крови.
 А. Б. В. Рис. Адреномодулирующее разведения сыворотки крови. влияние Механограммы 100 -кратного циркулярной полоски коронарной артерии свиньи (панель А) тонус повышен гиперкалиевым (КСl 25 м. М) раствором Кребса, циркулярной полоски трахеи коровы (панель Б) тонус повышен ацетилхолином (10 -6 г/мл), миокарда лягушки (панель В). Горизонтальные линии под механограммами обозначают момент воздействия гиперкалиевого раствора Кребса, адреналина (10 -8, 10 -7 и 10 -6 г/мл), ацетилхолина, обзидана (10 -8 г/мл) и сыворотки крови.
А. Б. В. Рис. Адреномодулирующее разведения сыворотки крови. влияние Механограммы 100 -кратного циркулярной полоски коронарной артерии свиньи (панель А) тонус повышен гиперкалиевым (КСl 25 м. М) раствором Кребса, циркулярной полоски трахеи коровы (панель Б) тонус повышен ацетилхолином (10 -6 г/мл), миокарда лягушки (панель В). Горизонтальные линии под механограммами обозначают момент воздействия гиперкалиевого раствора Кребса, адреналина (10 -8, 10 -7 и 10 -6 г/мл), ацетилхолина, обзидана (10 -8 г/мл) и сыворотки крови.
 снижает тонус, вызванный ацетилхолином (АХ 6, 10 6") Сыворотка крови практически здоровых людей (сыв) снижает тонус, вызванный ацетилхолином (АХ 6, 10 6 г/мл) Механограмма продольной полоски рога матки крысы Горизонтальные линии отражают момент воздействия ацетилхолина (АХ-6, 10 -6 г/мл) и исследуемого разведения сыворотки крови. Калибровка 10 м. Н/10 мин. Механограмма циркулярной полоски фундуса желудка (панель Б) Горизонтальные линии отражают момент воздействия ацетилхолина (АХ-6, 10 -6 г/мл) и исследуемого разведения сыворотки крови. Калибровка 10 м. Н/10 мин.
Сыворотка крови практически здоровых людей (сыв) снижает тонус, вызванный ацетилхолином (АХ 6, 10 6 г/мл) Механограмма продольной полоски рога матки крысы Горизонтальные линии отражают момент воздействия ацетилхолина (АХ-6, 10 -6 г/мл) и исследуемого разведения сыворотки крови. Калибровка 10 м. Н/10 мин. Механограмма циркулярной полоски фундуса желудка (панель Б) Горизонтальные линии отражают момент воздействия ацетилхолина (АХ-6, 10 -6 г/мл) и исследуемого разведения сыворотки крови. Калибровка 10 м. Н/10 мин.
. Так,") Активация постсинаптических рецепторов Для активации большинства рецепторов необходимо более одной молекулы агониста (медиатора). Так, для активации одного холинорецептора нужны две молекулы ацетилхолина. Это объясняется наличием двух симметричных агонист связывающих альфа субъединиц в составе холинорецептора
Активация постсинаптических рецепторов Для активации большинства рецепторов необходимо более одной молекулы агониста (медиатора). Так, для активации одного холинорецептора нужны две молекулы ацетилхолина. Это объясняется наличием двух симметричных агонист связывающих альфа субъединиц в составе холинорецептора
 G белок, вторичные посредники и протеинкиназы При активации метаботропных рецепторов передача сигнала внутрь клетки осуществляется с участием вторичных посредников – мелких, подвижных молекул (ц. АМФ, ц. ГМФ, инозитол 3 фосфат, диацилглицерол), способных активировать протеинкиназы (ферменты, переносящие фосфатную группу от АТФ на белок мишень). Фосфорилированный белок (ионный канал, ионный насос, фермент метаболической цепочки и др. ) меняет свою активность и тем самым обеспечивает передачу синаптического сигнала.
G белок, вторичные посредники и протеинкиназы При активации метаботропных рецепторов передача сигнала внутрь клетки осуществляется с участием вторичных посредников – мелких, подвижных молекул (ц. АМФ, ц. ГМФ, инозитол 3 фосфат, диацилглицерол), способных активировать протеинкиназы (ферменты, переносящие фосфатную группу от АТФ на белок мишень). Фосфорилированный белок (ионный канал, ионный насос, фермент метаболической цепочки и др. ) меняет свою активность и тем самым обеспечивает передачу синаптического сигнала.
 G белок, вторичные посредники и протеинкиназы Образованию вторичных посредников предшествует передача сигнала от мембранного рецептора через ГТФ связывающий белок (G белок) к внутриклеточному примембранному ферменту (аденилатциклазе или гуанилатциклазе). G белок может активировать или ингибировать этот фермент (разновидности этого белка обозначают соответственно как Gs белок и Gi белок).
G белок, вторичные посредники и протеинкиназы Образованию вторичных посредников предшествует передача сигнала от мембранного рецептора через ГТФ связывающий белок (G белок) к внутриклеточному примембранному ферменту (аденилатциклазе или гуанилатциклазе). G белок может активировать или ингибировать этот фермент (разновидности этого белка обозначают соответственно как Gs белок и Gi белок).
 G белок, вторичные посредники и протеинкиназы Для различных синапсов существует своя последовательность передачи сигнала (медиатор, G белок, вторичный посредник, протеинкиназа, белок эффектор).
G белок, вторичные посредники и протеинкиназы Для различных синапсов существует своя последовательность передачи сигнала (медиатор, G белок, вторичный посредник, протеинкиназа, белок эффектор).
 путь передачи сигнала") G белок, вторичные посредники и протеинкиназы Для моноаминов (адреналина, норадреналина, серотонина) путь передачи сигнала такой: Gs → аденилатциклаза → ц. АМФ → протеинкиназа А → ионные каналы, ферменты обмена углеводов и др.
G белок, вторичные посредники и протеинкиназы Для моноаминов (адреналина, норадреналина, серотонина) путь передачи сигнала такой: Gs → аденилатциклаза → ц. АМФ → протеинкиназа А → ионные каналы, ферменты обмена углеводов и др.
 G белок, вторичные посредники и протеинкиназы Путь передачи сигнала при активации рецепторов ацетилхолином: Gs → гуанилатциклаза → ц. ГМФ → протеинкиназа Г → калиевые ионные каналы, кальциевые насосы.
G белок, вторичные посредники и протеинкиназы Путь передачи сигнала при активации рецепторов ацетилхолином: Gs → гуанилатциклаза → ц. ГМФ → протеинкиназа Г → калиевые ионные каналы, кальциевые насосы.
