

Сенсорные системы.ppt
- Количество слайдов: 99
 Сенсорные системы Физиология анализаторов
Сенсорные системы Физиология анализаторов
 Часть нервной системы, состоящей из воспринимающих элементов") Сенсорной системой (анализатором, по И. П. Павлову) Часть нервной системы, состоящей из воспринимающих элементов — сенсорных рецепторов, получающих стимулы из внешней или внутренней среды, нервных путей, передающих информацию от рецепторов в мозг, и тех частей мозга, которые перерабатывают эту информацию
Сенсорной системой (анализатором, по И. П. Павлову) Часть нервной системы, состоящей из воспринимающих элементов — сенсорных рецепторов, получающих стимулы из внешней или внутренней среды, нервных путей, передающих информацию от рецепторов в мозг, и тех частей мозга, которые перерабатывают эту информацию
 и дендритов") Понятие рецептора Реце птор — сложное образование, состоящие из терминалей (нервных окончаний) и дендритов чувствительных нейронов, глии и специализированных клеток других тканей, которые в комплексе обеспечивают превращение влияния факторов внешней или внутренней среды (раздражение) в нервный импульс. Эта внешняя информация может поступать на рецептор в форме света, попадающего на сетчатку; механической деформации кожи, барабанной перепонки или полукружных каналов; химических веществ, проникающих в органы обоняния или вкуса.
Понятие рецептора Реце птор — сложное образование, состоящие из терминалей (нервных окончаний) и дендритов чувствительных нейронов, глии и специализированных клеток других тканей, которые в комплексе обеспечивают превращение влияния факторов внешней или внутренней среды (раздражение) в нервный импульс. Эта внешняя информация может поступать на рецептор в форме света, попадающего на сетчатку; механической деформации кожи, барабанной перепонки или полукружных каналов; химических веществ, проникающих в органы обоняния или вкуса.
 деполяризуется в ответ на") Принцип работы Большинство обычных сенсорных рецепторов (химических, температурных или механических) деполяризуется в ответ на стимул (такая же реакция, как и у обычных нейронов), деполяризация ведёт к высвобождению медиатора из аксонных окончаний. Cуществуют исключения: при освещении колбочки потенциал на её мембране возрастает — мембрана гиперполяризуется: свет, повышая потенциал, уменьшает выделение медиатора.
Принцип работы Большинство обычных сенсорных рецепторов (химических, температурных или механических) деполяризуется в ответ на стимул (такая же реакция, как и у обычных нейронов), деполяризация ведёт к высвобождению медиатора из аксонных окончаний. Cуществуют исключения: при освещении колбочки потенциал на её мембране возрастает — мембрана гиперполяризуется: свет, повышая потенциал, уменьшает выделение медиатора.
 Основные сенсорные модальности Модальность Зрение Слух Запах Вкус Вращательное ускорение Линейное ускорение Прикосновение и давление Изменения температуры Боль, тепло, холод Позиция сустава и движение Длина мышцы Мышечное напряжение АД Рецептор Палочки и колбочки Слуховые клетки Нейроны Вкусовые клетки Волосковые клетки ЧНО* ЧНО ЧНО ЧНО Орган Центральное венозное давление ЧНО Раздувание лёгких Температура крови в голове Артериальное напряжение кислорода p. H спинномозговой жидкости Осмотическое давление плазмы Артерио-венозная разница глюкозы ЧНО Нейроны Гломусные клетки Глаз Кортиев орган Обонятельная слизистая Вкусовые почки Полукружные протоки Маточка и мешочек Механорецепторы Различные ЧНО Болевые окончания Различные ЧНО Мышечное веретено Орган Гольджи Рецепторы дуги аорты и каротидного синуса Рецепторы больших вен, предсердия Рецепторы лёгких Гипоталамус Каротидные и аортальные тельца ЧНО Нейроны Клетки гипоталамуса Хемочувствительные нейроны Осмочувствительные нейроны Глюкозочувствительные клетки
Основные сенсорные модальности Модальность Зрение Слух Запах Вкус Вращательное ускорение Линейное ускорение Прикосновение и давление Изменения температуры Боль, тепло, холод Позиция сустава и движение Длина мышцы Мышечное напряжение АД Рецептор Палочки и колбочки Слуховые клетки Нейроны Вкусовые клетки Волосковые клетки ЧНО* ЧНО ЧНО ЧНО Орган Центральное венозное давление ЧНО Раздувание лёгких Температура крови в голове Артериальное напряжение кислорода p. H спинномозговой жидкости Осмотическое давление плазмы Артерио-венозная разница глюкозы ЧНО Нейроны Гломусные клетки Глаз Кортиев орган Обонятельная слизистая Вкусовые почки Полукружные протоки Маточка и мешочек Механорецепторы Различные ЧНО Болевые окончания Различные ЧНО Мышечное веретено Орган Гольджи Рецепторы дуги аорты и каротидного синуса Рецепторы больших вен, предсердия Рецепторы лёгких Гипоталамус Каротидные и аортальные тельца ЧНО Нейроны Клетки гипоталамуса Хемочувствительные нейроны Осмочувствительные нейроны Глюкозочувствительные клетки
 и вторичные (Б—Г) рецепторные клетки позвоночных А — обонятельный рецептор; В") Специализированные первичные (А) и вторичные (Б—Г) рецепторные клетки позвоночных А — обонятельный рецептор; В — вкусовой; В — фоторецептор; Г — вестибулярный и слуховой: 1 — митральная клетка, 2 — обонятельный клубочек, 3 — обонятельные нити, 4 — аксон, 5 — реснички, 6 — волокна барабанной струны, 7 — микроворсинки, 8 — биполярная клетка, 9 — волокна, 10 — ядро, 11 — внутренний членик, 12 ~ рудимент реснички, 13 — наружный членик, 14 — афферентное нервное волокно, 15 — эфферентный аксон, 16 — наружная волосковая клетка (улитка), 17 — волоски.
Специализированные первичные (А) и вторичные (Б—Г) рецепторные клетки позвоночных А — обонятельный рецептор; В — вкусовой; В — фоторецептор; Г — вестибулярный и слуховой: 1 — митральная клетка, 2 — обонятельный клубочек, 3 — обонятельные нити, 4 — аксон, 5 — реснички, 6 — волокна барабанной струны, 7 — микроворсинки, 8 — биполярная клетка, 9 — волокна, 10 — ядро, 11 — внутренний членик, 12 ~ рудимент реснички, 13 — наружный членик, 14 — афферентное нервное волокно, 15 — эфферентный аксон, 16 — наружная волосковая клетка (улитка), 17 — волоски.
 Рецепторный потенциал Изменение мембранного потенциала рецепторной клетки при воздействии раздражающего стимула
Рецепторный потенциал Изменение мембранного потенциала рецепторной клетки при воздействии раздражающего стимула
 Свойства рецепторного потенциала Регистрируется в самой нервной терминале, а не в клетках сенсорного органа Изменяется по амплитуде в зависимости от величины механического стимула (градуальность) Протекает в виде местной реакции, распространяясь по мембране электротонически Величина РП может увеличиваться посредством временной и пространственной суммации
Свойства рецепторного потенциала Регистрируется в самой нервной терминале, а не в клетках сенсорного органа Изменяется по амплитуде в зависимости от величины механического стимула (градуальность) Протекает в виде местной реакции, распространяясь по мембране электротонически Величина РП может увеличиваться посредством временной и пространственной суммации
 Трансформация сенсорных раздражений Стимул Физико химические изменения в мембране терминали или преобразование в рецепторной клетке и выделение медиатора Локальные изменения ионной проницаемости Локальная деполяризация (РП) Возникновение и проведение ПД
Трансформация сенсорных раздражений Стимул Физико химические изменения в мембране терминали или преобразование в рецепторной клетке и выделение медиатора Локальные изменения ионной проницаемости Локальная деполяризация (РП) Возникновение и проведение ПД
 Орган зрения
Орган зрения
 Зрение Мозг через орган зрения получает более 90% сенсорной информации. Фоторецепторы сетчатки глаза из всего спектра электромагнитных излучений регистрируют только волны длиной от 400 до 800 нм. Физиологическая роль органа зрения: оптический инструмент, собирающий свет от объектов внешней среды и проецирующий их изображения на сетчатку. фоторецепторы сетчатки преобразуют оптические изображения в нервные сигналы, передаваемые в зрительную кору.
Зрение Мозг через орган зрения получает более 90% сенсорной информации. Фоторецепторы сетчатки глаза из всего спектра электромагнитных излучений регистрируют только волны длиной от 400 до 800 нм. Физиологическая роль органа зрения: оптический инструмент, собирающий свет от объектов внешней среды и проецирующий их изображения на сетчатку. фоторецепторы сетчатки преобразуют оптические изображения в нервные сигналы, передаваемые в зрительную кору.
 Орган зрения Включает глазное яблоко, соединённое через зрительный нерв с мозгом, защитный аппарат (в том числе веки и слёзные железы) и аппарат движения (поперечнополосатые глазодвигательные мышцы).
Орган зрения Включает глазное яблоко, соединённое через зрительный нерв с мозгом, защитный аппарат (в том числе веки и слёзные железы) и аппарат движения (поперечнополосатые глазодвигательные мышцы).
 Глазное яблоко Стенка глазного яблока образована оболочками: ◦ в передней части расположены конъюнктива и роговица, ◦ в задней — сетчатка, сосудистая оболочка и склера. Полость глазного яблока занимает стекловидное тело. Кпереди от стекловидного тела расположен двояковыпуклый хрусталик. Между роговицей и хрусталиком находятся содержащие водянистую влагу перeдняя камера (между задней поверхностью роговицы и радужкой со зрачком) и задняя камера глаза (между радужкой и хрусталиком).
Глазное яблоко Стенка глазного яблока образована оболочками: ◦ в передней части расположены конъюнктива и роговица, ◦ в задней — сетчатка, сосудистая оболочка и склера. Полость глазного яблока занимает стекловидное тело. Кпереди от стекловидного тела расположен двояковыпуклый хрусталик. Между роговицей и хрусталиком находятся содержащие водянистую влагу перeдняя камера (между задней поверхностью роговицы и радужкой со зрачком) и задняя камера глаза (между радужкой и хрусталиком).

 Оболочки глазного яблока Роговица — прозрачная оболочка передней стенки глаза. Лимб — граница между прозрачной роговицей и непрозрачной склерой. Склера — наружная непрозрачная оболочка глазного яблока. В месте соединения склеры с роговицей расположены небольшие сообщающиеся полости, в совокупности образующие шлеммов канал, обеспечивающий отток жидкости из передней камеры глаза. Сосудистая оболочка осуществляет питание сетчатки и содержит большое количество сосудов, обусловливающих розовую окраску при офтальмоскопии. Радужка — передний вырост сосудистой оболочки. Другая часть сосудистой оболочки — ресничное тело — участвует в аккомодации зрения, регулируя форму хрусталика. ü Радужная оболочка расположена между роговицей и хрусталиком, разделяет переднюю и заднюю камеры глаза. Радужка имеет в центре отверстие — зрачок. В состав радужки входят гладкие мышцы: циркулярная суживающая (сфинктер зрачка) и радиальная расширяющая (дилататор зрачка). При раздражении парасимпатических (холинергических) нервных волокон зрачок суживается, симпатическая (адренергическая) стимуляция приводит к расширению зрачка. ü Ресничное тело находится позади от радужки вокруг хрусталика и обеспечивает аккомодацию. Основную массу ресничного тела занимает ресничная мышца (парасимпатическая иннервация). От ресничного тела по направлению к хрусталику отходят ресничные отростки, к которым прикрепляется циннова связка. При сокращении циркулярных волокон ресничной мышцы циннова связка расслабляется и хрусталик принимает более сферическую форму (увеличение рефракции).
