
СИГНАЛЬНА СИСТЕМА Засоби доставки сигнальних молекул до



















- Размер: 2.9 Mегабайта
- Количество слайдов: 23
Описание презентации СИГНАЛЬНА СИСТЕМА Засоби доставки сигнальних молекул до по слайдам
СИГНАЛЬНА СИСТЕМА
Засоби доставки сигнальних молекул до клітин • ендокринний механізм – гормони діють на клітини відстані від місця синтезу • паракринний механізм –сигнальні молекули активують ближні, сусідні клітини того самого типу. Сигнальні молекули при цьому не потрапляють у кровоток. • аутокринний механізм – клітина відповідає на власний сигнал, тобто має на своїй поверхні рецептори, необхідні для відповіді на секретований ліганд. Аутокринну стимуляцію використовують клітини імунної системи, ауто- та паракринна стимуляція часто відбуваються одночасно. • юкстакринний механізм – задіяний в процесі прикріплення клітин, сигнал передається від однієї клітини до іншої через адгезивні контакти (при запаленнях).
РЕЦЕПТОРИ Існує чотири типи рецепторів та їх сигнальних механізмів: • рецептори, які містять в своїй поліпептидній ланці тирозин-кіназний домен, тобто фактично являються тирозинкіназами (наприклад, рецептори фактору росту); • рецептори , які активуються специфічними цитоплазматичними тирозинкіназами (наприклад , рецептори інтерлейкінів); • рецептори, які містять в своїй структурі серин-треонінкіназний домен (наприклад, родина трансформуючих факторів роста ( TGF — ) ; • інтегринові рецептори (фізично зв ” язують клітину з структурами позаклітинного матриксу). Інформаційні сигнали, що виникають при активації цих рецепторів, приймають участь в механізмах проліферації або диференціації клітин.
Тирозинові протеїнкінази • контролюють фундаментальні клітинні процеси: ріст, диференціація і трансформація, клітинна адгезія, сигнальна трансдукція, трансмембранна сигналізація. • Гіперекспресія генів тирозинових протеїнкіназ активує мутації і викликає ряд патологічних процесів, що призводять до виникнення раку, незлоякісних проліферативних процесів типу атеросклерозу та псоріазу, запальних процесів. • Гіпоекспресія також небезпечна, зменшення активності інсулінової тирозинкінази є причиною різних форм діабету, а клітинної тирозинкінази В – агамаглобулінемії
ФАКТОРИ РОСТУ Фактори з широким діапазоном клітинної і тканинної специфічності : тромбоцитарний фактор росту ( PDGF ); епідермальний фактор росту (EGF); фактори росту фібробластів ( FGF — мають дев’ят ь ізоформ і володіють невеликою клітинною специфічністю). Фактори, специфічні для визначених типів клітин. 1) фактор росту нервів ( NGF ) (діє тільки на клітини нервової системи); 2) еритропоетін (ЕРО) (стимулює утворення еритроцитів в кістковому мозку); 3) інтерлейкін-2( IL 2) та інтерлейкін-3( IL 3) (фактори росту лімфоцитів).
