

Мембрани L.ppt
- Количество слайдов: 47
 Що таке життя? 21. 1
Що таке життя? 21. 1
 Одноклітинні (невидимі") Систематика різномаїття форм живого «Эдем» Ван Кессель 1626 -1679 Багатоклітинні (видно оком) Одноклітинні (невидимі оком)
Систематика різномаїття форм живого «Эдем» Ван Кессель 1626 -1679 Багатоклітинні (видно оком) Одноклітинні (невидимі оком)
 Основні функції живого 1. Гомеостаз здатність відкритої системи зберігати постійність свого внутрішнього стану шляхом скоординованих реакцій, що направлені на підтримку динамічної рівноваги 2. Розмноження (коментарів не потребує)
Основні функції живого 1. Гомеостаз здатність відкритої системи зберігати постійність свого внутрішнього стану шляхом скоординованих реакцій, що направлені на підтримку динамічної рівноваги 2. Розмноження (коментарів не потребує)
 Помилки: мутації та") Основні властивості живого 1. Складна система 2. Компартменталізація 3. Поділ (реплікація) Помилки: мутації та еволюція 4. Обмін речовиною та енергією 5. Функціональність 6. Реакція на середовище
Основні властивості живого 1. Складна система 2. Компартменталізація 3. Поділ (реплікація) Помилки: мутації та еволюція 4. Обмін речовиною та енергією 5. Функціональність 6. Реакція на середовище
 Перше наукове визначення живого сформульовано Ф. Энгельсом: «Життя є спосіб існування білкових тіл, і цей спосіб існування полягає за своєю суттю у постійному оновленні їх хімічних складових частин шляхом прийому їжі та виділення»
Перше наукове визначення живого сформульовано Ф. Энгельсом: «Життя є спосіб існування білкових тіл, і цей спосіб існування полягає за своєю суттю у постійному оновленні їх хімічних складових частин шляхом прийому їжі та виділення»
 Мінімальна одиниця живого клітина
Мінімальна одиниця живого клітина
 Разноманіття форм клітин c – клітини стебла рослини d – сперматозоїди людини e – еритроцити людини f – ембріон людини, 1 поділ
Разноманіття форм клітин c – клітини стебла рослини d – сперматозоїди людини e – еритроцити людини f – ембріон людини, 1 поділ
 Клітина бактерій
Клітина бактерій
 тварин Клітини рослин ядро мітохондрії хлоропласти
тварин Клітини рослин ядро мітохондрії хлоропласти
 Прокаріоти та еукаріоти безядерні і ядровмісні
Прокаріоти та еукаріоти безядерні і ядровмісні
 Рівні структурної організації % 7 15 3
Рівні структурної організації % 7 15 3
 Загальні риси структурної хімії живого Що справедливо для слона – справедливо і для бактерії
Загальні риси структурної хімії живого Що справедливо для слона – справедливо і для бактерії
 Живі організми являють собою відкриті (тобто такі, що обмінюються з оточуючим середовищем речовиною і енергією), самоврегульовані і самовідтворювані системи, у яких найважливішими функціонуючими речовинами є білки та нуклеїнові кислоти. Живому притаманний ряд сукупних рис, таких, як здатність до відтворювання (репродукції), використання та трансформація енергії, метаболізм, чутливість, здатність до змін. Сукупність цих ознак можна знайти вже на клітинному рівні. Нема меншої одиниці живого, ніж клітина.
Живі організми являють собою відкриті (тобто такі, що обмінюються з оточуючим середовищем речовиною і енергією), самоврегульовані і самовідтворювані системи, у яких найважливішими функціонуючими речовинами є білки та нуклеїнові кислоти. Живому притаманний ряд сукупних рис, таких, як здатність до відтворювання (репродукції), використання та трансформація енергії, метаболізм, чутливість, здатність до змін. Сукупність цих ознак можна знайти вже на клітинному рівні. Нема меншої одиниці живого, ніж клітина.