Оболочки глазного яблока Роговица — прозрачная оболочка передней стенки глаза. Лимб — граница между прозрачной роговицей и непрозрачной склерой. Склера — наружная непрозрачная оболочка глазного яблока. В месте соединения склеры с роговицей расположены небольшие сообщающиеся полости, в совокупности образующие шлеммов канал, обеспечивающий отток жидкости из передней камеры глаза. Сосудистая оболочка осуществляет питание сетчатки и содержит большое количество сосудов, обусловливающих розовую окраску при офтальмоскопии. Радужка — передний вырост сосудистой оболочки. Другая часть сосудистой оболочки — ресничное тело — участвует в аккомодации зрения, регулируя форму хрусталика. ü Радужная оболочка расположена между роговицей и хрусталиком, разделяет переднюю и заднюю камеры глаза. Радужка имеет в центре отверстие — зрачок. В состав радужки входят гладкие мышцы: циркулярная суживающая (сфинктер зрачка) и радиальная расширяющая (дилататор зрачка). При раздражении парасимпатических (холинергических) нервных волокон зрачок суживается, симпатическая (адренергическая) стимуляция приводит к расширению зрачка. ü Ресничное тело находится позади от радужки вокруг хрусталика и обеспечивает аккомодацию. Основную массу ресничного тела занимает ресничная мышца (парасимпатическая иннервация). От ресничного тела по направлению к хрусталику отходят ресничные отростки, к которым прикрепляется циннова связка. При сокращении циркулярных волокон ресничной мышцы циннова связка расслабляется и хрусталик принимает более сферическую форму (увеличение рефракции).
 Оболочки глазного яблока Водянистая влага. В передней и задней камерах глаза находится жидкость — водянистая влага. Её выделяет ресничное тело в заднюю камеру глаза, через зрачок водянистая влага попадает в переднюю камеру глаза, направляется в пространства радужно–роговичного угла и оттекает в венозный синус склеры. При нарушении оттока водянистой влаги происходит повышение внутриглазного давления (глаукома), что вызывает ишемию сетчатки и может привести к слепоте. Стекловидное тело — прозрачная среда глаза, заполняет полость между хрусталиком и сетчатой оболочкой; стекловидное тело — гель, содержащий воду, коллаген, белок витреин и гиалуроновую кислоту. Через стекловидное тело от сетчатки к хрусталику проходит канал. Хрусталик. Эпителиальные клетки хрусталика связаны щелевыми контактами, содержат кристаллины и с возрастом утрачивают ядра и органеллы: это прозрачные хрусталиковые волокна, составляющие основную часть хрусталика. Пpозpачность хpусталика и/или его капсулы нарушается при катаракте. Сетчатая оболочка (сетчатка) — внутренняя оболочка глаза — имеет зрительный отдел, по зубчатому краю переходящий в слепой отдел, покрывающий сзади ресничное тело и радужку. У заднего края оптической оси глаза сетчатка имеет округлое жёлтое пятно (пятно сетчатки) диаметром около 2 мм. Центральная ямка — углубление в средней части жёлтого пятна — место наилучшего восприятия. Зрительный нерв выходит из сетчатки на 4 мм кнутри от жёлтого пятна. Здесь образуется диск зрительного нерва (слепое пятно), где отсутствует зрительное восприятие. В центре диска имеется углубление, в котором видны питающие сетчатку сосуды.
Оболочки глазного яблока Водянистая влага. В передней и задней камерах глаза находится жидкость — водянистая влага. Её выделяет ресничное тело в заднюю камеру глаза, через зрачок водянистая влага попадает в переднюю камеру глаза, направляется в пространства радужно–роговичного угла и оттекает в венозный синус склеры. При нарушении оттока водянистой влаги происходит повышение внутриглазного давления (глаукома), что вызывает ишемию сетчатки и может привести к слепоте. Стекловидное тело — прозрачная среда глаза, заполняет полость между хрусталиком и сетчатой оболочкой; стекловидное тело — гель, содержащий воду, коллаген, белок витреин и гиалуроновую кислоту. Через стекловидное тело от сетчатки к хрусталику проходит канал. Хрусталик. Эпителиальные клетки хрусталика связаны щелевыми контактами, содержат кристаллины и с возрастом утрачивают ядра и органеллы: это прозрачные хрусталиковые волокна, составляющие основную часть хрусталика. Пpозpачность хpусталика и/или его капсулы нарушается при катаракте. Сетчатая оболочка (сетчатка) — внутренняя оболочка глаза — имеет зрительный отдел, по зубчатому краю переходящий в слепой отдел, покрывающий сзади ресничное тело и радужку. У заднего края оптической оси глаза сетчатка имеет округлое жёлтое пятно (пятно сетчатки) диаметром около 2 мм. Центральная ямка — углубление в средней части жёлтого пятна — место наилучшего восприятия. Зрительный нерв выходит из сетчатки на 4 мм кнутри от жёлтого пятна. Здесь образуется диск зрительного нерва (слепое пятно), где отсутствует зрительное восприятие. В центре диска имеется углубление, в котором видны питающие сетчатку сосуды.
 Защитный аппарат глаза Длинные ресницы верхнего века предохраняют глаз от попадания пыли; мигательный рефлекс (моргание) осуществляется автоматически. Веки содержат мейбомиевы железы, благодаря которым края век всегда увлажнены. Конъюнктива — тонкая слизистая оболочка — выстилает как внутреннюю поверхность век, так и наружную поверхность глазного яблока. Слёзная железа выделяет слёзную жидкость, которая орошает конъюнктиву. Слёзная жидкость выполняет три основные функции: поддерживает увлажнённой конъюнктиву, действует как мягкий антисептик и вымывает пыль и мелкие частицы, попадающие на конъюнктиву. Раздражители (пыль или микроорганизмы), а также эмоции вызывают увеличение секреции слёзной жидкости до такой степени, что дренажная система (слёзные протоки, слёзный мешок, носослёзный канал) переполняется и по лицу начинают течь слёзы. Моргание — нормальный рефлекторный акт, происходящий несколько раз в минуту — способствует увлажнению конъюнктивы. Оно также возникает как защитный рефлекс при внезапном приближении какого либо предмета близко к поверхности лица.
Защитный аппарат глаза Длинные ресницы верхнего века предохраняют глаз от попадания пыли; мигательный рефлекс (моргание) осуществляется автоматически. Веки содержат мейбомиевы железы, благодаря которым края век всегда увлажнены. Конъюнктива — тонкая слизистая оболочка — выстилает как внутреннюю поверхность век, так и наружную поверхность глазного яблока. Слёзная железа выделяет слёзную жидкость, которая орошает конъюнктиву. Слёзная жидкость выполняет три основные функции: поддерживает увлажнённой конъюнктиву, действует как мягкий антисептик и вымывает пыль и мелкие частицы, попадающие на конъюнктиву. Раздражители (пыль или микроорганизмы), а также эмоции вызывают увеличение секреции слёзной жидкости до такой степени, что дренажная система (слёзные протоки, слёзный мешок, носослёзный канал) переполняется и по лицу начинают течь слёзы. Моргание — нормальный рефлекторный акт, происходящий несколько раз в минуту — способствует увлажнению конъюнктивы. Оно также возникает как защитный рефлекс при внезапном приближении какого либо предмета близко к поверхности лица.
 Аккомодация Приспособление глаза к чёткому видению предметов, расположенных на различном расстоянии. Основная роль в процессе аккомодации принадлежит хрусталику, способному изменять свою кривизну. У молодых людей преломляющая сила хрусталика может увеличиваться от 20 до 34 диоптрий. При этом хрусталик изменяет форму от умеренно выпуклой до значительно выпуклой. При взгляде на удалённые предметы (А) ресничные мышцы расслабляются, поддерживающая связка растягивает и уплощает хрусталик, придавая ему дискообразную форму. При взгляде на близкие предметы (Б) для полной фокусировки необходима более значительная кривизна хрусталика, поэтому ГМК ресничного тела сокращаются, связки расслабляются, а хрусталик в силу своей эластичности становится более выпуклым.
Аккомодация Приспособление глаза к чёткому видению предметов, расположенных на различном расстоянии. Основная роль в процессе аккомодации принадлежит хрусталику, способному изменять свою кривизну. У молодых людей преломляющая сила хрусталика может увеличиваться от 20 до 34 диоптрий. При этом хрусталик изменяет форму от умеренно выпуклой до значительно выпуклой. При взгляде на удалённые предметы (А) ресничные мышцы расслабляются, поддерживающая связка растягивает и уплощает хрусталик, придавая ему дискообразную форму. При взгляде на близкие предметы (Б) для полной фокусировки необходима более значительная кривизна хрусталика, поэтому ГМК ресничного тела сокращаются, связки расслабляются, а хрусталик в силу своей эластичности становится более выпуклым.
 Острота зрения Точность, с которой виден объект; теоретически объект должен быть такого размера, чтобы мог простимулировать одну палочку или колбочку. Оба глаза действуют вместе (бинокулярное зрение) для передачи зрительной информации в зрительные центры коры больших полушарий, где зрительный образ оценивается в трёх измерениях
Острота зрения Точность, с которой виден объект; теоретически объект должен быть такого размера, чтобы мог простимулировать одну палочку или колбочку. Оба глаза действуют вместе (бинокулярное зрение) для передачи зрительной информации в зрительные центры коры больших полушарий, где зрительный образ оценивается в трёх измерениях
 Зрачковый рефлекс. Зрачок — круглое отверстие в радужной оболочке — очень быстро меняется в размере в зависимости от количества света, падающего на сетчатку. Просвет зрачка может изменяться от 1 мм до 8 мм. Это придаёт зрачку свойства диафрагмы. Сетчатка очень чувствительна к свету, слишком большое количество света искажает цвета и раздражает глаз. Изменяя просвет, зрачок регулирует количество света, попадающего в глаз. Яркий свет вызывает безусловнорефлекторную вегетативную реакцию, замыкающуюся в среднем мозге: сфинктер зрачка в радужной оболочке обоих глаз сокращается, а дилататор зрачка расслабляется, в результате диаметр зрачка уменьшается. Плохое освещение заставляет оба зрачка расшириться, чтобы достаточное количество света могло достичь сетчатки и возбудить фоторецепторы.
Зрачковый рефлекс. Зрачок — круглое отверстие в радужной оболочке — очень быстро меняется в размере в зависимости от количества света, падающего на сетчатку. Просвет зрачка может изменяться от 1 мм до 8 мм. Это придаёт зрачку свойства диафрагмы. Сетчатка очень чувствительна к свету, слишком большое количество света искажает цвета и раздражает глаз. Изменяя просвет, зрачок регулирует количество света, попадающего в глаз. Яркий свет вызывает безусловнорефлекторную вегетативную реакцию, замыкающуюся в среднем мозге: сфинктер зрачка в радужной оболочке обоих глаз сокращается, а дилататор зрачка расслабляется, в результате диаметр зрачка уменьшается. Плохое освещение заставляет оба зрачка расшириться, чтобы достаточное количество света могло достичь сетчатки и возбудить фоторецепторы.
 Содружественная реакция зрачков У здоровых людей зрачки обоих глаз одинакового размера. Освещение одного глаза ведет к сужению зрачка и другого глаза. Такая реакция называется содружественной реакцией зрачков. При некоторых заболеваниях размеры зрачков обоих глаз различны (анизокория).
Содружественная реакция зрачков У здоровых людей зрачки обоих глаз одинакового размера. Освещение одного глаза ведет к сужению зрачка и другого глаза. Такая реакция называется содружественной реакцией зрачков. При некоторых заболеваниях размеры зрачков обоих глаз различны (анизокория).
 Вегетативная иннервация глаза Парасимпатические преганглионарные волокна в составе глазодвигательного нерва проходят к ресничному ганглию и от него постганглионарные волокна в виде ресничных нервов поступают в глаз. Волокна этих нервов иннервируют сфинктер зрачка. Соответственно ацетилхолин и эзерин вызывают сужение зрачка, а блокада холинорецепторов сфинктера радужки атропином приводит к расширению зрачка. Симпатическая иннервация глаза происходит из клеток бокового рога первых грудных сегментов спинного мозга. Отсюда симпатические волокна проходят в верхний симпатический ганглий, где они синаптически контактируют с постганглионарными нейронами. Постганглионарные симпатические волокна распространяются по поверхности сонной артерии и её ветвей и достигают глаза. Здесь симпатические волокна иннервируют дилататор зрачка, и возбуждение симпатических волокон расширяет зрачок. Адреналин и его аналоги также расширяют зрачок. Зрачки расширяются при гипоксии, болевом шоке, при эмоциях ярости и страха. Симпатические волокна иннервируют также некоторые наружные глазные мышцы.