 белка Gs–белок стимулирует аденилатциклазу Gi белок – ингибирует аденилатциклазу") Виды G белка (ГТФ связывающего) белка Gs–белок стимулирует аденилатциклазу Gi белок – ингибирует аденилатциклазу Gр–белок активирует β изоформу фосфолипазы С
Виды G белка (ГТФ связывающего) белка Gs–белок стимулирует аденилатциклазу Gi белок – ингибирует аденилатциклазу Gр–белок активирует β изоформу фосфолипазы С
 ц. АМФ циклический аденозинмонофосфат (аденозин + рибоза + фосфат)") Вторичные посредники (внутриклеточные медиаторы, мессенджеры) ц. АМФ циклический аденозинмонофосфат (аденозин + рибоза + фосфат) ц. ГМФ циклический гуанозинмонофосфат (гаунозин + рибоза + фосфат) NO оксид азота; Эйказоноиды, т. е. содержащие 20 углеродных атомов простагландины типа Е 2 и F 2 альфа, простациклины, тромбоксаны и лейкотриены
Вторичные посредники (внутриклеточные медиаторы, мессенджеры) ц. АМФ циклический аденозинмонофосфат (аденозин + рибоза + фосфат) ц. ГМФ циклический гуанозинмонофосфат (гаунозин + рибоза + фосфат) NO оксид азота; Эйказоноиды, т. е. содержащие 20 углеродных атомов простагландины типа Е 2 и F 2 альфа, простациклины, тромбоксаны и лейкотриены
 Диацилглицерин (ДАГ) Инозитолтрифосфат (инозиттрифосфат, ИТФ) Лизофосфатидилхолин (ЛФХ) Церамид Сфингозин") Вторичные посредники (внутриклеточные медиаторы, месенджеры) Диацилглицерин (ДАГ) Инозитолтрифосфат (инозиттрифосфат, ИТФ) Лизофосфатидилхолин (ЛФХ) Церамид Сфингозин 1 фосфат Ras белок (активируется протеинкиназой С) Белок р53
Вторичные посредники (внутриклеточные медиаторы, месенджеры) Диацилглицерин (ДАГ) Инозитолтрифосфат (инозиттрифосфат, ИТФ) Лизофосфатидилхолин (ЛФХ) Церамид Сфингозин 1 фосфат Ras белок (активируется протеинкиназой С) Белок р53
 ГТФ (→ ц. ГМФ) Аргинин (→") Предшественники синтеза вторичных посредников АТФ (→ ц. АМФ) ГТФ (→ ц. ГМФ) Аргинин (→ NO) Фосфатидилинозитдифосфат (→ ДАГ и ИТФ) Фосфолипиды (→ арахидоновая кислота)
Предшественники синтеза вторичных посредников АТФ (→ ц. АМФ) ГТФ (→ ц. ГМФ) Аргинин (→ NO) Фосфатидилинозитдифосфат (→ ДАГ и ИТФ) Фосфолипиды (→ арахидоновая кислота)
 Фосфатидилхолин") Предшественники синтеза вторичных посредников Арахидоновая кислота (→ эйказоноиды: простагландины, простациклины , тромобоксаны, лейкотриены) Фосфатидилхолин (→ лизофосфатидилхолин) Сфинголипиды (сфингомиелин) (→ церамид, сфингозин 1 фосфат)
Предшественники синтеза вторичных посредников Арахидоновая кислота (→ эйказоноиды: простагландины, простациклины , тромобоксаны, лейкотриены) Фосфатидилхолин (→ лизофосфатидилхолин) Сфинголипиды (сфингомиелин) (→ церамид, сфингозин 1 фосфат)
 Ключевые ферменты, участвующие в образовании вторичных посредников Аденилатциклаза, участвует в синтезе ц. АМФ Гуанилатциклаза – мембраносвязанная (м. Г) и цитоплазматическая, или растворимая ( р. Г), участвует в синтезе ц. ГМФ NO синтаза, участвует в синтезе NO Фосфолипаза А 2 , участвует в синтезе арахидоновой кислоты и лизофосфатидилхолина
Ключевые ферменты, участвующие в образовании вторичных посредников Аденилатциклаза, участвует в синтезе ц. АМФ Гуанилатциклаза – мембраносвязанная (м. Г) и цитоплазматическая, или растворимая ( р. Г), участвует в синтезе ц. ГМФ NO синтаза, участвует в синтезе NO Фосфолипаза А 2 , участвует в синтезе арахидоновой кислоты и лизофосфатидилхолина
, участвует в синтезе простагландинов, простациклинов") Ключевые ферменты, участвующие в образовании вторичных посредников Циклооксигеназа (ЦОГ), участвует в синтезе простагландинов, простациклинов и тромбоксанов из арахидоновой кислоты Липооксигеназа (ЛОГ), участвует в синтезе лейкотриенов из арахидоновой кислоты
Ключевые ферменты, участвующие в образовании вторичных посредников Циклооксигеназа (ЦОГ), участвует в синтезе простагландинов, простациклинов и тромбоксанов из арахидоновой кислоты Липооксигеназа (ЛОГ), участвует в синтезе лейкотриенов из арахидоновой кислоты
 Ключевые ферменты, участвующие в образовании вторичных посредников Фосфолипаза С, в том числе β изоформа (активируется Gp белком) и γ изоформа (активируется тирозинкиназой, входящей в структуру ряда рецепторов), участвует в синтезе диацилглицерина (ДАГ) и инозитолтрифосфата (ИТФ) из фосфатидилинозитдифосфата (ФИД)
Ключевые ферменты, участвующие в образовании вторичных посредников Фосфолипаза С, в том числе β изоформа (активируется Gp белком) и γ изоформа (активируется тирозинкиназой, входящей в структуру ряда рецепторов), участвует в синтезе диацилглицерина (ДАГ) и инозитолтрифосфата (ИТФ) из фосфатидилинозитдифосфата (ФИД)
 Ключевые ферменты, участвующие в образовании вторичных посредников Сфингомиелиназа, участвует в синтезе церамида из сфингомиелина Церамидаза, участвует в синтезе сфингозина из церамида; Сфингозинкиназа, участвует в синтезе сфингозин 1 фосфата из сфингозина
Ключевые ферменты, участвующие в образовании вторичных посредников Сфингомиелиназа, участвует в синтезе церамида из сфингомиелина Церамидаза, участвует в синтезе сфингозина из церамида; Сфингозинкиназа, участвует в синтезе сфингозин 1 фосфата из сфингозина
 Фосфодиэстераза (для разрушения ц. ГМФ)") Ферменты, разрушающие посредники вторичные Фосфодиэстераза (для разрушения ц. АМФ) Фосфодиэстераза (для разрушения ц. ГМФ)
Ферменты, разрушающие посредники вторичные Фосфодиэстераза (для разрушения ц. АМФ) Фосфодиэстераза (для разрушения ц. ГМФ)
 ц. АМФ зависимая протеинкиназа,") Протеинкиназы – ферменты, фосфорилирующие внутриклеточные эффекторы Протеинкиназа А (ПК А) ц. АМФ зависимая протеинкиназа, активируется 4 молекулами ц. АМФ, фосфорилирует белки по остаткам серина или треонина Протеинкиназа G (ПК G) ц. ГМФ зависимая протеинкиназа, активируется 4 молекулами ц. ГМФ, фосфорилирует белки по остаткам серина или треонина