Фосфорилювання білків • контролює клітинний цикл та клітинний метаболізм • може викликати різноманітні клітинні відповіді: ріст та диференціацію, стан мітозу або псевдомітозу, проліферативні процеси та трансформацію клітин
Фосфорилювання виконує дві основні функції в процесі передачі сигналу. • По-перше — змінює конформацію білків і активує ферменти, які можуть проявляти кіназну активність. • По-друге – фосфорилювання, особливо тирозину, створює в молекулах білків «стиковочні» ділянки. З появою таких ділянок в процес залучаються нові білки, які взаємодіють з вже активованими елементами сигнального шляху
7 класів білків контролюють клітинний ріст
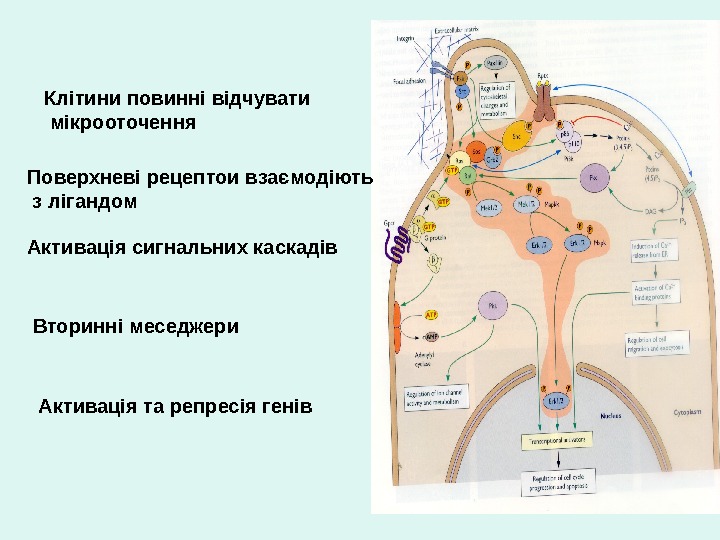
Клітини повинні відчувати мікрооточення Поверхневі рецептои взаємодіють з лігандом Активація сигнальних каскадів Вторинні меседжери Активація та репресія генів
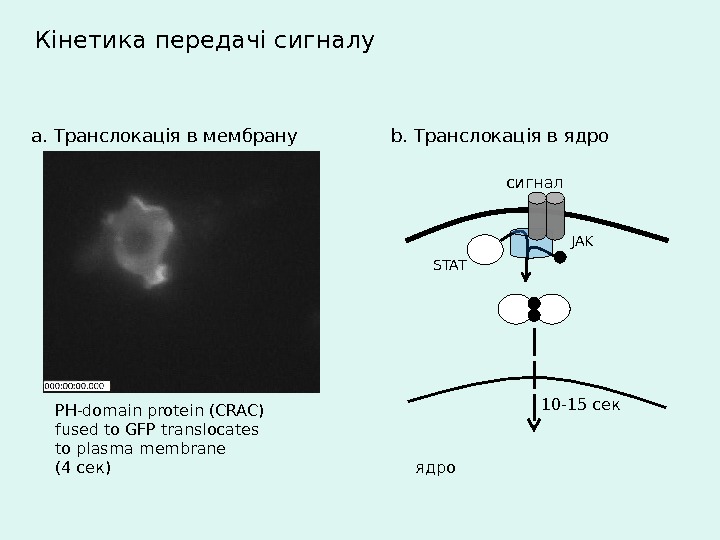
Кінетика передачі сигналу a. Транслокація в мембрану PH-domain protein (CRAC) fused to GFP translocates to plasma membrane (4 сек ) b. Транслокація в ядро 10 -15 сексигнал ядро STAT JAK
SOS –гуанін нуклеотидний фактор SH 2 білки — адапторні бвлки Ras — — важливий G G білок
Трансдукція сигналу Crb 2 Sos Raf Map 2 Mapk GDP GTP TF Активний білок
MAP кінази • Активований Ras індукує сигнальний кіназний каскад який формує та активує MAP кінази (mitogen-activated kinase) • MAP кінази це серин / треонін кінази які можуть : 1. Транслокуватись в ядро та 2. Фосфорилювати транскрипційні фактори що регулюють генну експресію
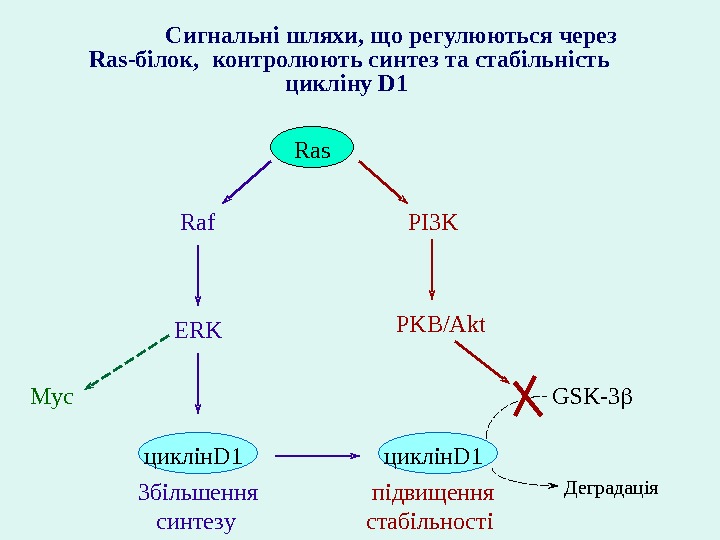