Вегетативная иннервация глаза Парасимпатические преганглионарные волокна в составе глазодвигательного нерва проходят к ресничному ганглию и от него постганглионарные волокна в виде ресничных нервов поступают в глаз. Волокна этих нервов иннервируют сфинктер зрачка. Соответственно ацетилхолин и эзерин вызывают сужение зрачка, а блокада холинорецепторов сфинктера радужки атропином приводит к расширению зрачка. Симпатическая иннервация глаза происходит из клеток бокового рога первых грудных сегментов спинного мозга. Отсюда симпатические волокна проходят в верхний симпатический ганглий, где они синаптически контактируют с постганглионарными нейронами. Постганглионарные симпатические волокна распространяются по поверхности сонной артерии и её ветвей и достигают глаза. Здесь симпатические волокна иннервируют дилататор зрачка, и возбуждение симпатических волокон расширяет зрачок. Адреналин и его аналоги также расширяют зрачок. Зрачки расширяются при гипоксии, болевом шоке, при эмоциях ярости и страха. Симпатические волокна иннервируют также некоторые наружные глазные мышцы.
 Пресбиопия Когда человек становится старше, хрусталик разрастается, становится толще и менее эластичным. Способность хрусталика изменять свою форму также уменьшается. Сила аккомодации падает с 14 диоптрий у ребенка до менее 2 диоптрий у человека в возрасте от 45 до 50 лет и до 0–в возрасте 70 лет. Таким образом, хрусталик утрачивает способность аккомодации, и это состояние называется пресбиопией (старческая дальнозоркость). Когда человек достигает состояния пресбиопии, каждый глаз остаётся с постоянным фокусным расстоянием; это расстояние зависит от физических характеристик глаз каждого отдельного человека. Поэтому пожилые люди вынуждены пользоваться очками с двояковыпуклыми линзами
Пресбиопия Когда человек становится старше, хрусталик разрастается, становится толще и менее эластичным. Способность хрусталика изменять свою форму также уменьшается. Сила аккомодации падает с 14 диоптрий у ребенка до менее 2 диоптрий у человека в возрасте от 45 до 50 лет и до 0–в возрасте 70 лет. Таким образом, хрусталик утрачивает способность аккомодации, и это состояние называется пресбиопией (старческая дальнозоркость). Когда человек достигает состояния пресбиопии, каждый глаз остаётся с постоянным фокусным расстоянием; это расстояние зависит от физических характеристик глаз каждого отдельного человека. Поэтому пожилые люди вынуждены пользоваться очками с двояковыпуклыми линзами
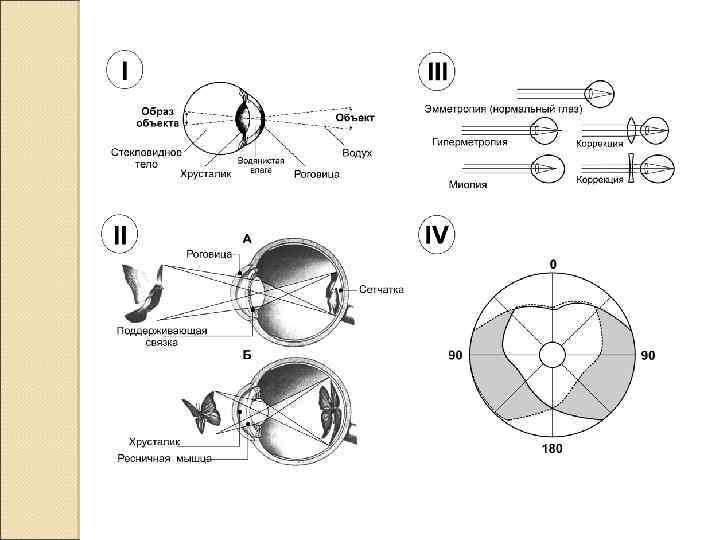
 соответствует нормальному глазу, если параллельные лучи от отдалённых предметов") Аномалии рефракции Эмметропия (нормальное зрение) соответствует нормальному глазу, если параллельные лучи от отдалённых предметов фокусируются на сетчатке, когда ресничная мышца полностью расслаблена. Это значит, что эмметропический глаз может видеть все отдалённые объекты очень ясно и легко переходить (посредством аккомодации) на ясное видение близлежащих предметов. Гиперметропия (дальнозоркость) может быть обусловлена слишком коротким глазным яблоком или в более редких случаях тем, что глаз имеет слишком малоэластичный хрусталик. В дальнозорком глазу продольная ось глаза короче, и луч от отдалённых предметов фокусируется за сетчаткой. Этот недостаток рефракции компенсируется дальнозорким человеком аккомодационным усилием. Дальнозоркий человек напрягает аккомодационную мышцу, рассматривая далёкие объекты. Попытки рассматривать близкие предметы вызывают чрезмерное напряжение аккомодации. Для работы с близкорасположенными предметами и чтения дальнозоркие люди должны пользоваться очками с двояковыпуклыми линзами.
Аномалии рефракции Эмметропия (нормальное зрение) соответствует нормальному глазу, если параллельные лучи от отдалённых предметов фокусируются на сетчатке, когда ресничная мышца полностью расслаблена. Это значит, что эмметропический глаз может видеть все отдалённые объекты очень ясно и легко переходить (посредством аккомодации) на ясное видение близлежащих предметов. Гиперметропия (дальнозоркость) может быть обусловлена слишком коротким глазным яблоком или в более редких случаях тем, что глаз имеет слишком малоэластичный хрусталик. В дальнозорком глазу продольная ось глаза короче, и луч от отдалённых предметов фокусируется за сетчаткой. Этот недостаток рефракции компенсируется дальнозорким человеком аккомодационным усилием. Дальнозоркий человек напрягает аккомодационную мышцу, рассматривая далёкие объекты. Попытки рассматривать близкие предметы вызывают чрезмерное напряжение аккомодации. Для работы с близкорасположенными предметами и чтения дальнозоркие люди должны пользоваться очками с двояковыпуклыми линзами.
 представляет тот случай, когда ресничная мышца полностью расслаблена, и лучи") Аномалии рефракции Миопия (близорукость) представляет тот случай, когда ресничная мышца полностью расслаблена, и лучи света от далекого объекта фокусируются впереди сетчатки. Близорукость возникает либо вследствие слишком длинного глазного яблока, либо в результате большой преломляющей силы хрусталика глаза. Не существует механизма, посредством которого глаз смог бы уменьшить преломляющую силу хрусталика в условиях полностью расслабленной ресничной мышцы. Однако если объект находится рядом с глазами, то близорукий человек может использовать механизм аккомодации для чёткого фокусирования объекта на сетчатке. Следовательно, близорукий человек имеет ограничения только в отношении ясной точки «дальнего видения» . Для ясного видения вдаль близорукому человеку необходимо использовать очки с двояковогнутыми линзами. Астигматизм — неодинаковое преломление лучей в разных направлениях, вызванное различной кривизной сферической поверхности роговицы. Аккомодация глаза не в силах преодолеть астигматизм, потому что кривизна хрусталика при аккомодации изменяется одинаково. Для компенсации недостатков рефракции роговицы применяют специальные цилиндрические линзы.
Аномалии рефракции Миопия (близорукость) представляет тот случай, когда ресничная мышца полностью расслаблена, и лучи света от далекого объекта фокусируются впереди сетчатки. Близорукость возникает либо вследствие слишком длинного глазного яблока, либо в результате большой преломляющей силы хрусталика глаза. Не существует механизма, посредством которого глаз смог бы уменьшить преломляющую силу хрусталика в условиях полностью расслабленной ресничной мышцы. Однако если объект находится рядом с глазами, то близорукий человек может использовать механизм аккомодации для чёткого фокусирования объекта на сетчатке. Следовательно, близорукий человек имеет ограничения только в отношении ясной точки «дальнего видения» . Для ясного видения вдаль близорукому человеку необходимо использовать очки с двояковогнутыми линзами. Астигматизм — неодинаковое преломление лучей в разных направлениях, вызванное различной кривизной сферической поверхности роговицы. Аккомодация глаза не в силах преодолеть астигматизм, потому что кривизна хрусталика при аккомодации изменяется одинаково. Для компенсации недостатков рефракции роговицы применяют специальные цилиндрические линзы.
 и сетчатки глаза (Б) С — колбочка; R — палочки; Н") Строение фоторецептора (А) и сетчатки глаза (Б) С — колбочка; R — палочки; Н — горизонтальная клетка: MB, RB; FB — биполярные клет ки; А — амакриновая клетка; MG и DG — ганглиозные клетки; 1 — фрагмент диска; 2 — наружный сегмент; 3 — внутренний сегмент; 4 — митохондрии внутреннего сегмента; 5 — корешок реснички; 6 — центриоли; 7 — микротрубочки реснички; 8 — наружная плазматическая мембрана; 9 — диски наружного сегмента.
Строение фоторецептора (А) и сетчатки глаза (Б) С — колбочка; R — палочки; Н — горизонтальная клетка: MB, RB; FB — биполярные клет ки; А — амакриновая клетка; MG и DG — ганглиозные клетки; 1 — фрагмент диска; 2 — наружный сегмент; 3 — внутренний сегмент; 4 — митохондрии внутреннего сегмента; 5 — корешок реснички; 6 — центриоли; 7 — микротрубочки реснички; 8 — наружная плазматическая мембрана; 9 — диски наружного сегмента.
 Родопсин и активация ионных каналов А. Молекула опсина содержит 7 трансмембранных альфа спиральных участков. Б. Трансмембранный белок родопсин и его связь с G‑белком (трансдуцин) в плазмолемме фоторецепторной клетки. Возбуждённый фотонами родопсин активирует G‑белок. При этом гуанозиндифосфат, связанный с a‑СЕ G‑белка, заменяется на ГТФ. Отщеплённые a‑СЕ и b‑СЕ действуют на фосфодиэстеразу и заставляют её превращать ц. ГМФ в гуанозинмонофосфат. Это закрывает Na+‑каналы, и ионы Na+ не могут попасть в клетку, что приводит к её гиперполяризации. R — родопсин; a, b и g — СЕ G‑белка; A — агонист (в данном случае кванты света); E — фермент– эффектор фосфодиэстераза. В. Схема палочки. В наружном сегменте расположена стопка дисков, содержащих зрительный пигмент родопсин. Мембрана дисков и клеточная мембрана разобщены. Свет (hn) активирует родопсин (Rh*) в дисках, что закрывает Na+‑каналы в клеточной мембране и снижает вход Na+ в клетку.
Родопсин и активация ионных каналов А. Молекула опсина содержит 7 трансмембранных альфа спиральных участков. Б. Трансмембранный белок родопсин и его связь с G‑белком (трансдуцин) в плазмолемме фоторецепторной клетки. Возбуждённый фотонами родопсин активирует G‑белок. При этом гуанозиндифосфат, связанный с a‑СЕ G‑белка, заменяется на ГТФ. Отщеплённые a‑СЕ и b‑СЕ действуют на фосфодиэстеразу и заставляют её превращать ц. ГМФ в гуанозинмонофосфат. Это закрывает Na+‑каналы, и ионы Na+ не могут попасть в клетку, что приводит к её гиперполяризации. R — родопсин; a, b и g — СЕ G‑белка; A — агонист (в данном случае кванты света); E — фермент– эффектор фосфодиэстераза. В. Схема палочки. В наружном сегменте расположена стопка дисков, содержащих зрительный пигмент родопсин. Мембрана дисков и клеточная мембрана разобщены. Свет (hn) активирует родопсин (Rh*) в дисках, что закрывает Na+‑каналы в клеточной мембране и снижает вход Na+ в клетку.