Протеинкиназы – ферменты, фосфорилирующие внутриклеточные эффекторы Протеинкиназа А (ПК А) ц. АМФ зависимая протеинкиназа, активируется 4 молекулами ц. АМФ, фосфорилирует белки по остаткам серина или треонина Протеинкиназа G (ПК G) ц. ГМФ зависимая протеинкиназа, активируется 4 молекулами ц. ГМФ, фосфорилирует белки по остаткам серина или треонина
 – диацилглицерин (ДАГ) зависимая") Протеинкиназы – ферменты, фосфорилирующие внутриклеточные эффекторы Протеинкиназа С (ПК С) – диацилглицерин (ДАГ) зависимая протеинкиназа, т. е. активируется ДАГ, фосфорилирует белки по остаткам серина или треонина Тирозинкиназа компонент ряда рецепторов или самостоятельный фермент, фосфорилирует белки по остаткам тирозина
Протеинкиназы – ферменты, фосфорилирующие внутриклеточные эффекторы Протеинкиназа С (ПК С) – диацилглицерин (ДАГ) зависимая протеинкиназа, т. е. активируется ДАГ, фосфорилирует белки по остаткам серина или треонина Тирозинкиназа компонент ряда рецепторов или самостоятельный фермент, фосфорилирует белки по остаткам тирозина
, в том числе Raf, МЕКК,") Протеинкиназы – ферменты, фосфорилирующие внутриклеточные эффекторы Митогенактивируемые протеинкиназы (МАПК), в том числе Raf, МЕКК, ERK, РAK, JNK активируются Ras белком, фосфорилируют ядерные транскрипционные факторы, которые контролируют активность генов клеточного цикла Циклинзависимые киназы (Сdks) активируемые белком циклином, фосфорилируют белки, регулирующие фазы клеточного цикла
Протеинкиназы – ферменты, фосфорилирующие внутриклеточные эффекторы Митогенактивируемые протеинкиназы (МАПК), в том числе Raf, МЕКК, ERK, РAK, JNK активируются Ras белком, фосфорилируют ядерные транскрипционные факторы, которые контролируют активность генов клеточного цикла Циклинзависимые киназы (Сdks) активируемые белком циклином, фосфорилируют белки, регулирующие фазы клеточного цикла
 Ионные насосы (натрий калиевый, кальциевые) Киназа легкой") Внутриклеточные эффекторы Ионные каналы (натриевые, кальциевые, хлорные) Ионные насосы (натрий калиевый, кальциевые) Киназа легкой цепи миозина (КЛЦМ) Ключевые ферменты метаболизма Структурные белки Факторы транскрипции Факторы трансляции Энхансеры
Внутриклеточные эффекторы Ионные каналы (натриевые, кальциевые, хлорные) Ионные насосы (натрий калиевый, кальциевые) Киназа легкой цепи миозина (КЛЦМ) Ключевые ферменты метаболизма Структурные белки Факторы транскрипции Факторы трансляции Энхансеры
 Протеинфосфатазы фосфодиэстраза") Внутриклеточные терминаторы эффектов гормонов и медиаторов (дефосфорилируют внутриклеточные эффекторы) Протеинфосфатазы фосфодиэстраза
Внутриклеточные терминаторы эффектов гормонов и медиаторов (дефосфорилируют внутриклеточные эффекторы) Протеинфосфатазы фосфодиэстраза
 Примеры каскада реакций при активации метаботропных рецепторов постсинаптической мембраны
Примеры каскада реакций при активации метаботропных рецепторов постсинаптической мембраны
 + рецептор ↓ 2. Активация Gs") ц. АМФ –опосредованные пути передачи сигнала. Гормон (медиатор) + рецептор ↓ 2. Активация Gs белка или G белка (ГДФ+α+ βγ субъединицы) за счет замены ГДФ α– субъединицы на ГТФ ↓ 3. Распад G белка на ГТФ+α–субъединицу и βγ субъединицы ↓ 4. Латеральная диффузия ГТФ+α–субъединицы в мембране и встреча с аденилатциклазой ↓
ц. АМФ –опосредованные пути передачи сигнала. Гормон (медиатор) + рецептор ↓ 2. Активация Gs белка или G белка (ГДФ+α+ βγ субъединицы) за счет замены ГДФ α– субъединицы на ГТФ ↓ 3. Распад G белка на ГТФ+α–субъединицу и βγ субъединицы ↓ 4. Латеральная диффузия ГТФ+α–субъединицы в мембране и встреча с аденилатциклазой ↓
 ц. АМФ –опосредованные пути передачи сигнала 5. Изменение активности аденилатциклазы под влиянием α–субъединицы (активация с участием αs–субъединицы или ингибирование с участием αi–субъединицы) и изменение внутриклеточного содержания ц. АМФ ↓ 6. α–субъединица приобретает ГТФ азную активность 7. Замена α–субъединице ГТФ на ГДФ (за счет гидролиза ГТФ), в результате чего происходит образование комплекса «ГДФ+α–субъединица»
ц. АМФ –опосредованные пути передачи сигнала 5. Изменение активности аденилатциклазы под влиянием α–субъединицы (активация с участием αs–субъединицы или ингибирование с участием αi–субъединицы) и изменение внутриклеточного содержания ц. АМФ ↓ 6. α–субъединица приобретает ГТФ азную активность 7. Замена α–субъединице ГТФ на ГДФ (за счет гидролиза ГТФ), в результате чего происходит образование комплекса «ГДФ+α–субъединица»
 ц. АМФ –опосредованные пути передачи сигнала 8. Отсоединение комплекса «ГДФ+α– субъединица» от аденилатциклазы и восстановление ее активности до первоначального уровня ↓ 9. Воссоединение комплекса «ГДФ+α– субъединица» с βγ субъединицами, т. е. восстановление целостности G белка ↓ 10. Возвращение G белка в первоначальное состояние, восстановление готовности для проведения нового сигнала от рецептора внутрь клетки
ц. АМФ –опосредованные пути передачи сигнала 8. Отсоединение комплекса «ГДФ+α– субъединица» от аденилатциклазы и восстановление ее активности до первоначального уровня ↓ 9. Воссоединение комплекса «ГДФ+α– субъединица» с βγ субъединицами, т. е. восстановление целостности G белка ↓ 10. Возвращение G белка в первоначальное состояние, восстановление готовности для проведения нового сигнала от рецептора внутрь клетки
, в частности, его") ц. АМФ –опосредованные пути передачи сигнала Примечание: белок возбудителя холеры (холеротоксин), в частности, его субъединица А проникает внутрь клетки и катализирует перенос АДФ рибозы от НАД+ на Gs белок и тем самым лишает возможность α–субъединицу проявлять ГТФ азную активность. Поэтому α– субъединица не отсоединяется от аденилатциклазы и продолжает ее активировать, т. е. нарушается терминация передачи сигнала от рецептора внутрь клетки.