 Клітина – це обмежена активною мембраною, впорядкована, структурована система біополімерів (білків, нуклеїнових кислот та ін. ) і їх молекулярних комплексів, що приймають участь в єдиній сукупності метаболічних і енергетичних процесів, які здійснюють підтримку та відтворюваність усієї системи в цілому. Нема меншої одиниці живого, ніж клітина.
Клітина – це обмежена активною мембраною, впорядкована, структурована система біополімерів (білків, нуклеїнових кислот та ін. ) і їх молекулярних комплексів, що приймають участь в єдиній сукупності метаболічних і енергетичних процесів, які здійснюють підтримку та відтворюваність усієї системи в цілому. Нема меншої одиниці живого, ніж клітина.
 15 Нуклеїнові кислоти 7 Полісахариди 3 Білки Молекули Вода ліпіди органічні та") Полімери (25) 15 Нуклеїнові кислоти 7 Полісахариди 3 Білки Молекули Вода ліпіди органічні та неорганічні (75) 70 2 3
Полімери (25) 15 Нуклеїнові кислоти 7 Полісахариди 3 Білки Молекули Вода ліпіди органічні та неорганічні (75) 70 2 3
 Біологічна мембрана = ліпідний бішар + білки
Біологічна мембрана = ліпідний бішар + білки
 БІОМЕМБРАНИ • Відділяють клітину від зовнішнього середовища і обумовлюють їх індивідуальність. • Бар’єр з високою вибірковою проникністю, що має специфічні молекулярні та іонні помпи та канали та регулює молекулярний та іонний склад внутрішньоклітинного середовища. Бар’єр для токсичних молекул. Допомагає в накопиченні та зберіганні поживних речовин • Регулюють обмін інформацією між клітинами та середовищем, несуть специфічні рецептори, що сприймають зовнішні стимули (рух бактерій до джерела їжі, сприйняття світла та ін. ). • Мембранні системи, що містять високо упорядковані набори ферментів та інших білків є середовищем для процесів перетворення енергії (фотосинтез, окислювальне фосфорилювання) • Полегшує рух клітини • Допомагає при репродукції Мембрани відіграють центральну роль в системі біологічної комунікації
БІОМЕМБРАНИ • Відділяють клітину від зовнішнього середовища і обумовлюють їх індивідуальність. • Бар’єр з високою вибірковою проникністю, що має специфічні молекулярні та іонні помпи та канали та регулює молекулярний та іонний склад внутрішньоклітинного середовища. Бар’єр для токсичних молекул. Допомагає в накопиченні та зберіганні поживних речовин • Регулюють обмін інформацією між клітинами та середовищем, несуть специфічні рецептори, що сприймають зовнішні стимули (рух бактерій до джерела їжі, сприйняття світла та ін. ). • Мембранні системи, що містять високо упорядковані набори ферментів та інших білків є середовищем для процесів перетворення енергії (фотосинтез, окислювальне фосфорилювання) • Полегшує рух клітини • Допомагає при репродукції Мембрани відіграють центральну роль в системі біологічної комунікації
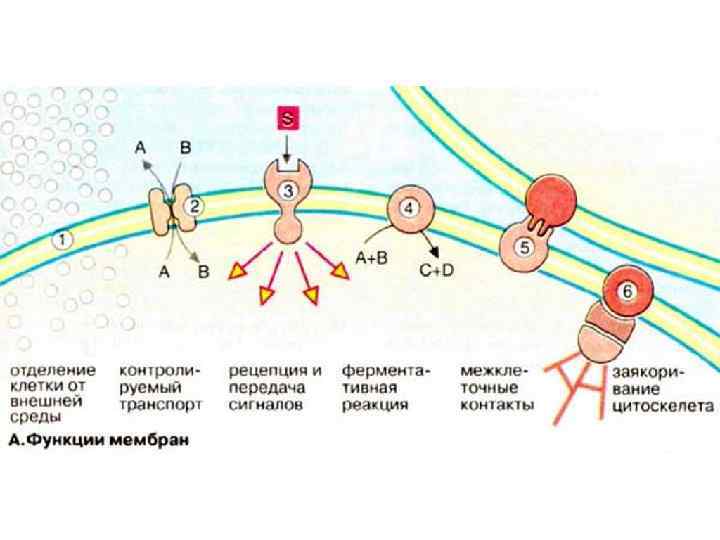