 Родопсин и активация ионных каналов
Родопсин и активация ионных каналов
 Ионные основы фоторецепторных потенциалов В темноте Na+‑каналы мембраны наружных сегментов палочек и колбочек открыты, и ток течёт из цитоплазмы внутренних сегментов в мембраны наружных сегментов. Ток течёт также в синаптическое окончание фоторецептора, вызывая постоянное выделение нейромедиатора. Na+, K+– насос, находящийся во внутреннем сегменте, поддерживает ионное равновесие, компенсируя выход Na+ входом K+. Таким образом, в темноте ионные каналы поддерживаются в открытом состоянии и потоки внутрь клетки Na+ и Ca 2+ через открытые каналы обеспечивают появление тока (темновой ток). На свету, т. е. когда свет возбуждает наружный сегмент, Na+‑каналы закрываются и возникает гиперполяризационный рецепторный потенциал. Этот потенциал, появившийся на мембране наружного сегмента, распространяется до синаптического окончания фоторецептора и уменьшает выделение синаптического медиатора — глутамата. Это немедленно приводит к появлению ПД в аксонах ганглиозных клетках. Таким образом, гиперполяризация плазмолеммы — следствие закрытия ионных каналов.
Ионные основы фоторецепторных потенциалов В темноте Na+‑каналы мембраны наружных сегментов палочек и колбочек открыты, и ток течёт из цитоплазмы внутренних сегментов в мембраны наружных сегментов. Ток течёт также в синаптическое окончание фоторецептора, вызывая постоянное выделение нейромедиатора. Na+, K+– насос, находящийся во внутреннем сегменте, поддерживает ионное равновесие, компенсируя выход Na+ входом K+. Таким образом, в темноте ионные каналы поддерживаются в открытом состоянии и потоки внутрь клетки Na+ и Ca 2+ через открытые каналы обеспечивают появление тока (темновой ток). На свету, т. е. когда свет возбуждает наружный сегмент, Na+‑каналы закрываются и возникает гиперполяризационный рецепторный потенциал. Этот потенциал, появившийся на мембране наружного сегмента, распространяется до синаптического окончания фоторецептора и уменьшает выделение синаптического медиатора — глутамата. Это немедленно приводит к появлению ПД в аксонах ганглиозных клетках. Таким образом, гиперполяризация плазмолеммы — следствие закрытия ионных каналов.
 Фоторецепция В состав дисков фоторецепторных клеток входят зрительные пигменты, в том числе родопсин палочек. Родопсин состоит из белковой части (опсин) и хромофора — 11 цис ретиналя, под действием фотонов переходящего в транс ретиналь (фотоизомеризация). При попадании квантов света на наружные сегменты в фоторецепторных клетках последовательно происходят следующие события : активация родопсина в результате фотоизомеризации каталитическая активация G‑белка (Gt, трансдуцин) родопсином активация фосфодиэстеразы при связывании с Gta гидролиз ц. ГМФ–фосфодиэстеразой переход ц. ГМФ–зависимых Na+‑каналов из открытого состояния в закрытое гиперполяризация плазмолеммы фоторецепторной клетки передача сигнала на биполярные клетки
Фоторецепция В состав дисков фоторецепторных клеток входят зрительные пигменты, в том числе родопсин палочек. Родопсин состоит из белковой части (опсин) и хромофора — 11 цис ретиналя, под действием фотонов переходящего в транс ретиналь (фотоизомеризация). При попадании квантов света на наружные сегменты в фоторецепторных клетках последовательно происходят следующие события : активация родопсина в результате фотоизомеризации каталитическая активация G‑белка (Gt, трансдуцин) родопсином активация фосфодиэстеразы при связывании с Gta гидролиз ц. ГМФ–фосфодиэстеразой переход ц. ГМФ–зависимых Na+‑каналов из открытого состояния в закрытое гиперполяризация плазмолеммы фоторецепторной клетки передача сигнала на биполярные клетки
 Связи между клетками сетчатки Фоторецепторы образуют синапсы с биполярными нейронами. В области центральной ямки расположены преимущественно колбочки. Каждая колбочка центральной ямки образует синапс только с одним биполярным нейроном. Биполярные нейроны синаптически связаны с ганглиозными и амакринными клетками. Популярна концепция о том, что ограниченное число биполярных клеток передает информацию 16 типам ганглиозных клеток при участии не менее 20 типов амакринных клеток. Горизонтальные клетки формируют синаптические контакты с отростками фоторецепторов. Горизонтальные клетки получают информацию от колбочек и передают её также колбочкам. Соседние горизонтальные клетки связаны между собой щелевыми контактами. Ганглиозные клетки — крупные мультиполярные нейроны многих разновидностей. Биполярные и амакринные клетки образуют синапсы на ганглиозных нейронах. Их аксоны образуют зрительный нерв.
Связи между клетками сетчатки Фоторецепторы образуют синапсы с биполярными нейронами. В области центральной ямки расположены преимущественно колбочки. Каждая колбочка центральной ямки образует синапс только с одним биполярным нейроном. Биполярные нейроны синаптически связаны с ганглиозными и амакринными клетками. Популярна концепция о том, что ограниченное число биполярных клеток передает информацию 16 типам ганглиозных клеток при участии не менее 20 типов амакринных клеток. Горизонтальные клетки формируют синаптические контакты с отростками фоторецепторов. Горизонтальные клетки получают информацию от колбочек и передают её также колбочкам. Соседние горизонтальные клетки связаны между собой щелевыми контактами. Ганглиозные клетки — крупные мультиполярные нейроны многих разновидностей. Биполярные и амакринные клетки образуют синапсы на ганглиозных нейронах. Их аксоны образуют зрительный нерв.
 Связи между клетками сетчатки Амакринные клетки. Их перикарионы находятся во внутренней части внутреннего ядерного слоя в области синапсов между биполярными и ганглиозными клетками. Эти клетки получают информацию от биполярных нейронов и передают её ганглиозным нейронам. Функции десятков подтипов амакринных клеток полностью не выяснены. Часть клеток участвует в формировании пути палочкового зрения в сетчатке, другие отвечают за начало и окончание световых сигналов, третьи следят за правильностью движения светового пятна. В целом амакринные клетки выполняют функцию вставочных нейронов, осуществляющих первоначальный анализ световых сигналов. Мюллеровские клетки. Кроме нейронов, сетчатка содержит крупные клетки радиальной глии — мюллеровские клетки. Их ядра расположены на уровне центральной части внутреннего ядерного слоя. Наружные отростки заканчиваются микроворсинками, образуя наружный пограничный слой. Внутренние отростки имеют расширение (ножку) во внутреннем пограничном слое на границе со стекловидным телом. Глиальные клетки играют важную роль в регуляции ионного гомеостаза сетчатки. В частности, они снижают концентрацию K+ во внеклеточном пространстве, где концентрация этих ионов при световом раздражении резко увеличивается. Плазматическая мембрана мюллеровских клеток в области ножки характеризуется высокой проницаемостью для ионов K+. Мюллеровская клетка захватывает K+ из наружных слоёв сетчатки и направляет поток этих ионов через свою ножку в жидкость стекловидного тела.
Связи между клетками сетчатки Амакринные клетки. Их перикарионы находятся во внутренней части внутреннего ядерного слоя в области синапсов между биполярными и ганглиозными клетками. Эти клетки получают информацию от биполярных нейронов и передают её ганглиозным нейронам. Функции десятков подтипов амакринных клеток полностью не выяснены. Часть клеток участвует в формировании пути палочкового зрения в сетчатке, другие отвечают за начало и окончание световых сигналов, третьи следят за правильностью движения светового пятна. В целом амакринные клетки выполняют функцию вставочных нейронов, осуществляющих первоначальный анализ световых сигналов. Мюллеровские клетки. Кроме нейронов, сетчатка содержит крупные клетки радиальной глии — мюллеровские клетки. Их ядра расположены на уровне центральной части внутреннего ядерного слоя. Наружные отростки заканчиваются микроворсинками, образуя наружный пограничный слой. Внутренние отростки имеют расширение (ножку) во внутреннем пограничном слое на границе со стекловидным телом. Глиальные клетки играют важную роль в регуляции ионного гомеостаза сетчатки. В частности, они снижают концентрацию K+ во внеклеточном пространстве, где концентрация этих ионов при световом раздражении резко увеличивается. Плазматическая мембрана мюллеровских клеток в области ножки характеризуется высокой проницаемостью для ионов K+. Мюллеровская клетка захватывает K+ из наружных слоёв сетчатки и направляет поток этих ионов через свою ножку в жидкость стекловидного тела.
 Нейромедиаторы Нейроны сетчатки синтезируют ацетилхолин, дофамин, L глутаминовую кислоту, глицин, g аминомасляную кислоту. Некоторые нейроны содержат серотонин, его аналоги (индоламины) и нейропептиды. Палочки и колбочки в синапсах с биполярными клетками секретируют глутамат. Разные амакринные клетки выделяют ГАМК, глицин, дофамин, ацетилхолин и индоламин, оказывающие тормозные эффекты. Нейромедиаторы для биполярных и горизонтальных не идентифицированы
Нейромедиаторы Нейроны сетчатки синтезируют ацетилхолин, дофамин, L глутаминовую кислоту, глицин, g аминомасляную кислоту. Некоторые нейроны содержат серотонин, его аналоги (индоламины) и нейропептиды. Палочки и колбочки в синапсах с биполярными клетками секретируют глутамат. Разные амакринные клетки выделяют ГАМК, глицин, дофамин, ацетилхолин и индоламин, оказывающие тормозные эффекты. Нейромедиаторы для биполярных и горизонтальных не идентифицированы
 Проведение сигналов от палочек и колбочек Зрительные пути от палочек и колбочек имеют некоторые отличия. Нейроны и аксоны ганглиозных клеток, проводящие зрительные сигналы от колбочек, значительно толще, чем те, что проводят сигналы от палочек. Скорость проведения сигналов для колбочкового зрения в два раза выше, чем для проведения сигналов палочкового зрения. Для колбочек, эволюционно представляющих молодую и быструю систему проведения, нейронный путь имеет три звена: колбочки биполярные клетки ганглиозные нейроны. Более древняя палочковая система зрения имеет зрительный путь из 4 звеньев: палочки биполярные клетки амакринные клетки ганглиозные клетки
Проведение сигналов от палочек и колбочек Зрительные пути от палочек и колбочек имеют некоторые отличия. Нейроны и аксоны ганглиозных клеток, проводящие зрительные сигналы от колбочек, значительно толще, чем те, что проводят сигналы от палочек. Скорость проведения сигналов для колбочкового зрения в два раза выше, чем для проведения сигналов палочкового зрения. Для колбочек, эволюционно представляющих молодую и быструю систему проведения, нейронный путь имеет три звена: колбочки биполярные клетки ганглиозные нейроны. Более древняя палочковая система зрения имеет зрительный путь из 4 звеньев: палочки биполярные клетки амакринные клетки ганглиозные клетки
 Зрительные образы Сетчатка вовлечена в формирование трёх зрительных образов. Первый образ формируется под действием света на уровне фоторецепторов, превращается во второй образ на уровне биполярных клеток, в ганглиозных нейронах формируется третий образ. В формировании второго образа принимают также участие горизонтальные клетки, а в образовании третьего задействованы амакринные клетки
Зрительные образы Сетчатка вовлечена в формирование трёх зрительных образов. Первый образ формируется под действием света на уровне фоторецепторов, превращается во второй образ на уровне биполярных клеток, в ганглиозных нейронах формируется третий образ. В формировании второго образа принимают также участие горизонтальные клетки, а в образовании третьего задействованы амакринные клетки
 Проводящие пути зрительного анализатора 1 Левая половина зрительного поля, 2 Правая половина зрительного поля, 3 Глаз, 4 Сетчатка, 5 Зрительные нервы, 6 Глазодвигательный нерв, 7 Хиазма, 8 Зрительный тракт, 9 Латеральное коленчатое тело, 10 Верхние бугры четверохолмия, 11 Неспецифический зрительный путь, 12 Зрительная кора головного мозга
Проводящие пути зрительного анализатора 1 Левая половина зрительного поля, 2 Правая половина зрительного поля, 3 Глаз, 4 Сетчатка, 5 Зрительные нервы, 6 Глазодвигательный нерв, 7 Хиазма, 8 Зрительный тракт, 9 Латеральное коленчатое тело, 10 Верхние бугры четверохолмия, 11 Неспецифический зрительный путь, 12 Зрительная кора головного мозга
. Аксоны ганглиозных клеток в составе зрительных нервов") Основной путь к зрительной коре (новая система). Аксоны ганглиозных клеток в составе зрительных нервов и (после перекреста) в составе зрительных трактов достигают латеральных коленчатых тел (ЛКТ). При этом волокна от носовой половины сетчатки в зрительном перекресте не переходят на другую сторону. В левом ЛКТ (ипсилатеральном глазу) волокна от носовой половины сетчатки левого глаза и волокна от височной половины сетчатки правого глаза синаптически контактируют с нейронами ЛКТ, аксоны которых образуют коленчато–шпорный тракт (зрительная лучистость). Коленчато шпорные волокна проходят к первичной зрительной коре той же стороны. Аналогично организованы пути от правого глаза
Основной путь к зрительной коре (новая система). Аксоны ганглиозных клеток в составе зрительных нервов и (после перекреста) в составе зрительных трактов достигают латеральных коленчатых тел (ЛКТ). При этом волокна от носовой половины сетчатки в зрительном перекресте не переходят на другую сторону. В левом ЛКТ (ипсилатеральном глазу) волокна от носовой половины сетчатки левого глаза и волокна от височной половины сетчатки правого глаза синаптически контактируют с нейронами ЛКТ, аксоны которых образуют коленчато–шпорный тракт (зрительная лучистость). Коленчато шпорные волокна проходят к первичной зрительной коре той же стороны. Аналогично организованы пути от правого глаза
 Аксоны ганглиозных нейронов сетчатки проходят также в некоторые древние области") Другие пути (старая система) Аксоны ганглиозных нейронов сетчатки проходят также в некоторые древние области мозга: ◦ к надперекрестным ядрам гипоталамуса (контроль и синхронизация циркадных ритмов); ◦ в ядра покрышки (рефлекторные движения глаз при фокусировании объекта, активация зрачкового рефлекса); ◦ в верхнее двухолмие (контроль быстрых направленных движений обоих глаз); ◦ в ЛКТ и окружающие их области (контроль поведенческих реакций).