ц. АМФ –опосредованные пути передачи сигнала Примечание: белок возбудителя холеры (холеротоксин), в частности, его субъединица А проникает внутрь клетки и катализирует перенос АДФ рибозы от НАД+ на Gs белок и тем самым лишает возможность α–субъединицу проявлять ГТФ азную активность. Поэтому α– субъединица не отсоединяется от аденилатциклазы и продолжает ее активировать, т. е. нарушается терминация передачи сигнала от рецептора внутрь клетки.
 ц. АМФ –опосредованные пути передачи сигнала В эпителиоцитах кишечника это приводит к резкой активации Na К насоса (он, вероятно, активируется серотонином по Gs аденилатциклазному пути) и к потере Na и воды. Подобным блокирующим Gs белок действием обладает и дифтерийный токсин, но это приводит к длительной активации фактора трансляции.
ц. АМФ –опосредованные пути передачи сигнала В эпителиоцитах кишечника это приводит к резкой активации Na К насоса (он, вероятно, активируется серотонином по Gs аденилатциклазному пути) и к потере Na и воды. Подобным блокирующим Gs белок действием обладает и дифтерийный токсин, но это приводит к длительной активации фактора трансляции.
 Путь «ц. АМФ – внутриклеточный эффектор» 1. Повышение внутриклеточной концентрации ц. АМФ ↓ 2. Активация ц. АМФ зависимой протеинкиназы (ПК А) ↓ 3. Фосфорилирование клеточного эффектора или второй протеинкиназы путем присоединения к белку эффектору фосфатной группы к остаткам серина или (реже) треонина (но не тирозина; это делают тирозинкиназы при передачи сигнала по другим путям) ↓ 4. Внутриклеточный эффект (изменение состояния клетки) ↓
Путь «ц. АМФ – внутриклеточный эффектор» 1. Повышение внутриклеточной концентрации ц. АМФ ↓ 2. Активация ц. АМФ зависимой протеинкиназы (ПК А) ↓ 3. Фосфорилирование клеточного эффектора или второй протеинкиназы путем присоединения к белку эффектору фосфатной группы к остаткам серина или (реже) треонина (но не тирозина; это делают тирозинкиназы при передачи сигнала по другим путям) ↓ 4. Внутриклеточный эффект (изменение состояния клетки) ↓
 Путь «ц. АМФ – внутриклеточный эффектор» 5. Активация фосфодиэстеразы под влияниям ц. АМФ ↓ 6. Превращение ц. АМФ в АМФ (разрыв двойной связи фосфатной группы с рибозой) под влиянием фосфодиэстеразы ↓ 7. Снижение активности протеинкиназы А ↓ 8. Активация протеинфосфатазы
Путь «ц. АМФ – внутриклеточный эффектор» 5. Активация фосфодиэстеразы под влияниям ц. АМФ ↓ 6. Превращение ц. АМФ в АМФ (разрыв двойной связи фосфатной группы с рибозой) под влиянием фосфодиэстеразы ↓ 7. Снижение активности протеинкиназы А ↓ 8. Активация протеинфосфатазы
 Путь «ц. АМФ – внутриклеточный эффектор» 9. Дефосфорилирование клеточного эффектора или второй протеинкиназы ↓ 10. Терминация действия гормона или медиатора (завершение вутриклеточного эффекта, т. е. возврат клетки к исходному функциональному состоянию)
Путь «ц. АМФ – внутриклеточный эффектор» 9. Дефосфорилирование клеточного эффектора или второй протеинкиназы ↓ 10. Терминация действия гормона или медиатора (завершение вутриклеточного эффекта, т. е. возврат клетки к исходному функциональному состоянию)
 Пример участия адреналина в активации гликогенолиза в гепатоцитах 1. Адреналин + β 2 адренорецептор ↓ 2. Активация Gs белка ↓ 3. Активация аденилатциклазы ↓ 4. Повышение внутриклеточной концентрации ц. АМФ ↓ 5. Активация протеинкиназы А под влиянием ц. АМФ ↓
Пример участия адреналина в активации гликогенолиза в гепатоцитах 1. Адреналин + β 2 адренорецептор ↓ 2. Активация Gs белка ↓ 3. Активация аденилатциклазы ↓ 4. Повышение внутриклеточной концентрации ц. АМФ ↓ 5. Активация протеинкиназы А под влиянием ц. АМФ ↓
 Пример участия адреналина в активации гликогенолиза в гепатоцитах 6. Активация киназы фосфорилазы b (КФ b) и ингибирование гликогенсинтетазы под влиянием протеинкиназы А ↓ 7. Активация гликогенфосфорилазы b (ГФ b) под влиянием фосфорилазы b ↓ 8. Расщепление гликогена до глюкозо 1 фосфата ↓ 9. Превращение глюкозо 1 фосфата в глюкозо 6 фосфат и его использование на нужды клетки
Пример участия адреналина в активации гликогенолиза в гепатоцитах 6. Активация киназы фосфорилазы b (КФ b) и ингибирование гликогенсинтетазы под влиянием протеинкиназы А ↓ 7. Активация гликогенфосфорилазы b (ГФ b) под влиянием фосфорилазы b ↓ 8. Расщепление гликогена до глюкозо 1 фосфата ↓ 9. Превращение глюкозо 1 фосфата в глюкозо 6 фосфат и его использование на нужды клетки
 в адипоцитах. Адреналин + β") Пример участия адреналина в активации распада жира ( липолиза) в адипоцитах. Адреналин + β 2 адренорецептор ↓ 2. Активация Gs белка ↓ 3. Активация аденилатциклазы ↓ 4. Повышение внутриклеточной концентрации ц. АМФ ↓ 5. Активация протеинкиназы А под влиянием ц. АМФ ↓ 6. Активация триглицеридлипазы (ТГЛ) под влиянием протеинкиназы А ↓ 7. Распад нейтральных жиров (липолиз) до жирных кислот и глицерина