 Загальні властивості БІОМЕМБРАН Мембрани – це площинні структури товщиною у декілька молекул, що утворюють суцільну перегородку між окремими відсіками. Товщина мембран зазвичай 60 -100Å. Мембрани складаються головним чином із ліпідів та білків (співвідношення від 1: 4 до 4: 1). У мембранах є також вуглеводневі компоненти. Ліпіди мембран представлені відносно невеликими молекулами з гідрофільними та гідрофобними групами, які у воді спонтанно утворюють замкнуті бімолекулярні шари, що слугують бар’єром для полярних сполук. За окремі функції мембран є відповідальними окремі білки, що виконують роль помп, каналів, рецепторів, ферментів та перетворювачів енергії. Мембрани – це нековалентні надмолекулярні структури. Білки та ліпіди, що утворюють мембрану, утримуються разом за рахунок виникнення багатьох нековалентних взаємодій, що є кооперативними. Мембрани є асиметричними, їх зовнішня та внутрішня поверхні відрізняються Мембрани – рідкі структури. Молекули ліпідів та білків можуть у багатьох випадках легко дифундувати у площині мембрани.
Загальні властивості БІОМЕМБРАН Мембрани – це площинні структури товщиною у декілька молекул, що утворюють суцільну перегородку між окремими відсіками. Товщина мембран зазвичай 60 -100Å. Мембрани складаються головним чином із ліпідів та білків (співвідношення від 1: 4 до 4: 1). У мембранах є також вуглеводневі компоненти. Ліпіди мембран представлені відносно невеликими молекулами з гідрофільними та гідрофобними групами, які у воді спонтанно утворюють замкнуті бімолекулярні шари, що слугують бар’єром для полярних сполук. За окремі функції мембран є відповідальними окремі білки, що виконують роль помп, каналів, рецепторів, ферментів та перетворювачів енергії. Мембрани – це нековалентні надмолекулярні структури. Білки та ліпіди, що утворюють мембрану, утримуються разом за рахунок виникнення багатьох нековалентних взаємодій, що є кооперативними. Мембрани є асиметричними, їх зовнішня та внутрішня поверхні відрізняються Мембрани – рідкі структури. Молекули ліпідів та білків можуть у багатьох випадках легко дифундувати у площині мембрани.
 ЛІПІДИ
ЛІПІДИ
 Класи ліпідів Усі біологічні ліпіди є амфіфільними: • • Гліцерофосфоліпіди Сфінголіпіди Гліколіпіди Холестерол Полярна “голова” Гідрофобний “хвіст”
Класи ліпідів Усі біологічні ліпіди є амфіфільними: • • Гліцерофосфоліпіди Сфінголіпіди Гліколіпіди Холестерол Полярна “голова” Гідрофобний “хвіст”
 Гліцерофосфоліпіди: основний компонент біомембран
Гліцерофосфоліпіди: основний компонент біомембран
 Структура фосфолипида
Структура фосфолипида
 Жирні кислоти • Карбонові кислоти з довгим, нерозгалуженим ланцюгом • відрізняються по довжині вуглецевого ланцюга, кількості і положенню подвійних зв’язків • Подвійні зв’язки (якщо присутні), як правило, виявляють цис-конфігурацію Насичені – Пальмітінова к-та (16 C) – Стеаринова к-та (18 C) – Арахінова к-та (20 C) Ненасичені – Олеїнова к-та 18: 1(9) – Лінолева к-та 18: 2 (9, 12) – Арахідонова к-та 20: 4 (5, 8, 11, 14)
Жирні кислоти • Карбонові кислоти з довгим, нерозгалуженим ланцюгом • відрізняються по довжині вуглецевого ланцюга, кількості і положенню подвійних зв’язків • Подвійні зв’язки (якщо присутні), як правило, виявляють цис-конфігурацію Насичені – Пальмітінова к-та (16 C) – Стеаринова к-та (18 C) – Арахінова к-та (20 C) Ненасичені – Олеїнова к-та 18: 1(9) – Лінолева к-та 18: 2 (9, 12) – Арахідонова к-та 20: 4 (5, 8, 11, 14)