Другие пути (старая система) Аксоны ганглиозных нейронов сетчатки проходят также в некоторые древние области мозга: ◦ к надперекрестным ядрам гипоталамуса (контроль и синхронизация циркадных ритмов); ◦ в ядра покрышки (рефлекторные движения глаз при фокусировании объекта, активация зрачкового рефлекса); ◦ в верхнее двухолмие (контроль быстрых направленных движений обоих глаз); ◦ в ЛКТ и окружающие их области (контроль поведенческих реакций).
 Часть новой зрительной системы, где оканчиваются все волокна, проходящие в") Латеральное коленчатое тело (ЛКТ) Часть новой зрительной системы, где оканчиваются все волокна, проходящие в составе зрительного тракта. ЛКТ выполняет функцию передачи информации из зрительного тракта к зрительной коре, в точности сохраняя топологию (пространственное расположение) разного уровня путей из сетчатки. Другая функция ЛКТ заключается в контроле над количеством информации, поступающей к коре. Сигналы для осуществления ЛКТ входного контроля поступают в ЛКТ в виде обратной импульсации из первичной зрительной коры и из ретикулярной области среднего мозга.
Латеральное коленчатое тело (ЛКТ) Часть новой зрительной системы, где оканчиваются все волокна, проходящие в составе зрительного тракта. ЛКТ выполняет функцию передачи информации из зрительного тракта к зрительной коре, в точности сохраняя топологию (пространственное расположение) разного уровня путей из сетчатки. Другая функция ЛКТ заключается в контроле над количеством информации, поступающей к коре. Сигналы для осуществления ЛКТ входного контроля поступают в ЛКТ в виде обратной импульсации из первичной зрительной коры и из ретикулярной области среднего мозга.
 Зрительная кора Первичная зрительная воспринимающая область располагается на соответствующей стороне шпорной борозды. Подобно другим частям новой коры, зрительная кора состоит из шести слоев, волокна коленчато–шпорного пути заканчиваются преимущественно на нейронах слоя IV. Этот слой подразделяется на подслои, принимающие волокна от ганглиозных клеток типа Y и X. В первичной зрительной коре (поле 17 по Бродманну) и зрительной области II (поле 18) осуществляется анализ трёхмерного расположения объектов, величины объектов, детализация предметов и их окраски, движения объектов и т. д.
Зрительная кора Первичная зрительная воспринимающая область располагается на соответствующей стороне шпорной борозды. Подобно другим частям новой коры, зрительная кора состоит из шести слоев, волокна коленчато–шпорного пути заканчиваются преимущественно на нейронах слоя IV. Этот слой подразделяется на подслои, принимающие волокна от ганглиозных клеток типа Y и X. В первичной зрительной коре (поле 17 по Бродманну) и зрительной области II (поле 18) осуществляется анализ трёхмерного расположения объектов, величины объектов, детализация предметов и их окраски, движения объектов и т. д.
 Особенности строения зрительной коры Колонки и полоски. Зрительная кора содержит несколько миллионов вертикальных первичных колонок, каждая колонка имеет диаметр от 30 до 50 мкм и содержит около 1000 нейронов. Нейронные колонки формируют переплетённые между собой полоски шириной в 0, 5 мм. Цветовые колонкообразные структуры. Среди первичных зрительных колонок распределяются вторичные области — колонкоподобные образования ( «цветовые сгустки» ). «Цветовые сгустки» получают сигналы от прилежащих колонок и специфически активируются цветовыми сигналами. Взаимодействие зрительных сигналов из двух глаз. Зрительные сигналы, поступающие в мозг, остаются раздельными до их вхождения в слой IV первичной зрительной коры. Сигналы из одного глаза входят в колонки каждой полоски, то же самое происходит с сигналами из другого глаза. В ходе взаимодействия зрительных сигналов зрительная кора расшифровывает расположение двух зрительных образов, находит их корреспондирующие точки (точки в одинаковых участках сетчатки обоих глаз) и приспосабливает расшифрованную информацию для определения расстояния до объектов.
Особенности строения зрительной коры Колонки и полоски. Зрительная кора содержит несколько миллионов вертикальных первичных колонок, каждая колонка имеет диаметр от 30 до 50 мкм и содержит около 1000 нейронов. Нейронные колонки формируют переплетённые между собой полоски шириной в 0, 5 мм. Цветовые колонкообразные структуры. Среди первичных зрительных колонок распределяются вторичные области — колонкоподобные образования ( «цветовые сгустки» ). «Цветовые сгустки» получают сигналы от прилежащих колонок и специфически активируются цветовыми сигналами. Взаимодействие зрительных сигналов из двух глаз. Зрительные сигналы, поступающие в мозг, остаются раздельными до их вхождения в слой IV первичной зрительной коры. Сигналы из одного глаза входят в колонки каждой полоски, то же самое происходит с сигналами из другого глаза. В ходе взаимодействия зрительных сигналов зрительная кора расшифровывает расположение двух зрительных образов, находит их корреспондирующие точки (точки в одинаковых участках сетчатки обоих глаз) и приспосабливает расшифрованную информацию для определения расстояния до объектов.
 Специализация нейронов В колонках зрительной коры имеются нейроны, выполняющие совершенно определённые функции [например, анализ контраста (в том числе цветового), границ и направлений линий зрительного образа и др. ]. В основе детекторной функции нейронов лежит, прежде всего, анализ контраста зрительного образа. Первоначально выявляются границы, степень и градиент контраста, после чего другие нейроны детализируют зрительный образ объекта. Располагающиеся в наружных слоях зрительной коры нейроны являются детекторами линий, определённой длины, углов и других форм зрительных образов. Некоторые группы нейронов определяют границы линий и их направление (детекторы ориентаций). Это простые клетки. Другие клетки определяют направление линий при их вертикальном или боковом смещении (комплексные клетки). Цвет определяется тем же способом, каким детектируются линии, то есть посредством цветового контраста. Например, нейронная область для красного цвета контрастирует с областью для зелёного цвета, область голубого цвета контрастирует с областью красного и т. д. Все цвета контрастируют против белого цвета. Механизм цветового контраста зависит от того, что контрастирующие (оппонентные) цвета обоюдно возбуждают специфические нейроны. Полагают, что начальные детали цветового контраста детектируются простыми клетками, в то время как более сложные цветовые контрасты определяются комплексами или даже сверхкомплексами нейронов.
Специализация нейронов В колонках зрительной коры имеются нейроны, выполняющие совершенно определённые функции [например, анализ контраста (в том числе цветового), границ и направлений линий зрительного образа и др. ]. В основе детекторной функции нейронов лежит, прежде всего, анализ контраста зрительного образа. Первоначально выявляются границы, степень и градиент контраста, после чего другие нейроны детализируют зрительный образ объекта. Располагающиеся в наружных слоях зрительной коры нейроны являются детекторами линий, определённой длины, углов и других форм зрительных образов. Некоторые группы нейронов определяют границы линий и их направление (детекторы ориентаций). Это простые клетки. Другие клетки определяют направление линий при их вертикальном или боковом смещении (комплексные клетки). Цвет определяется тем же способом, каким детектируются линии, то есть посредством цветового контраста. Например, нейронная область для красного цвета контрастирует с областью для зелёного цвета, область голубого цвета контрастирует с областью красного и т. д. Все цвета контрастируют против белого цвета. Механизм цветового контраста зависит от того, что контрастирующие (оппонентные) цвета обоюдно возбуждают специфические нейроны. Полагают, что начальные детали цветового контраста детектируются простыми клетками, в то время как более сложные цветовые контрасты определяются комплексами или даже сверхкомплексами нейронов.
 Цветовосприятие Существует три типа колбочек, каждый из которых содержит только один из трёх разных (красный, зелёный и синий) зрительных пигментов Трихромазия — возможность различать любые цвета — определяется присутствием в сетчатке всех трёх зрительных пигментов (для красного, зелёного и синего — первичные цвета). Эти основы теории цветного зрения предложил Томас Янг (1802) и развил Герман Гельмгольц Дихромазии (цветовая слепота, или дальтонизм)— дефекты цветового восприятия (преимущественно у мужчин; например, в Европе разные дефекты у мужчин составляют 8% общей популяции) по одному из первичных цветов — подразделяют на протанопии, дейтанопии и тританопии (от греч. первый, второй и третий [имеются в виду порядковые номера первичных цветов: соответственно красный, зелёный, синий]).
Цветовосприятие Существует три типа колбочек, каждый из которых содержит только один из трёх разных (красный, зелёный и синий) зрительных пигментов Трихромазия — возможность различать любые цвета — определяется присутствием в сетчатке всех трёх зрительных пигментов (для красного, зелёного и синего — первичные цвета). Эти основы теории цветного зрения предложил Томас Янг (1802) и развил Герман Гельмгольц Дихромазии (цветовая слепота, или дальтонизм)— дефекты цветового восприятия (преимущественно у мужчин; например, в Европе разные дефекты у мужчин составляют 8% общей популяции) по одному из первичных цветов — подразделяют на протанопии, дейтанопии и тританопии (от греч. первый, второй и третий [имеются в виду порядковые номера первичных цветов: соответственно красный, зелёный, синий]).
 развивается при связанном с") Дихромазии Протанопия (страдает восприятие красного, примерно 25% случаев цветовой слепоты) развивается при связанном с хромосомой Х наследовании генного дефекта. Дейтанопия (цветовая слепота по восприятию зелёного, около 75% всех случаев; связанное с хромосомой Х наследование, полиморфизм гена). Тританопия (страдает преимущественно восприятие фиолетового цвета, дефектное зрение по синему и жёлтому). Аутосомное доминантное наследование дефектного гена. Полная цветовая слепота — ахромазия — встречается крайне редко. При ахромазии поражается весь колбочковый аппарат, и человек видит все предметы в разных оттенках серого.