Пример участия адреналина в активации распада жира ( липолиза) в адипоцитах. Адреналин + β 2 адренорецептор ↓ 2. Активация Gs белка ↓ 3. Активация аденилатциклазы ↓ 4. Повышение внутриклеточной концентрации ц. АМФ ↓ 5. Активация протеинкиназы А под влиянием ц. АМФ ↓ 6. Активация триглицеридлипазы (ТГЛ) под влиянием протеинкиназы А ↓ 7. Распад нейтральных жиров (липолиз) до жирных кислот и глицерина
 Пример участия адреналина в ингибировании сокращений гладких мышц 1. Адреналин + β 2 адренорецептор ↓ 2. Активация Gs белка ↓ 3. Активация аденилатциклазы ↓ 4. Повышение внутриклеточной концентрации ц. АМФ ↓ 5. Активация протеинкиназы А под влиянием ц. АМФ ↓ 6. Ингибирование киназы легких цепей миозина (КЛЦМ) путем ее фосфорилирования под влиянием протеинкиназы А ↓ 7. Прекращение взаимодействия головок миозина с активноыми нитями ↓ 8. Расслабление гладкой мышечной клетки
Пример участия адреналина в ингибировании сокращений гладких мышц 1. Адреналин + β 2 адренорецептор ↓ 2. Активация Gs белка ↓ 3. Активация аденилатциклазы ↓ 4. Повышение внутриклеточной концентрации ц. АМФ ↓ 5. Активация протеинкиназы А под влиянием ц. АМФ ↓ 6. Ингибирование киназы легких цепей миозина (КЛЦМ) путем ее фосфорилирования под влиянием протеинкиназы А ↓ 7. Прекращение взаимодействия головок миозина с активноыми нитями ↓ 8. Расслабление гладкой мышечной клетки
 Виды рецепторов, при которых передача сигнала идет с участием Gi белка, т. е. при наличии ингибирующей αi субъединицы (снижение ц. АМФ, угнетение активности протеинкиназы А) α 2 – адренорецепторы эндотелия сосудов М 2 холинорецепторы сердца (часть М 2 ХР активируют ц. ГТФ путь) Опиоидные рецепторы нейронов головного мозга, в том числе воспринимающие препараты опия Канабиноидные рецепторы нейронов головного мозга, в том числе воспринимающие препараты конопли
Виды рецепторов, при которых передача сигнала идет с участием Gi белка, т. е. при наличии ингибирующей αi субъединицы (снижение ц. АМФ, угнетение активности протеинкиназы А) α 2 – адренорецепторы эндотелия сосудов М 2 холинорецепторы сердца (часть М 2 ХР активируют ц. ГТФ путь) Опиоидные рецепторы нейронов головного мозга, в том числе воспринимающие препараты опия Канабиноидные рецепторы нейронов головного мозга, в том числе воспринимающие препараты конопли
 и") Пути передачи сигнала от рецепторов внутрь клетки, опосредованные липидами (ДАГ диацилглицерин, ИТФ инозитолтрифосфат) и ионами Са 1. Рецептор + гормон ↓ 2. Активация Gp белка (аналог Gs белка) ↓ 3. Активация β изоформы фосфолипазы С (ФЛ Сβ) под влиянием αр субъединицы Gs белка ↓ 4. Расщепление фосфатидилинозитдифосфата (ФИД) до диацилглицерина (ДАГ) и инозитолтрифосфата (ИТФ) с участием β фосфолипазы С ↓
Пути передачи сигнала от рецепторов внутрь клетки, опосредованные липидами (ДАГ диацилглицерин, ИТФ инозитолтрифосфат) и ионами Са 1. Рецептор + гормон ↓ 2. Активация Gp белка (аналог Gs белка) ↓ 3. Активация β изоформы фосфолипазы С (ФЛ Сβ) под влиянием αр субъединицы Gs белка ↓ 4. Расщепление фосфатидилинозитдифосфата (ФИД) до диацилглицерина (ДАГ) и инозитолтрифосфата (ИТФ) с участием β фосфолипазы С ↓
 и") Пути передачи сигнала от рецепторов внутрь клетки, опосредованные липидами (ДАГ диацилглицерин, ИТФ инозитолтрифосфат) и ионами Са 5 а Активация Са каналов поверхностной мембраны клетки и саркоплазматического ретикулюма под влиянием инозитолтрифосфата (ИТФ) ↓ 6 а. Повышение активности клеток, обусловленное ростом концентрации свободного Са внутри клетки ↓ 5 б Активация мембраносвязанной протеинкиназы С под влиянием диацилглицерина (ДАГ), предназначнной для фосфорилирования многих клеточных белкок и фосфолипазы С ↓ 6 б. Изменение активности клеток, обусловленное фосфорилировнием клеточных эффекторов под влиянием протеинкиназы С ↓ 7. Терминация процессов за счет метаболизма ИТФ и ДАГ.
Пути передачи сигнала от рецепторов внутрь клетки, опосредованные липидами (ДАГ диацилглицерин, ИТФ инозитолтрифосфат) и ионами Са 5 а Активация Са каналов поверхностной мембраны клетки и саркоплазматического ретикулюма под влиянием инозитолтрифосфата (ИТФ) ↓ 6 а. Повышение активности клеток, обусловленное ростом концентрации свободного Са внутри клетки ↓ 5 б Активация мембраносвязанной протеинкиназы С под влиянием диацилглицерина (ДАГ), предназначнной для фосфорилирования многих клеточных белкок и фосфолипазы С ↓ 6 б. Изменение активности клеток, обусловленное фосфорилировнием клеточных эффекторов под влиянием протеинкиназы С ↓ 7. Терминация процессов за счет метаболизма ИТФ и ДАГ.