 • Насичені ланцюги упаковані щільно та утворюють більш жорстко") Структурні наслідки ненасиченості (подвійні зв’язки) • Насичені ланцюги упаковані щільно та утворюють більш жорстко організовані агрегати (мембрани) • Ненасичені ланцюги зігнуті та упаковані у менш організований спосіб, з більшою конформаційною гнучкістю
Структурні наслідки ненасиченості (подвійні зв’язки) • Насичені ланцюги упаковані щільно та утворюють більш жорстко організовані агрегати (мембрани) • Ненасичені ланцюги зігнуті та упаковані у менш організований спосіб, з більшою конформаційною гнучкістю
 Типові фосфогліцероліпіди HO-CH 2 -NH 3+ фосфатидилетаноламін CH 3 HO-CH 2 -NCH 3+ CH 3 фосфатидилхолін R = H: фосфатидова к-та COOHO-CH 2 -CH NH 3+ фосфатидилсерин фосфатидилiнозітол
Типові фосфогліцероліпіди HO-CH 2 -NH 3+ фосфатидилетаноламін CH 3 HO-CH 2 -NCH 3+ CH 3 фосфатидилхолін R = H: фосфатидова к-та COOHO-CH 2 -CH NH 3+ фосфатидилсерин фосфатидилiнозітол
 •") Сфінголіпіди + R-COOH H 3 O+ • Похідні аміноспирту сфінгозину (18 вуглецевих атомів) • Містяться у мембранах нейронів; опосередковують специфічність міжклітинних контактів
Сфінголіпіди + R-COOH H 3 O+ • Похідні аміноспирту сфінгозину (18 вуглецевих атомів) • Містяться у мембранах нейронів; опосередковують специфічність міжклітинних контактів
 СФІНГОЛІПІДИ Сфінгомієлин – єдиний фосфоліпід мембран, що не є похідним гліцеролу Гліколіпіди є похідними сфінгозину. Відмінність гліколіпідів від сфінгомієліну полягає в тому, що у ліпідах присутній не фосфорилхолін, а залишки одного або декількох цукрів Залишок фосфорилхоліну Залишок цукру глюкоза або галактоза
СФІНГОЛІПІДИ Сфінгомієлин – єдиний фосфоліпід мембран, що не є похідним гліцеролу Гліколіпіди є похідними сфінгозину. Відмінність гліколіпідів від сфінгомієліну полягає в тому, що у ліпідах присутній не фосфорилхолін, а залишки одного або декількох цукрів Залишок фосфорилхоліну Залишок цукру глюкоза або галактоза
 Сфінголіпіди ГЛІКОЛІПІДИ Найпростіший гліколіпід – цереброзид, що містить лише один залишок цукру Гангліозиди – більш складні гліколіпіди з розгалуженим ланцюгом із декількох (до семи) залишків цукру
Сфінголіпіди ГЛІКОЛІПІДИ Найпростіший гліколіпід – цереброзид, що містить лише один залишок цукру Гангліозиди – більш складні гліколіпіди з розгалуженим ланцюгом із декількох (до семи) залишків цукру
 • Kлючовий ліпід клітинних мембран • Попередник стероїдних гормонів: Статеві гормони естрадіол тестостерон") Холестерин(ол) • Kлючовий ліпід клітинних мембран • Попередник стероїдних гормонів: Статеві гормони естрадіол тестостерон Mетаболічна регуляція глюкокортикоїди Перетравлення Жовчні кислоти Вагітність прогестерон
Холестерин(ол) • Kлючовий ліпід клітинних мембран • Попередник стероїдних гормонів: Статеві гормони естрадіол тестостерон Mетаболічна регуляція глюкокортикоїди Перетравлення Жовчні кислоти Вагітність прогестерон
 Спонтанне утворення ліпідних структур В міцелах у водному середовищі неполярні хвостики знаходяться в центрі сферичної структури Неполярний розчинник В неполярному розчиннику міцели переорієнтовуються вода
Спонтанне утворення ліпідних структур В міцелах у водному середовищі неполярні хвостики знаходяться в центрі сферичної структури Неполярний розчинник В неполярному розчиннику міцели переорієнтовуються вода