Дихромазии Протанопия (страдает восприятие красного, примерно 25% случаев цветовой слепоты) развивается при связанном с хромосомой Х наследовании генного дефекта. Дейтанопия (цветовая слепота по восприятию зелёного, около 75% всех случаев; связанное с хромосомой Х наследование, полиморфизм гена). Тританопия (страдает преимущественно восприятие фиолетового цвета, дефектное зрение по синему и жёлтому). Аутосомное доминантное наследование дефектного гена. Полная цветовая слепота — ахромазия — встречается крайне редко. При ахромазии поражается весь колбочковый аппарат, и человек видит все предметы в разных оттенках серого.
 Орган слуха
Орган слуха
 Орган слуха анатомически состоит из наружного, среднего и внутреннего уха
Орган слуха анатомически состоит из наружного, среднего и внутреннего уха
 Наружное ухо Представлено ушной раковиной и наружным слуховым проходом. Ушная раковина — эластический хрящ сложной формы, покрытый кожей, на дне которого находится наружное слуховое отверстие. Форма ушной раковины способствует тому, чтобы направлять звук в наружный слуховой проход. Некоторые люди могут двигать ушами при помощи слабых мышц, прикреплённых к черепу. Наружный слуховой проход — слепая трубка длиной 2, 5 см, заканчивающаяся у барабанной перепонки. Наружная треть прохода состоит из хряща и покрыта тонкими защитными волосами. Внутренние части прохода находятся в височной кости и содержат модифицированные потовые железы — церуминозные железы, которые производят восковидный секрет — ушную серу — для защиты кожи прохода и фиксации пыли и бактерий.
Наружное ухо Представлено ушной раковиной и наружным слуховым проходом. Ушная раковина — эластический хрящ сложной формы, покрытый кожей, на дне которого находится наружное слуховое отверстие. Форма ушной раковины способствует тому, чтобы направлять звук в наружный слуховой проход. Некоторые люди могут двигать ушами при помощи слабых мышц, прикреплённых к черепу. Наружный слуховой проход — слепая трубка длиной 2, 5 см, заканчивающаяся у барабанной перепонки. Наружная треть прохода состоит из хряща и покрыта тонкими защитными волосами. Внутренние части прохода находятся в височной кости и содержат модифицированные потовые железы — церуминозные железы, которые производят восковидный секрет — ушную серу — для защиты кожи прохода и фиксации пыли и бактерий.
 трубы и отделена") Среднее ухо Его полость сообщается с носоглоткой при помощи евстахиевой (слуховой) трубы и отделена от наружного слухового прохода барабанной перепонкой диаметром 9 мм, а от преддверия и барабанной лестницы улитки — овальным и круглым окнами соответственно. Барабанная перепонка передаёт звуковые колебания на три маленькие взаимосвязанные слуховые косточки: молоточек прикреплён к барабанной перепонке, а стремечко — к овальному окну. Эти косточки вибрируют в унисон и усиливают звук в двадцать раз. Слуховая труба поддерживает давление воздуха в полости среднего уха на уровне атмосферного.
Среднее ухо Его полость сообщается с носоглоткой при помощи евстахиевой (слуховой) трубы и отделена от наружного слухового прохода барабанной перепонкой диаметром 9 мм, а от преддверия и барабанной лестницы улитки — овальным и круглым окнами соответственно. Барабанная перепонка передаёт звуковые колебания на три маленькие взаимосвязанные слуховые косточки: молоточек прикреплён к барабанной перепонке, а стремечко — к овальному окну. Эти косточки вибрируют в унисон и усиливают звук в двадцать раз. Слуховая труба поддерживает давление воздуха в полости среднего уха на уровне атмосферного.
 Внутреннее ухо Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся в перилимфе полукружные каналы, маточка, мешочек и улитковый проток (перепончатый канал улитки) — эндолимфой. Между эндолимфой и перилимфой существует электрический потенциал — около +80 м. В (внутриулитковый, или эндокохлеарный потенциал). Эндолимфа — вязкая жидкость, заполняет перепончатый канал улитки и соединяется через специальный канал (ductus reuniens) с эндолимфой вестибулярного аппарата. Концентрация K+ в эндолимфе в 100 раз больше, чем в спинномозговой жидкости (ликворе) и перилимфе; концентрация Na+ в эндолимфе в 10 раз меньше, чем в перилимфе. Перилимфа по химическому составу близка к плазме крови и ликвору и занимает промежуточное положение между ними по содержанию белка. Эндокохлеарный потенциал. Перепончатый канал улитки заряжен положительно (+60–+80 м. В) относительно двух других лестниц. Источник этого (эндокохлеарного) потенциала — сосудистая полоска. Волосковые клетки поляризованы эндокохлеарным потенциалом до критического уровня, что повышает их чувствительность к механическому воздействию.
Внутреннее ухо Полость преддверия, барабанная и вестибулярная лестницы улитки заполнены перилимфой, а находящиеся в перилимфе полукружные каналы, маточка, мешочек и улитковый проток (перепончатый канал улитки) — эндолимфой. Между эндолимфой и перилимфой существует электрический потенциал — около +80 м. В (внутриулитковый, или эндокохлеарный потенциал). Эндолимфа — вязкая жидкость, заполняет перепончатый канал улитки и соединяется через специальный канал (ductus reuniens) с эндолимфой вестибулярного аппарата. Концентрация K+ в эндолимфе в 100 раз больше, чем в спинномозговой жидкости (ликворе) и перилимфе; концентрация Na+ в эндолимфе в 10 раз меньше, чем в перилимфе. Перилимфа по химическому составу близка к плазме крови и ликвору и занимает промежуточное положение между ними по содержанию белка. Эндокохлеарный потенциал. Перепончатый канал улитки заряжен положительно (+60–+80 м. В) относительно двух других лестниц. Источник этого (эндокохлеарного) потенциала — сосудистая полоска. Волосковые клетки поляризованы эндокохлеарным потенциалом до критического уровня, что повышает их чувствительность к механическому воздействию.
 орган Канал улитки разделён на барабанную и вестибулярную лестницы") Перепончатый канал и спиральный (кортиев) орган Канал улитки разделён на барабанную и вестибулярную лестницы и перепончатый канал (средняя лестница), в котором расположен кортиев орган. Перепончатый канал отделён от барабанной лестницы базилярной мембраной. В её составе проходят периферические отростки нейронов спирального ганглия, образующие синаптические контакты с наружными и внутренними волосковыми клетками
Перепончатый канал и спиральный (кортиев) орган Канал улитки разделён на барабанную и вестибулярную лестницы и перепончатый канал (средняя лестница), в котором расположен кортиев орган. Перепончатый канал отделён от барабанной лестницы базилярной мембраной. В её составе проходят периферические отростки нейронов спирального ганглия, образующие синаптические контакты с наружными и внутренними волосковыми клетками
 с увеличенной частью спираль ного (кортиева) органа (Б) 1") Поперечный разрез завитка улитки (А) с увеличенной частью спираль ного (кортиева) органа (Б) 1 — секреторный эпителий; 2 — покровная мембрана; 3 — спиральный (кортиев) орган; 4 — спиральная связка; 5 — барабанная лестница; 6 — слой миелина; 7 — спиральный ганглий; 9 — спиральная артерия; 10 — преддверная мембрана Рейсснера; 11— лестница преддверия; 12 — перепончатый канал; 13 — основная мембрана; 14 — внутренняя спи ральная борозда; 15 — наружные волосковые клетки; 16 — решетчатая пластинка; 17 — нервные волокна, идущие к волосковым клеткам; 18 — внутренняя волосковая клетка.
Поперечный разрез завитка улитки (А) с увеличенной частью спираль ного (кортиева) органа (Б) 1 — секреторный эпителий; 2 — покровная мембрана; 3 — спиральный (кортиев) орган; 4 — спиральная связка; 5 — барабанная лестница; 6 — слой миелина; 7 — спиральный ганглий; 9 — спиральная артерия; 10 — преддверная мембрана Рейсснера; 11— лестница преддверия; 12 — перепончатый канал; 13 — основная мембрана; 14 — внутренняя спи ральная борозда; 15 — наружные волосковые клетки; 16 — решетчатая пластинка; 17 — нервные волокна, идущие к волосковым клеткам; 18 — внутренняя волосковая клетка.
 Проведение звука к улитке Цепочка передачи звукового давления выглядит следующим образом: барабанная перепонка молоточек наковальня стремя мембрана овального окна перилимфа базилярная и текториальная мембраны мембрана круглого окна. При смещении стремени перилимфа перемещается по вестибулярной лестнице и затем через геликотрему по барабанной лестнице к круглому окну. Жидкость, сдвинутая смещением мембраны овального окна, создаёт избыточное давление в вестибулярном канале. Под действием этого давления базилярная мембрана смещается в сторону барабанной лестницы. Колебательная реакция в виде волны распространяется от базилярной мембраны к геликотреме. Смещение текториальной мембраны относительно волосковых клеток при действии звука вызывает их возбуждение. Возникающая электрическая реакция (микрофонный эффект) повторяет форму звукового сигнала
Проведение звука к улитке Цепочка передачи звукового давления выглядит следующим образом: барабанная перепонка молоточек наковальня стремя мембрана овального окна перилимфа базилярная и текториальная мембраны мембрана круглого окна. При смещении стремени перилимфа перемещается по вестибулярной лестнице и затем через геликотрему по барабанной лестнице к круглому окну. Жидкость, сдвинутая смещением мембраны овального окна, создаёт избыточное давление в вестибулярном канале. Под действием этого давления базилярная мембрана смещается в сторону барабанной лестницы. Колебательная реакция в виде волны распространяется от базилярной мембраны к геликотреме. Смещение текториальной мембраны относительно волосковых клеток при действии звука вызывает их возбуждение. Возникающая электрическая реакция (микрофонный эффект) повторяет форму звукового сигнала
 Слуховые косточки Звук колеблет барабанную перепонку и передаёт энергию колебаний по системе слуховых косточек перилимфе вестибулярной лестницы. Если бы не существовало барабанной перепонки и слуховых косточек, звук мог бы достигать внутреннего уха, но значительная часть звуковой энергии отражалась бы обратно из за разницы акустических сопротивлений (импедансов) воздушной и жидкой сред. Поэтому важнейшая роль барабанной перепонки и цепи слуховых косточек заключается в создании соответствия между импедансами внешней воздушной среды и жидкой среды внутреннего уха. Амплитуда движений подошвы стремени во время каждого звукового колебания составляет всего лишь три четверти от амплитуды колебаний рукоятки молоточка. Следовательно, колебательная рычажная система косточек не увеличивает размах движений стремечка. Вместо этого рычажная система уменьшает размах колебаний, но увеличивает их силу примерно в 1, 3 раза. К этому следует добавить, что площадь барабанной перепонки составляет 55 мм 2, в то время как площадь подошвы стремени равна 3, 2 мм 2. Разница в системе рычагов в 17 раз приводит к тому, что давление на жидкость в улитке в 22 раза выше, чем давление воздуха на барабанную перепонку. Выравнивание импедансов между звуковыми волнами и звуковыми колебаниями жидкости улучшает чёткость восприятия звуковых частот в пределах от 300 до 3000 Гц.