 Пример использования пути передачи сигнала с участием ИТФ и ДАГ (Активация α 1 АР в миоцитах ряда сосудов) 1. Норадреналин + α 1 АР миоцитов ↓ 2. Активация 1 Gp белка ↓ 3. Активация β изоформы фосфолипазы С (ФЛ Сβ) под влиянием αр субъединицы Gs белка ↓ 4. Расщепление фосфатидилинозитдифосфата (ФИД) до диацилглицерина (ДАГ) и инозитолтрифосфата (ИТФ) с участием β фосфолипазы С ↓
Пример использования пути передачи сигнала с участием ИТФ и ДАГ (Активация α 1 АР в миоцитах ряда сосудов) 1. Норадреналин + α 1 АР миоцитов ↓ 2. Активация 1 Gp белка ↓ 3. Активация β изоформы фосфолипазы С (ФЛ Сβ) под влиянием αр субъединицы Gs белка ↓ 4. Расщепление фосфатидилинозитдифосфата (ФИД) до диацилглицерина (ДАГ) и инозитолтрифосфата (ИТФ) с участием β фосфолипазы С ↓
 Пример использования пути передачи сигнала с участием ИТФ и ДАГ (Активация α 1 АР в миоцитах ряда сосудов) 5 а Активация Са каналов поверхностной мембраны миоцитоа сосудов под влиянием инозитолтрифосфата (ИТФ) ↓ 6 а. Повышение тонуса миоцита, обусловленное ростом внутриклеточной концентрации свободного Са внутри клетки ↓ 5 б Активация мембраносвязанной протеинкиназы С под влиянием диацилглицерина (ДАГ) ↓ 6 б. Фосфорилирование (с участием протеинкиназы С) какого то белка эффектора, способного повысить тонус миоцита ↓ 7. Терминация процессов за счет метаболизма ИТФ и ДАГ.
Пример использования пути передачи сигнала с участием ИТФ и ДАГ (Активация α 1 АР в миоцитах ряда сосудов) 5 а Активация Са каналов поверхностной мембраны миоцитоа сосудов под влиянием инозитолтрифосфата (ИТФ) ↓ 6 а. Повышение тонуса миоцита, обусловленное ростом внутриклеточной концентрации свободного Са внутри клетки ↓ 5 б Активация мембраносвязанной протеинкиназы С под влиянием диацилглицерина (ДАГ) ↓ 6 б. Фосфорилирование (с участием протеинкиназы С) какого то белка эффектора, способного повысить тонус миоцита ↓ 7. Терминация процессов за счет метаболизма ИТФ и ДАГ.
 Медиаторы. Комедиаторы. Модуляторы В роли медиатора выступают десятки и даже сотни химических веществ. Для того чтобы назвать вещество медиатором, используют следующие критерии. 1. Вещество выделяется из клетки при ее активации. 2. В клетке имеются ферменты для синтеза данного вещества. 3. В соседних клетках имеются белки рецепторы, активируемые данным медиатором. 4. Фармакологический (экзогенный) аналог имитирует действие медиатора. Наряду с медиаторами в синапсе есть модуляторы, т. е. вещества, которые прямо не участвуют в процессе передачи сигнала от нейрона к нейрону, но могут этот процесс существенно усиливать или ослаблять.
Медиаторы. Комедиаторы. Модуляторы В роли медиатора выступают десятки и даже сотни химических веществ. Для того чтобы назвать вещество медиатором, используют следующие критерии. 1. Вещество выделяется из клетки при ее активации. 2. В клетке имеются ферменты для синтеза данного вещества. 3. В соседних клетках имеются белки рецепторы, активируемые данным медиатором. 4. Фармакологический (экзогенный) аналог имитирует действие медиатора. Наряду с медиаторами в синапсе есть модуляторы, т. е. вещества, которые прямо не участвуют в процессе передачи сигнала от нейрона к нейрону, но могут этот процесс существенно усиливать или ослаблять.
 Нейропептиды как медиаторы ЦНС. В ЦНС выявлено большое число нейроактивных пептидов. Также как и медиаторы (нейротрансмиттеры), пептиды идентифицируются как нейроактивные вещества в соответствии с указанными выше критериями. Для них характерна следующая локализация в ЦНС. Субстанция Р – головной мозг, окончания афферентных нейронов в ноцицептивной системе. Вазопрессин, окситоцин – задний гипофиз, продолговатый мозг, спинной мозг
Нейропептиды как медиаторы ЦНС. В ЦНС выявлено большое число нейроактивных пептидов. Также как и медиаторы (нейротрансмиттеры), пептиды идентифицируются как нейроактивные вещества в соответствии с указанными выше критериями. Для них характерна следующая локализация в ЦНС. Субстанция Р – головной мозг, окончания афферентных нейронов в ноцицептивной системе. Вазопрессин, окситоцин – задний гипофиз, продолговатый мозг, спинной мозг
 Нейропептиды как медиаторы ЦНС. Кортиколиберин – медиальное возвышение гипоталамуса и другие отделы мозга. Тиреолиберин – гипоталамус, сетчатка. Соматолиберин – гипоталамус. Соматостатин – гипоталамус и другие отделы мозга, желатинозная субстнация, сетчатка. Гонадолиберин – гипотламус, хеморецептивные зоны желудочков мозга, преганглионарные окончания, сетчатка.
Нейропептиды как медиаторы ЦНС. Кортиколиберин – медиальное возвышение гипоталамуса и другие отделы мозга. Тиреолиберин – гипоталамус, сетчатка. Соматолиберин – гипоталамус. Соматостатин – гипоталамус и другие отделы мозга, желатинозная субстнация, сетчатка. Гонадолиберин – гипотламус, хеморецептивные зоны желудочков мозга, преганглионарные окончания, сетчатка.
 Нейропептиды как медиаторы ЦНС. Эндотелин – задний гипофиз, ствол мозга. Энкефалины – желатинозная субстнация, многие другие отделы ЦНС, сетчатка. Эндорфины – гипоталамус, ствол мозга, сетчатка. Холецистокинин – кора головного мозга, гипоталамус, сетчатка. Вазоактивный интестинальный полипептид – ганглионарные холинергические нейроны, некоторые чувствительные нейроны, гипоталамус, кора больших полушарий головного мозга, сетчатка.