 Деякі ліпідні структури, що утворюються спонтанно: Подвійні шари ліпідів – основа для біологічних мембран одношарові утворення (ліпосоми) Ключовою рушійною силою є гідрофобна взаємодія багатошарові утворення
Деякі ліпідні структури, що утворюються спонтанно: Подвійні шари ліпідів – основа для біологічних мембран одношарові утворення (ліпосоми) Ключовою рушійною силою є гідрофобна взаємодія багатошарові утворення
 Липидная мицелла, бислой, липосома Свойства бислоя зависят от липидного состава: предельные и непредельные кислоты, холестерин
Липидная мицелла, бислой, липосома Свойства бислоя зависят от липидного состава: предельные и непредельные кислоты, холестерин
 Бімолекулярні шари ліпідів та їх властивості • • • У водному середовищі більшість фосфоліпідів та гліколіпідів методом самозбірки (спонтанно) утворюють бімолекулярний шар. Основною рушійною силою спонтанного утворення бішару є гідрофобні взаємодії (збільшення ентропії при вивільненні молекул води), виникнення вандерваальсівських, електростатичних взаємодій та водневих зв’язків між головками та молекулами води. Ліпідні бішари мають тенденцію до збільшення своєї поверхні Ліпідні бішари прагнуть замкнутися на себе так, щоб на кінцях не залишалося досяжних для контакту з водою вуглеводневих ланцюгів (утворюється замкнутий простір – компартмент) Ліпідні бішари можуть самозапечатуватися (самозшиватися)
Бімолекулярні шари ліпідів та їх властивості • • • У водному середовищі більшість фосфоліпідів та гліколіпідів методом самозбірки (спонтанно) утворюють бімолекулярний шар. Основною рушійною силою спонтанного утворення бішару є гідрофобні взаємодії (збільшення ентропії при вивільненні молекул води), виникнення вандерваальсівських, електростатичних взаємодій та водневих зв’язків між головками та молекулами води. Ліпідні бішари мають тенденцію до збільшення своєї поверхні Ліпідні бішари прагнуть замкнутися на себе так, щоб на кінцях не залишалося досяжних для контакту з водою вуглеводневих ланцюгів (утворюється замкнутий простір – компартмент) Ліпідні бішари можуть самозапечатуватися (самозшиватися)
 Проникність ліпідних двохшарових мембран K Na+ 10 -14 10 -12 Триптофан + Глюкоза Cl- 10 -10 10 -8 Коефіцієнт проникності, см/с Збільшення проникності Індол Сечовина Гліцерол 10 -6 H 2 O 10 -4 10 -2
Проникність ліпідних двохшарових мембран K Na+ 10 -14 10 -12 Триптофан + Глюкоза Cl- 10 -10 10 -8 Коефіцієнт проникності, см/с Збільшення проникності Індол Сечовина Гліцерол 10 -6 H 2 O 10 -4 10 -2
 Біологічна мембрана = ліпідний бішар + білки Мембранні ліпіди створюють бар’єри проникності, а специфічні білки виконують такі функції мембран як транспорт, передача інформації та перетворення енергії Мембрани розрізняються за вмістом білка, наприклад: -Ізолююча оболонка нервових волокон – біля 18% -Плазматичні мембрани, що містять різноманітні помпи, канали, рецептори, ферменти – біля 50% -внутрішні мембрани мітохондрій та хлоропластів (перетворення енергії) – біля 75% Мембрани, що виконують різні функції відрізняються також за білковим складом.