Слуховые косточки Звук колеблет барабанную перепонку и передаёт энергию колебаний по системе слуховых косточек перилимфе вестибулярной лестницы. Если бы не существовало барабанной перепонки и слуховых косточек, звук мог бы достигать внутреннего уха, но значительная часть звуковой энергии отражалась бы обратно из за разницы акустических сопротивлений (импедансов) воздушной и жидкой сред. Поэтому важнейшая роль барабанной перепонки и цепи слуховых косточек заключается в создании соответствия между импедансами внешней воздушной среды и жидкой среды внутреннего уха. Амплитуда движений подошвы стремени во время каждого звукового колебания составляет всего лишь три четверти от амплитуды колебаний рукоятки молоточка. Следовательно, колебательная рычажная система косточек не увеличивает размах движений стремечка. Вместо этого рычажная система уменьшает размах колебаний, но увеличивает их силу примерно в 1, 3 раза. К этому следует добавить, что площадь барабанной перепонки составляет 55 мм 2, в то время как площадь подошвы стремени равна 3, 2 мм 2. Разница в системе рычагов в 17 раз приводит к тому, что давление на жидкость в улитке в 22 раза выше, чем давление воздуха на барабанную перепонку. Выравнивание импедансов между звуковыми волнами и звуковыми колебаниями жидкости улучшает чёткость восприятия звуковых частот в пределах от 300 до 3000 Гц.
 Звукопонижающий рефлекс Мышцы среднего уха. Функциональная роль мышц среднего уха заключается в уменьшении воздействия громких звуков на слуховую систему. При действии громких звуков на передающую систему и поступлении сигналов в ЦНС через 40– 80 мсек возникает звукопонижающий рефлекс, вызывающий сокращение мышц, прикреплённых к стремечку и молоточку. Мышца молоточка тянет ручку молоточка вперёд и вниз, а мышца стремечка тянет стремечко наружу и вверх. Эти две противоположно направленные силы увеличивают ригидность рычаговой системы косточек, уменьшая проведение низкочастотных звуков, особенно звуков частотой ниже 1000 Гц. Звукопонижающий рефлекс может уменьшать интенсивность передачи низкочастотных звуков на 30– 40 д. Б, в то же самое время не затрагивая восприятия громкого голоса и шепотной речи. Значение этого рефлекторного механизма двояко: защита улитки от повреждающего вибрационного действия низкого звука и маскировка низких звуков в окружающей среде. Кроме того, мышцы слуховых косточек уменьшают чувствительность слуха человека к его собственной речи в момент, когда мозг активирует голосовой механизм.
Звукопонижающий рефлекс Мышцы среднего уха. Функциональная роль мышц среднего уха заключается в уменьшении воздействия громких звуков на слуховую систему. При действии громких звуков на передающую систему и поступлении сигналов в ЦНС через 40– 80 мсек возникает звукопонижающий рефлекс, вызывающий сокращение мышц, прикреплённых к стремечку и молоточку. Мышца молоточка тянет ручку молоточка вперёд и вниз, а мышца стремечка тянет стремечко наружу и вверх. Эти две противоположно направленные силы увеличивают ригидность рычаговой системы косточек, уменьшая проведение низкочастотных звуков, особенно звуков частотой ниже 1000 Гц. Звукопонижающий рефлекс может уменьшать интенсивность передачи низкочастотных звуков на 30– 40 д. Б, в то же самое время не затрагивая восприятия громкого голоса и шепотной речи. Значение этого рефлекторного механизма двояко: защита улитки от повреждающего вибрационного действия низкого звука и маскировка низких звуков в окружающей среде. Кроме того, мышцы слуховых косточек уменьшают чувствительность слуха человека к его собственной речи в момент, когда мозг активирует голосовой механизм.
 Механизмы слуховой рецепции При действии звука основная мембрана начинает колебаться, наиболее длинные волоски рецепторных клеток (стереоцилии) касаются покровной мембраны и несколько наклоняются. Отклонение волоска на несколько градусов приводит к натяжению тончайших вертикальных нитей (микрофиламент), связывающих между собой верхушки соседних волосков данной клетки. Это натяжение чисто механически открывает от 1 до 5 ионных каналов в мембране стереоцилии. Через открытый канал в волосок начинает течь калиевый ионный ток. Сила натяжения нити, необходимая для открывания одного канала, ничтожна, около 2 • 10 13 ньютонов.
Механизмы слуховой рецепции При действии звука основная мембрана начинает колебаться, наиболее длинные волоски рецепторных клеток (стереоцилии) касаются покровной мембраны и несколько наклоняются. Отклонение волоска на несколько градусов приводит к натяжению тончайших вертикальных нитей (микрофиламент), связывающих между собой верхушки соседних волосков данной клетки. Это натяжение чисто механически открывает от 1 до 5 ионных каналов в мембране стереоцилии. Через открытый канал в волосок начинает течь калиевый ионный ток. Сила натяжения нити, необходимая для открывания одного канала, ничтожна, около 2 • 10 13 ньютонов.
 Основные слуховые пути
Основные слуховые пути
 Слуховые пути и центры Афферентные нервные волокна от улитки входят в спиральный ганглий и от него поступают в дорсальные (задние) и вентральные (передние) улитковые ядра, расположенные в верхней части продолговатого мозга. Здесь восходящие нервные волокна образуют синапсы с нейронами второго порядка, аксоны которых частью переходят на противоположную сторону к ядрам верхней оливы, а частью оканчиваются на ядрах верхней оливы этой же стороны. От ядер верхней оливы слуховые пути поднимаются вверх через латеральный лемнисковой путь; часть волокон оканчивается в латеральных лемнисковых ядрах, а большинство аксонов минует эти ядра и следует до нижнего двухолмия, где все или почти все слуховые волокна образуют синапсы. Отсюда слуховой путь проходит к медиальным коленчатым телам, где все волокна заканчиваются синапсами. Окончательно слуховой путь завершается в слуховой коре, располагающейся главным образом в верхней извилине височной доли. Базилярная мембрана улитки на всех уровнях слухового пути представлена в форме определённых проекционных карт различных частот. Уже на уровне среднего мозга появляются нейроны, детектирующие на принципах латерального и возвратного торможения несколько признаков звука.
Слуховые пути и центры Афферентные нервные волокна от улитки входят в спиральный ганглий и от него поступают в дорсальные (задние) и вентральные (передние) улитковые ядра, расположенные в верхней части продолговатого мозга. Здесь восходящие нервные волокна образуют синапсы с нейронами второго порядка, аксоны которых частью переходят на противоположную сторону к ядрам верхней оливы, а частью оканчиваются на ядрах верхней оливы этой же стороны. От ядер верхней оливы слуховые пути поднимаются вверх через латеральный лемнисковой путь; часть волокон оканчивается в латеральных лемнисковых ядрах, а большинство аксонов минует эти ядра и следует до нижнего двухолмия, где все или почти все слуховые волокна образуют синапсы. Отсюда слуховой путь проходит к медиальным коленчатым телам, где все волокна заканчиваются синапсами. Окончательно слуховой путь завершается в слуховой коре, располагающейся главным образом в верхней извилине височной доли. Базилярная мембрана улитки на всех уровнях слухового пути представлена в форме определённых проекционных карт различных частот. Уже на уровне среднего мозга появляются нейроны, детектирующие на принципах латерального и возвратного торможения несколько признаков звука.
 Слуховая кора Проекционные области слуховой коры располагаются не только в верхней части верхней височной извилины, но и простираются на наружную сторону височной доли, захватывая часть островковой коры и теменной покрышки. Первичная слуховая кора непосредственно получает сигналы от внутреннего (медиального) коленчатого тела, в то время как слуховая ассоциативная область вторично возбуждается импульсами из первичной слуховой коры и таламических областей, граничащих с медиальным коленчатым телом
Слуховая кора Проекционные области слуховой коры располагаются не только в верхней части верхней височной извилины, но и простираются на наружную сторону височной доли, захватывая часть островковой коры и теменной покрышки. Первичная слуховая кора непосредственно получает сигналы от внутреннего (медиального) коленчатого тела, в то время как слуховая ассоциативная область вторично возбуждается импульсами из первичной слуховой коры и таламических областей, граничащих с медиальным коленчатым телом
 Тонотопические карты В каждой из 6 тонотопических карт звуки высокой частоты возбуждают нейроны в задней части карты, в то время как звуки низкой частоты возбуждают нейроны в передней её части. Предполагают, что каждая отдельная область воспринимает свои специфические особенности звука. Например, одна большая карта в первичной слуховой коре почти целиком дискриминирует звуки, которые субъекту кажутся высокими. Другая карта используется для определения направления поступления звука. Некоторые области слуховой коры выявляют специальные качества звуковых сигналов (например, неожиданное начало звуков или модуляции звуков). Диапазон звуковой частоты, на которую отвечают нейроны слуховой коры уже, чем для нейронов спирального ганглия и мозгового ствола. Это объясняется, с одной стороны, высокой степенью специализации нейронов коры, а с другой стороны — феном латерального и возвратного торможения, усиливающего разрешающую способность нейронов воспринимать необходимую частоту звука. Многие нейроны слуховой коры, особенно в слуховой ассоциативной коре, отвечают не только на специфические звуковые частоты. Эти нейроны «ассоциируют» звуковые частоты с другими видами сенсорной информации. В самом деле, теменная часть слуховой ассоциативной коры перекрывает соматосенсорную область II, что создаёт возможность ассоциации слуховой информации с соматосенсорной информацией
Тонотопические карты В каждой из 6 тонотопических карт звуки высокой частоты возбуждают нейроны в задней части карты, в то время как звуки низкой частоты возбуждают нейроны в передней её части. Предполагают, что каждая отдельная область воспринимает свои специфические особенности звука. Например, одна большая карта в первичной слуховой коре почти целиком дискриминирует звуки, которые субъекту кажутся высокими. Другая карта используется для определения направления поступления звука. Некоторые области слуховой коры выявляют специальные качества звуковых сигналов (например, неожиданное начало звуков или модуляции звуков). Диапазон звуковой частоты, на которую отвечают нейроны слуховой коры уже, чем для нейронов спирального ганглия и мозгового ствола. Это объясняется, с одной стороны, высокой степенью специализации нейронов коры, а с другой стороны — феном латерального и возвратного торможения, усиливающего разрешающую способность нейронов воспринимать необходимую частоту звука. Многие нейроны слуховой коры, особенно в слуховой ассоциативной коре, отвечают не только на специфические звуковые частоты. Эти нейроны «ассоциируют» звуковые частоты с другими видами сенсорной информации. В самом деле, теменная часть слуховой ассоциативной коры перекрывает соматосенсорную область II, что создаёт возможность ассоциации слуховой информации с соматосенсорной информацией
 Слуховые ощущения Диапазон частот, который воспринимает человек, включает около 10 октав музыкальной шкалы (от 16 Гц до 20 к. Гц). Этот диапазон постепенно уменьшается с возрастом за счёт снижения восприятия высоких частот. Различение частоты звука характеризуется минимальным различием по частоте двух близких звуков, которое ещё улавливается человеком. Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек в 50% случаев его предъявления. Порог слышимости зависит от частоты звуковых волн. Максимальная чувствительность слуха человека располагается в области от 500 до 4000 Гц. В этих границах воспринимается звук, имеющий чрезвычайно малую энергию. В диапазоне этих частот располагается область звукового восприятия речи человека. Чувствительность к звуковым частотам ниже 500 Гц прогрессивно снижается. Это предохраняет человека от возможного постоянного ощущения низкочастотных колебаний и шумов, производимых собственным телом.
Слуховые ощущения Диапазон частот, который воспринимает человек, включает около 10 октав музыкальной шкалы (от 16 Гц до 20 к. Гц). Этот диапазон постепенно уменьшается с возрастом за счёт снижения восприятия высоких частот. Различение частоты звука характеризуется минимальным различием по частоте двух близких звуков, которое ещё улавливается человеком. Абсолютный порог слуховой чувствительности — минимальная сила звука, которую слышит человек в 50% случаев его предъявления. Порог слышимости зависит от частоты звуковых волн. Максимальная чувствительность слуха человека располагается в области от 500 до 4000 Гц. В этих границах воспринимается звук, имеющий чрезвычайно малую энергию. В диапазоне этих частот располагается область звукового восприятия речи человека. Чувствительность к звуковым частотам ниже 500 Гц прогрессивно снижается. Это предохраняет человека от возможного постоянного ощущения низкочастотных колебаний и шумов, производимых собственным телом.