PI 3 KRaf ERK PKB/Akt цикл і н. D 1 Збільшення синтез у підвищення стаб і льност і GSK-3 Деградац і я. Ras Сигнальні шляхи, що регулюються через Ras -білок, контрол ю ют ь синтез та стаб і льн і сть цикл і н у D 1 Myc
Мутац ії онкоген і в Ras-MAP шляхів Перманентна стимуляц і я р о змножен н я (самодостат ні сть у прол і феративн и х сигналах) зміна цитоскелет у /локомоц ії , стимуляц і я м і грац ії ( і нваз і я/метастаз у ван ня ) Без межне р о змножен ня неопластич н их кл і т ин
Мутац ії протоонкоген і в Ras характерн і для пухлин людини Мутац ії в кодонах 12, 13, 61 ген і в K-RAS, N-RAS, H-RAS виявляються у : 60 -80% випадків рак у підшлункової залози ; 25 -40% випадків багатьох інших ново утворень — рак у ле гень , то в стого кишечника, щито подібної залози
Онкоген Myc стимул ює вх і д у S фазу за рахунок декількох механ і зм і в циклин. D/Cdk 4 циклин. Е/Cdk 2 вх і д в S фазу. Rb. E 2 FRb. PMyc Id 2 Cdk 4 циклин. Е ? p 27 Kip 1 деградац і я. Сdc
Шляхи активац ії Myc в неопластич н их кл і т ин ах: 1) Хромосомн і транслокац ії , що перем іщують MYC п і д контроль регуляторн и х е лемент і в ген і в і муноглобул і н і в (л і мфома Б е рк іт та, л і мфосарком и ); 2) Ампл і ф і кац і я та г і пер е кспрес і я ген у (рак молочно ї залози , простат и ); 3) Мутац ії , що п ідвищують стаб і льн і сть б і лк у (л і мфома Б е рк іт та) Зміна структур и /регуляц ії ген у MY
Рак щитовидной железы, MEN 2 ab Рак молочной железы Острый и хрон. миелолейкозы Рак щитовидной железы, эпифиза Опухоли яичника, надпочечника Многие новообразования Рак толстого кишечника Острый лимфолейкоз Лимфома Бэркита, лимфосаркома Опухоли толстого кишечника Саркома Юинга Рак пищевода, печени и др. опухоли Рецепторные тирозинкиназы: мутации, инверсии