Біологічна мембрана = ліпідний бішар + білки Мембранні ліпіди створюють бар’єри проникності, а специфічні білки виконують такі функції мембран як транспорт, передача інформації та перетворення енергії Мембрани розрізняються за вмістом білка, наприклад: -Ізолююча оболонка нервових волокон – біля 18% -Плазматичні мембрани, що містять різноманітні помпи, канали, рецептори, ферменти – біля 50% -внутрішні мембрани мітохондрій та хлоропластів (перетворення енергії) – біля 75% Мембрани, що виконують різні функції відрізняються також за білковим складом.
 Вуглеводи в мембранах У кліткових мембранах еукаріот міститься від 2% до 10% вуглеводів у формі гліколіпідів та глікопротеїнів Гліколіпіди представлені похідними сфінгозину з одним і більше залишками цукру У глікопротеїнах один або декілька вуглеводних ланцюгів приєднані до бокових ланцюгів серину, треоніну або аспарагіну. В усіх досліджених клітинах ссавців залишки цукрів в плазматичних мембранах локалізовані лише на зовнішній стороні мембрани. Вуглеводні групи слугують для -Орієнтування глікопротеїнів у мембрані -Підтримання асиметрії біологічних мембран -Міжклітинного впізнавання (вуглеводи мають потенційні можливості для створення дуже великого структурного різноманіття)
Вуглеводи в мембранах У кліткових мембранах еукаріот міститься від 2% до 10% вуглеводів у формі гліколіпідів та глікопротеїнів Гліколіпіди представлені похідними сфінгозину з одним і більше залишками цукру У глікопротеїнах один або декілька вуглеводних ланцюгів приєднані до бокових ланцюгів серину, треоніну або аспарагіну. В усіх досліджених клітинах ссавців залишки цукрів в плазматичних мембранах локалізовані лише на зовнішній стороні мембрани. Вуглеводні групи слугують для -Орієнтування глікопротеїнів у мембрані -Підтримання асиметрії біологічних мембран -Міжклітинного впізнавання (вуглеводи мають потенційні можливості для створення дуже великого структурного різноманіття)
 • Функції мембранних протеїнів залежать від їх орієнтації всередині мембрани") Поперечна асиметрія мембран (транссиметрія) • Функції мембранних протеїнів залежать від їх орієнтації всередині мембрани • Мембранні протеїни перетинають ліпідний шар мембрани не у випадкових, а в певних місцях, тобто вони мають специфічну топологію. Глікофорин Ззовні Всередині
Поперечна асиметрія мембран (транссиметрія) • Функції мембранних протеїнів залежать від їх орієнтації всередині мембрани • Мембранні протеїни перетинають ліпідний шар мембрани не у випадкових, а в певних місцях, тобто вони мають специфічну топологію. Глікофорин Ззовні Всередині
 Поперечна асиметрія ліпідів В більшості мембран клітин ліпідний склад зовнішнього та внутрішнього моношарів неоднаковий: фосфатидилхолін фосфатидилетаноламін фосфатидилсерин сфингоміелін Всього: Процентне співвідношення ліпідів
Поперечна асиметрія ліпідів В більшості мембран клітин ліпідний склад зовнішнього та внутрішнього моношарів неоднаковий: фосфатидилхолін фосфатидилетаноламін фосфатидилсерин сфингоміелін Всього: Процентне співвідношення ліпідів
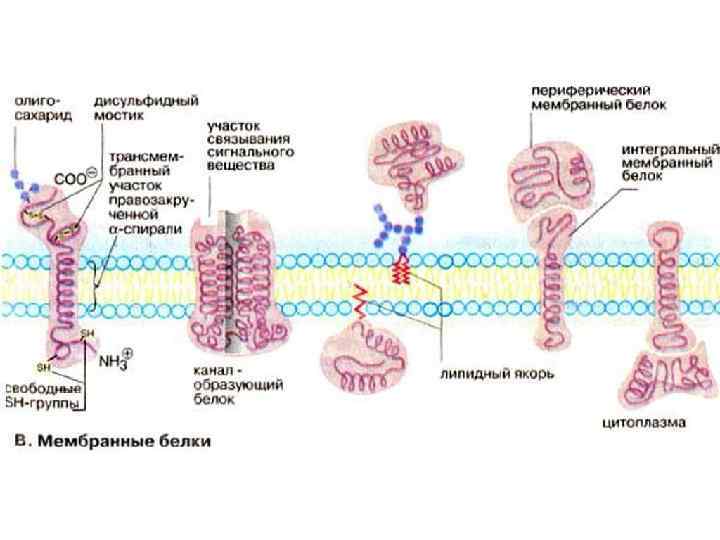