Нейропептиды как медиаторы ЦНС. Эндотелин – задний гипофиз, ствол мозга. Энкефалины – желатинозная субстнация, многие другие отделы ЦНС, сетчатка. Эндорфины – гипоталамус, ствол мозга, сетчатка. Холецистокинин – кора головного мозга, гипоталамус, сетчатка. Вазоактивный интестинальный полипептид – ганглионарные холинергические нейроны, некоторые чувствительные нейроны, гипоталамус, кора больших полушарий головного мозга, сетчатка.
 Нейропептиды как медиаторы ЦНС. Нейротензин – гипоталамус, сетчатка. Гастрин – гипоталамус, продолговатый мозг. Глюкагон – гипоталамус, сетчатка Мотилин – нейрогипофиз, кора мозга, мозжечок. Секретин – гипоталамус, обонятельная луковица, ствол мозга, кора мозга, перегородка, гиппокамп, стриатум. Пептид, генетически родственный кальцитонину – окончания первичных афферентов, вкусовой анализатор.
Нейропептиды как медиаторы ЦНС. Нейротензин – гипоталамус, сетчатка. Гастрин – гипоталамус, продолговатый мозг. Глюкагон – гипоталамус, сетчатка Мотилин – нейрогипофиз, кора мозга, мозжечок. Секретин – гипоталамус, обонятельная луковица, ствол мозга, кора мозга, перегородка, гиппокамп, стриатум. Пептид, генетически родственный кальцитонину – окончания первичных афферентов, вкусовой анализатор.
 Нейропептиды как медиаторы ЦНС. Нейропетид Y – симпатические нейроны вегетативной нервной системы. Ангиотензин II – гипоталамус, миндалевидное тело, ствол мозга. Галанин – гипоталамус. Атриопептин – гипоталамус, ствол мозга.
Нейропептиды как медиаторы ЦНС. Нейропетид Y – симпатические нейроны вегетативной нервной системы. Ангиотензин II – гипоталамус, миндалевидное тело, ствол мозга. Галанин – гипоталамус. Атриопептин – гипоталамус, ствол мозга.
 Принцип Дейла. Английский физиолог , Дж. Дейл сформулировал принцип – «Один нейрон – один медиатор» . Дейл полагал, что каждый нейрон выделяет один и тот же медиатор из всех своих нервных окончаний.
Принцип Дейла. Английский физиолог , Дж. Дейл сформулировал принцип – «Один нейрон – один медиатор» . Дейл полагал, что каждый нейрон выделяет один и тот же медиатор из всех своих нервных окончаний.
 Принцип Дейла. Однако в 60 80 е годы 20 века было показано, что выделение многих классических медиаторов (ацетилхолина, норадреналина, ГАМК) сопровождается одновременным высвобождением веществ – комедиаторов, прежде всего – нейропептидов. Примеры: 1. В симпатических нервных окончаниях наряду с норадреналином выделяется нейропетид Y. 2. В парасимпатических окончаниях одновременно с ацетилхолином выделяется вазоактивный интестинальный пептид.
Принцип Дейла. Однако в 60 80 е годы 20 века было показано, что выделение многих классических медиаторов (ацетилхолина, норадреналина, ГАМК) сопровождается одновременным высвобождением веществ – комедиаторов, прежде всего – нейропептидов. Примеры: 1. В симпатических нервных окончаниях наряду с норадреналином выделяется нейропетид Y. 2. В парасимпатических окончаниях одновременно с ацетилхолином выделяется вазоактивный интестинальный пептид.
 Принцип Дейла При этом медиатор обычно оказывает быстрое действие, а комедиатор – медленное или только модулирующее. Поэтому было предложено правило Дейла видоизменить: «Один нейрон – один быстрый медиатор» . Но и это определение потребовало редакции Новая редакция принципа Дейла «Один нейрон – один быстрый синаптический эффект» .
Принцип Дейла При этом медиатор обычно оказывает быстрое действие, а комедиатор – медленное или только модулирующее. Поэтому было предложено правило Дейла видоизменить: «Один нейрон – один быстрый медиатор» . Но и это определение потребовало редакции Новая редакция принципа Дейла «Один нейрон – один быстрый синаптический эффект» .
 Принцип Дейла. В 1999 году были получены данные о том, что в спинном мозге из одного нейрона может выделяться быстрый тормозной медиатор (ГАМК) и быстрый возбуждающий медиатор (АТФ). Таким образом, можно полагать, что число и видов медиаторов, а также комедиаторов и модуляторов в синапсе может быть различным.
Принцип Дейла. В 1999 году были получены данные о том, что в спинном мозге из одного нейрона может выделяться быстрый тормозной медиатор (ГАМК) и быстрый возбуждающий медиатор (АТФ). Таким образом, можно полагать, что число и видов медиаторов, а также комедиаторов и модуляторов в синапсе может быть различным.
 Патология, связанная с обменом медиаторов. Нарушение продукции медиатора – достаточно распространенное явление среди людей. При этом основные симптомы, наблюдаемые в условиях дефицита медиатора, определяются его функцией. Так, при нарушении синтеза ацетилхолина развивается миастения и старческая деменция (болезнь Альцгеймера). Нарушение продукции дофамина вызывает развитие болезни Паркинсона и шизофрении. При нарушении продукции норадреналина и серотонина наблюдаются депрессии, галлюцинации, нарушения сна.
Патология, связанная с обменом медиаторов. Нарушение продукции медиатора – достаточно распространенное явление среди людей. При этом основные симптомы, наблюдаемые в условиях дефицита медиатора, определяются его функцией. Так, при нарушении синтеза ацетилхолина развивается миастения и старческая деменция (болезнь Альцгеймера). Нарушение продукции дофамина вызывает развитие болезни Паркинсона и шизофрении. При нарушении продукции норадреналина и серотонина наблюдаются депрессии, галлюцинации, нарушения сна.
 Патология, связанная с обменом медиаторов Снижение выработки гистамина вызывает вегетативные нарушения, Снижение выработки глутамата вызывает эпилепсию, моторные нарушения, нарушения памяти, дегенеративные нарушения; Снижение выработки глицина вызывает судорожный синдром
Патология, связанная с обменом медиаторов Снижение выработки гистамина вызывает вегетативные нарушения, Снижение выработки глутамата вызывает эпилепсию, моторные нарушения, нарушения памяти, дегенеративные нарушения; Снижение выработки глицина вызывает судорожный синдром
 Патология, связанная с обменом медиаторов Снижение выработки ГАМК вызывает хорею, судорожный синдром, депрессию; Снижение выработки АТФ вызывает нарушение болевой чувствительности, сосудистые расстройства; Снижение выработки аденозина вызывает – судорожные состояния.