RET/GDNF-R амплификация ERBB 2/HER 2 транслокации PDGF-R GPCR (G Protein-Coupled Receptors): мутации GSP мутации GIP 2 G-белки: мутации K-RAS, N-RAS, H-RAS Нерецепторные тирозинкиназы: мутации SRC Компоненты STAT-путей: хром. транслокации JAK 2 Факторы транскрипции: транслокации, мутации C-MYC мутации CTNNB 1 (-катенина) хром. транслокации FLI 1 Циклинзависимые киназы: амплификация РRAD 1/циклин. D 1 Новообразования человека, ассоциированные с изменениями протоонкогенов, контролирующих вход в S-фазу
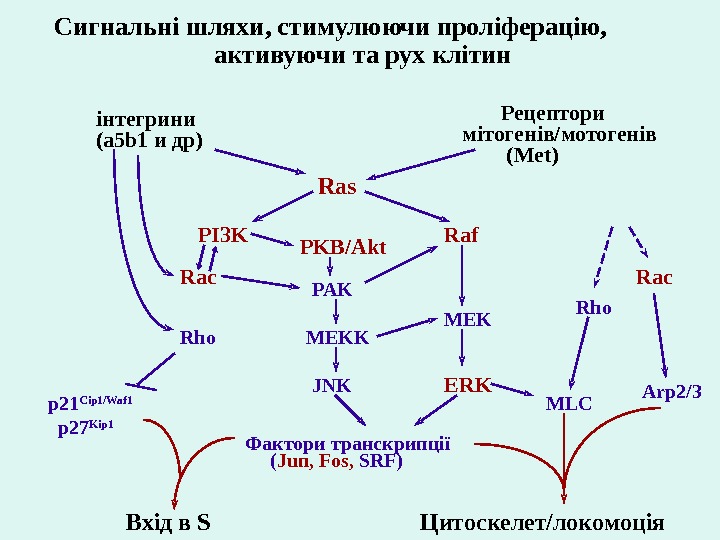
і нтегрин и (a 5 b 1 и др) Рецептор и м і тоген і в/мотоген і в (Met) PI 3 K Raf ERKRho Ras MEKRac PAK MEKK JNK Фактор и транскрипц ії ( Jun, Fos, SRF)p 21 Cip 1/Waf 1 p 27 Kip 1 Вх і д в S Цитоскелет/локомоц і я. Сигнальн і шляхи , стимулю ючи прол і ферац і ю, активую чи та рух кл і т ин PKB/Akt MLC Rac Arp 2/3 Rho
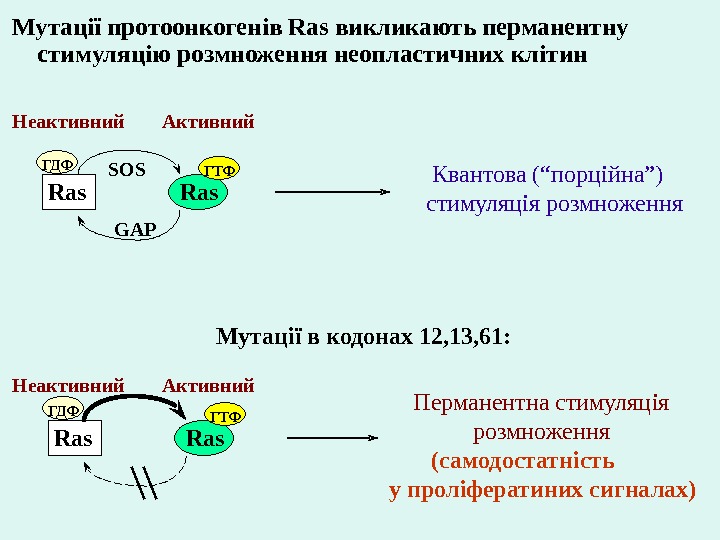
Мутац ії протоонкоген і в Ras в иклика ют ь перманентну стимуляц і ю р о змножен н я неопластич н их кл і т ин Ras. ГДФ ГТФ SOS GAP Ras. ГДФ ГТФ Мутац ії в кодонах 12, 13, 61: Квантова (“порц ій на”) стимуляц і я р о змножен н я Неактивн ий Активн ий Перманентна стимуляц і я р о змножен н я (самодостатн і сть у прол і фератин и х сигналах)Неактивн ий Активн ий
Нечувствительность к рост-супрессирующим сигналам Отсутствие репликативного старения (“иммортализация”) Стимуляция неоангиогенеза Изменения цитоскелета/локомоции (инвазия/метастазирование) Самодостаточность в пролиферативных сигналах Блокирование клеточной дифференцировки Ослабление индукции апоптоза Генетическая нестабильность. Важнейшие свойства неопластической клетки