 протеїни • Периферичні (зовнішньо асоційовані) протеїни •") Структура мембранних протеїнів • Інтегральні (внутрішньо асоційовані) протеїни • Периферичні (зовнішньо асоційовані) протеїни • Ліпід-якірні протеїни
Структура мембранних протеїнів • Інтегральні (внутрішньо асоційовані) протеїни • Периферичні (зовнішньо асоційовані) протеїни • Ліпід-якірні протеїни
 Периферичні мембранні протеїни • Периферичні протеїни не щільно прилягають до мембрани • Вони можуть дисоціювати під дією м’якої детергентної обробки або при високих концентраціях солей
Периферичні мембранні протеїни • Периферичні протеїни не щільно прилягають до мембрани • Вони можуть дисоціювати під дією м’якої детергентної обробки або при високих концентраціях солей
 Інтегральні мембранні протеїни • Інтегральні протеїни щільно вбудовані в ліпідний подвійний шар • Вони можуть бути видалені лише при розриві мембрани (очищення) • Часто трансмембранні
Інтегральні мембранні протеїни • Інтегральні протеїни щільно вбудовані в ліпідний подвійний шар • Вони можуть бути видалені лише при розриві мембрани (очищення) • Часто трансмембранні
 Структурна: адгезія протеїнів • Сигнальна: опосередковує ріст") Роль ІМП: • Ідентифікація типу клітини (‘face’) Структурна: адгезія протеїнів • Сигнальна: опосередковує ріст та диференціацію клітин • Система насосів та каналів: контроль імпорту та експорту
Роль ІМП: • Ідентифікація типу клітини (‘face’) Структурна: адгезія протеїнів • Сигнальна: опосередковує ріст та диференціацію клітин • Система насосів та каналів: контроль імпорту та експорту
 Ліпід-якірні протеїни Відносно новий клас мембранних протеїнів Відомі чотири типи: – Амід-зв’язані меристильні якорі – Тіоетер-зв’язані жирні ацильні якорі – Тіоетер-зв’язані фенільні якорі – Глікозил фосфатидилинозитольні якорі Нерідко приймають участь у процесах передачі сигналів
Ліпід-якірні протеїни Відносно новий клас мембранних протеїнів Відомі чотири типи: – Амід-зв’язані меристильні якорі – Тіоетер-зв’язані жирні ацильні якорі – Тіоетер-зв’язані фенільні якорі – Глікозил фосфатидилинозитольні якорі Нерідко приймають участь у процесах передачі сигналів
 • • Рідинно-мозаічна модель Сінгера та Ніколсона Подвійні шари фосфоліпідів та гліколіпідів є рідкою матрицею з бар’єрними характеристиками Два класи протеїнів: – Периферичні протеїни (зовнішньо асоційовані протеїни) – Інтегральні протеїни (внутрішньо асоційовані протеїни) Подвійний шар є двовимірним розчинником для інтегральних білків Ліпіди та протеїни можуть здійснювати обертальний рух та рух у площині, але не можуть переміщуватися у поперечному напрямку
• • Рідинно-мозаічна модель Сінгера та Ніколсона Подвійні шари фосфоліпідів та гліколіпідів є рідкою матрицею з бар’єрними характеристиками Два класи протеїнів: – Периферичні протеїни (зовнішньо асоційовані протеїни) – Інтегральні протеїни (внутрішньо асоційовані протеїни) Подвійний шар є двовимірним розчинником для інтегральних білків Ліпіди та протеїни можуть здійснювати обертальний рух та рух у площині, але не можуть переміщуватися у поперечному напрямку
 МЕМБРАННИЙ ТРАНСПОРТ
МЕМБРАННИЙ ТРАНСПОРТ