Патология, связанная с обменом медиаторов Снижение выработки ГАМК вызывает хорею, судорожный синдром, депрессию; Снижение выработки АТФ вызывает нарушение болевой чувствительности, сосудистые расстройства; Снижение выработки аденозина вызывает – судорожные состояния.
 Свойства химических синапсов. Механизм функционирования химических синапсов обеспечивает реализацию ряда свойств, характерных для ЦНС. 1. Односторонняя проводимость – одно из важнейших свойств химического синапса. Морфологическая и функциональная асимметрия синапса является предпосылкой для суще ствования односторонней проводимости.
Свойства химических синапсов. Механизм функционирования химических синапсов обеспечивает реализацию ряда свойств, характерных для ЦНС. 1. Односторонняя проводимость – одно из важнейших свойств химического синапса. Морфологическая и функциональная асимметрия синапса является предпосылкой для суще ствования односторонней проводимости.
 Свойства химических синапсов. 2. Наличие синаптической задержки. Для того, чтобы в ответ на генерацию ПД в области пресинапса выделился медиатор и произошла генерация ВПСП или ТПСП, требуется определенное время (синаптическая задержка). Она составляет 0, 2 0, 5 мс. В рефлекторных дугах, состоящих из множества нейронов и синаптических связей, это латентное время суммируется и достигает 300 500 мс.
Свойства химических синапсов. 2. Наличие синаптической задержки. Для того, чтобы в ответ на генерацию ПД в области пресинапса выделился медиатор и произошла генерация ВПСП или ТПСП, требуется определенное время (синаптическая задержка). Она составляет 0, 2 0, 5 мс. В рефлекторных дугах, состоящих из множества нейронов и синаптических связей, это латентное время суммируется и достигает 300 500 мс.
,") Свойства химических синапсов. 3. Благодаря синаптическому процессу нервная клетка, управляющая данным постсинаптическим элементом (эффектором), может оказывать возбуждающее воздействие или, наоборот, тормозное. Эффект определяется медиатором
Свойства химических синапсов. 3. Благодаря синаптическому процессу нервная клетка, управляющая данным постсинаптическим элементом (эффектором), может оказывать возбуждающее воздействие или, наоборот, тормозное. Эффект определяется медиатором
 Свойства химических синапсов. 4. В синапсах существует явление отрицательной обратной связи – антидромный эффект. Он состоит в том, что выделяемый в синаптическую щель медиатор может регулиро вать выделение следующей порции медиатора из пресинапса за счет активации рецепторов на пресинаптичекой мембране Свойства химических синапсов. Так, в адренергических синапсах при активации α АР снижается выделение норадреналина при поступлении очередного сигнала к синапсу, а при активации β 2 АР, наоборот, повышается выделение норадреналина
Свойства химических синапсов. 4. В синапсах существует явление отрицательной обратной связи – антидромный эффект. Он состоит в том, что выделяемый в синаптическую щель медиатор может регулиро вать выделение следующей порции медиатора из пресинапса за счет активации рецепторов на пресинаптичекой мембране Свойства химических синапсов. Так, в адренергических синапсах при активации α АР снижается выделение норадреналина при поступлении очередного сигнала к синапсу, а при активации β 2 АР, наоборот, повышается выделение норадреналина
 Свойства химических синапсов 5. Эффективность передачи в синапсе зависит от интервала следования сигналов через синапс. 5 а При уменьшении интервала (до определенной величины) на каждый последующий ПД ответ постсинаптической мембраны (величина ВПСП или ТПСП) будет возрастать (до определенного предела). Это облегчает передачу в синапсе, усиливает ответ постсинаптического элемента на очередной раздражитель. Такое явление называется «облегчение» или «потенциация» . В основе этого явления лежит накопление кальция внутри пресинапса.
Свойства химических синапсов 5. Эффективность передачи в синапсе зависит от интервала следования сигналов через синапс. 5 а При уменьшении интервала (до определенной величины) на каждый последующий ПД ответ постсинаптической мембраны (величина ВПСП или ТПСП) будет возрастать (до определенного предела). Это облегчает передачу в синапсе, усиливает ответ постсинаптического элемента на очередной раздражитель. Такое явление называется «облегчение» или «потенциация» . В основе этого явления лежит накопление кальция внутри пресинапса.
 Свойства химических синапсов 5 б. При выраженном укорочении интервала раздражения происходит избыточное накопление медиатора. Это может приводить к стойкой деполяризации (катодической депрессии), что снижает эффективности синаптической передачи. Это явление называется депрессией.
Свойства химических синапсов 5 б. При выраженном укорочении интервала раздражения происходит избыточное накопление медиатора. Это может приводить к стойкой деполяризации (катодической депрессии), что снижает эффективности синаптической передачи. Это явление называется депрессией.
 Формы синаптической пластичности
Формы синаптической пластичности
 Свойства химических синапсов 6. Если через синапс проходит много импульсов, то в конечном итоге постсинаптическая мембрана может уменьшить ответ на выделение очередной порции медиатора. Это явление называется десенситизацией, т. е. утратой чувствительности к медиатору.
Свойства химических синапсов 6. Если через синапс проходит много импульсов, то в конечном итоге постсинаптическая мембрана может уменьшить ответ на выделение очередной порции медиатора. Это явление называется десенситизацией, т. е. утратой чувствительности к медиатору.
 Свойства химических синапсов 7. Для синапсов характерно утомление, т. е временное снижение эффективности передачи сигнала. В его основе лежат такие процессы как: а) истощение запасов медиатора, б) затруднение выделения медиатора; в) десенситизация постсинаптических рецепторов; Поэтому утомление в синапсах можно рассматривать как интегральный показатель их функционирования.
Свойства химических синапсов 7. Для синапсов характерно утомление, т. е временное снижение эффективности передачи сигнала. В его основе лежат такие процессы как: а) истощение запасов медиатора, б) затруднение выделения медиатора; в) десенситизация постсинаптических рецепторов; Поэтому утомление в синапсах можно рассматривать как интегральный показатель их функционирования.
 Сравнительная продолжительность различных . событий в синапсах Логарифмическая шкала
Сравнительная продолжительность различных . событий в синапсах Логарифмическая шкала
 Частная синаптология продолжение в презентации «Синапс 2»
Частная синаптология продолжение в презентации «Синапс 2»