 Обонятельный анализатор
Обонятельный анализатор
 Топография обонятельной выстилки, обонятельной луковицы и обонятельного тракта
Топография обонятельной выстилки, обонятельной луковицы и обонятельного тракта
 образуют в") Обонятельная слизистая оболочка Воспринимающие обонятельные структуры (периферическая часть обонятельного анализатора, орган обоняния) образуют в слизистой оболочке носа специализированную область — парную обонятельную выстилку (обонятельное поле), расположенную под решётчатой пластинкой. Суммарная площадь обонятельного поля (обонятельного эпителия) менее 10 см 2
Обонятельная слизистая оболочка Воспринимающие обонятельные структуры (периферическая часть обонятельного анализатора, орган обоняния) образуют в слизистой оболочке носа специализированную область — парную обонятельную выстилку (обонятельное поле), расположенную под решётчатой пластинкой. Суммарная площадь обонятельного поля (обонятельного эпителия) менее 10 см 2
 Орган обоняния эпителии обонятельной выстилки расположены обонятельные рецепторные нейроны. Их наружные отростки заканчиваются обонятельными булавами, от которых в разные стороны параллельно поверхности эпителия отходят обонятельные волоски. Аксоны рецепторных нейронов в составе обонятельных нитей (fila olfactoria) обонятельного нерва проходят через lamina cribrosa решётчатой кости и образуют синапсы с нейронами обонятельной луковицы (bulbus olfactorius). Многочисленные опорные клетки окружают рецепторные клетки
Орган обоняния эпителии обонятельной выстилки расположены обонятельные рецепторные нейроны. Их наружные отростки заканчиваются обонятельными булавами, от которых в разные стороны параллельно поверхности эпителия отходят обонятельные волоски. Аксоны рецепторных нейронов в составе обонятельных нитей (fila olfactoria) обонятельного нерва проходят через lamina cribrosa решётчатой кости и образуют синапсы с нейронами обонятельной луковицы (bulbus olfactorius). Многочисленные опорные клетки окружают рецепторные клетки
 Строение обонятельного эпителия
Строение обонятельного эпителия
 Архитектоника нейронов и связей в обонятельной луковице ПК — пучковая клетка, МК — митральная клетка с возвратными коллатералями (К), ЗК — зернистая клетка, ВПЯ — нисходящие в обонятельную луковицу нервные волокна из переднего обонятельного ядра, ВПС — комиссуральные нервные волокна передней спайки мозолистого тела (commissura anterior). Прерывистые линии — границы между анатомическими структурами.
Архитектоника нейронов и связей в обонятельной луковице ПК — пучковая клетка, МК — митральная клетка с возвратными коллатералями (К), ЗК — зернистая клетка, ВПЯ — нисходящие в обонятельную луковицу нервные волокна из переднего обонятельного ядра, ВПС — комиссуральные нервные волокна передней спайки мозолистого тела (commissura anterior). Прерывистые линии — границы между анатомическими структурами.
") Восходящие пути и обонятельный мозг Обонятельные нервы (nn. olfactorii, пучки аксонов рецепторных обонятельных клеток) в виде 18 20 тонких ветвей вступают в обонятельную луковицу (bulbus olfactorius). Последняя переходит в обонятельный тракт (tractus olfactorius, аксоны митральных и пучковых клеток), заканчивающиеся в первичных обонятельных центрах (обонятельный треугольник – trigonum olfactorium, переднее продырявленное вещество – substancia perforata anterior, прозрачная перегородка – septum pellucidum). От первичных обонятельных центров сигналы направляются к корковым центрам обоняния своей (ипсилатеральной) и противоположной (контрлатеральной) стороны: извилине гиппокампа (gyrus hippocampi) и крючку (uncus). Обонятельный мозг (rhinencephalon) образуют обонятельная луковица (bulbus olfactorius), обонятельный тракт (tractus olfactorius), обонятельный треугольник (trigonum olfactorium), переднее продырявленное вещество (substancia perforata anterior), гиппокампока борозда, зубчатая извилина и ряд прилежащих структур.
Восходящие пути и обонятельный мозг Обонятельные нервы (nn. olfactorii, пучки аксонов рецепторных обонятельных клеток) в виде 18 20 тонких ветвей вступают в обонятельную луковицу (bulbus olfactorius). Последняя переходит в обонятельный тракт (tractus olfactorius, аксоны митральных и пучковых клеток), заканчивающиеся в первичных обонятельных центрах (обонятельный треугольник – trigonum olfactorium, переднее продырявленное вещество – substancia perforata anterior, прозрачная перегородка – septum pellucidum). От первичных обонятельных центров сигналы направляются к корковым центрам обоняния своей (ипсилатеральной) и противоположной (контрлатеральной) стороны: извилине гиппокампа (gyrus hippocampi) и крючку (uncus). Обонятельный мозг (rhinencephalon) образуют обонятельная луковица (bulbus olfactorius), обонятельный тракт (tractus olfactorius), обонятельный треугольник (trigonum olfactorium), переднее продырявленное вещество (substancia perforata anterior), гиппокампока борозда, зубчатая извилина и ряд прилежащих структур.
 Обонятельный тракт и обонятельный мозг Аксоны митральных клеток в составе обонятельного тракта поднимаются в первичные обонятельные центры. В составе обонятельного мозга различают медиальную и латеральную обонятельные области. Нейроны медиальной обонятельной области (ядра перегородки) проецируются в гипоталамус и другие области, контролирующие поведение. Аксоны нервных клеток латеральной обонятельной области, расположенных в грушевидной коре и миндалевидному телу, направляются к гиппокампу. Обонятельные стимулы активируют грушевидную кору билатерально. Наконец, существуют гомолатеральные (только односторонние и той же стороны) проекции к дорсомедиальному ядру таламуса и далее — к обонятельной борозде.
Обонятельный тракт и обонятельный мозг Аксоны митральных клеток в составе обонятельного тракта поднимаются в первичные обонятельные центры. В составе обонятельного мозга различают медиальную и латеральную обонятельные области. Нейроны медиальной обонятельной области (ядра перегородки) проецируются в гипоталамус и другие области, контролирующие поведение. Аксоны нервных клеток латеральной обонятельной области, расположенных в грушевидной коре и миндалевидному телу, направляются к гиппокампу. Обонятельные стимулы активируют грушевидную кору билатерально. Наконец, существуют гомолатеральные (только односторонние и той же стороны) проекции к дорсомедиальному ядру таламуса и далее — к обонятельной борозде.
 РЕГИСТРАЦИЯ И ПРЕОБРАЗОВАНИЕ ОБОНЯТЕЛЬНОГО СИГНАЛА Рецепторные клетки обонятельной выстилки регистрируют небольшое количество первичных запахов, но их комбинации формируют ощущение многих воспринимаемых запахов. Возникает вопрос: каким образом обонятельная система может различать множество различных запахов? Варианты ответа таковы: или рецепторные обонятельные нейроны имеют множество различных молекулярных рецепторов (в том числе и каждый нейрон), или/и содержат один или несколько типов молекулярных обонятельных рецепторов, но посылают в ЦНС различные спектры ПД. Наконец, формирование ощущения конкретного запаха может зависеть от фиксированных связей между нервными клетками обонятельной системы
РЕГИСТРАЦИЯ И ПРЕОБРАЗОВАНИЕ ОБОНЯТЕЛЬНОГО СИГНАЛА Рецепторные клетки обонятельной выстилки регистрируют небольшое количество первичных запахов, но их комбинации формируют ощущение многих воспринимаемых запахов. Возникает вопрос: каким образом обонятельная система может различать множество различных запахов? Варианты ответа таковы: или рецепторные обонятельные нейроны имеют множество различных молекулярных рецепторов (в том числе и каждый нейрон), или/и содержат один или несколько типов молекулярных обонятельных рецепторов, но посылают в ЦНС различные спектры ПД. Наконец, формирование ощущения конкретного запаха может зависеть от фиксированных связей между нервными клетками обонятельной системы
 Механизмы трансдукции сигнала ОБОНЯТЕЛЬНЫЕ РЕЦЕПТОРНЫЕ БЕЛКИ кодируют гены, расположенные практически во всех хромосомах, кроме аутосомы 20 и половой хромосомы Y. В геноме человека идентифицировано более 900 генов, кодирующих белки обонятельных рецепторов, что практически равно одной тридцатой всего генома. Эти белки относят к семейству связанных с G‑белком рецепторов.
Механизмы трансдукции сигнала ОБОНЯТЕЛЬНЫЕ РЕЦЕПТОРНЫЕ БЕЛКИ кодируют гены, расположенные практически во всех хромосомах, кроме аутосомы 20 и половой хромосомы Y. В геноме человека идентифицировано более 900 генов, кодирующих белки обонятельных рецепторов, что практически равно одной тридцатой всего генома. Эти белки относят к семейству связанных с G‑белком рецепторов.
 ПОСЛЕДОВАТЕЛЬНОСТЬ СОБЫТИЙ при регистрации обонятельного сигнала может быть представлена следующим образом: ◦ взаимодействие пахучего вещества с белком рецептором в плазмолемме обонятельных волосков ◦ активация G‑белка ◦ повышение активности аденилатциклазы ◦ увеличение уровня ц. АМФ ◦ активация ц. АМФ–зависимых воротных катионных каналов ◦ деполяризация рецепторных нейронов ◦ генерация ПД и его проведение по аксону.
ПОСЛЕДОВАТЕЛЬНОСТЬ СОБЫТИЙ при регистрации обонятельного сигнала может быть представлена следующим образом: ◦ взаимодействие пахучего вещества с белком рецептором в плазмолемме обонятельных волосков ◦ активация G‑белка ◦ повышение активности аденилатциклазы ◦ увеличение уровня ц. АМФ ◦ активация ц. АМФ–зависимых воротных катионных каналов ◦ деполяризация рецепторных нейронов ◦ генерация ПД и его проведение по аксону.
 Трансформация сигнала в обонятельном рецепторе А — рецепторный обонятельный нейрон; Б — вход Na+ в клетку; В — пахучие молекулы соединяются с рецептором (R). Рецептор активирует G‑белок (G), G‑белок активирует аденилатциклазу (Ац), образующийся ц. АМФ открывает Na+‑каналы.
Трансформация сигнала в обонятельном рецепторе А — рецепторный обонятельный нейрон; Б — вход Na+ в клетку; В — пахучие молекулы соединяются с рецептором (R). Рецептор активирует G‑белок (G), G‑белок активирует аденилатциклазу (Ац), образующийся ц. АМФ открывает Na+‑каналы.
 ОБОНЯНИЕ, ПОЛОВОЕ ПОВЕДЕНИЕ И ПАМЯТЬ Между запахом и половой функцией у многих видов животных существует тесная взаимосвязь (скорее всего, реализуемая через cошниково–носовой [вомероназальный] орган Якобсона — участок слизистой оболочки носовых ходов, аналогичный обонятельной выстилке), а использование духов даёт достаточные основания считать, что подобная взаимосвязь существует и у людей (орган Якобсона у человека отсутствует). Обоняние у женщин развито сильнее, и оно обостряется ещё больше в период овуляции. Запах и (в меньшей степени) вкус обладают уникальной способностью оживлять воспоминания, заложенные в долговременной памяти. Этот факт отмечен писателями и экспериментально доказан психологами
ОБОНЯНИЕ, ПОЛОВОЕ ПОВЕДЕНИЕ И ПАМЯТЬ Между запахом и половой функцией у многих видов животных существует тесная взаимосвязь (скорее всего, реализуемая через cошниково–носовой [вомероназальный] орган Якобсона — участок слизистой оболочки носовых ходов, аналогичный обонятельной выстилке), а использование духов даёт достаточные основания считать, что подобная взаимосвязь существует и у людей (орган Якобсона у человека отсутствует). Обоняние у женщин развито сильнее, и оно обостряется ещё больше в период овуляции. Запах и (в меньшей степени) вкус обладают уникальной способностью оживлять воспоминания, заложенные в долговременной памяти. Этот факт отмечен писателями и экспериментально доказан психологами
 имеет уникальный") КОМБИНАТОРНОЕ КОДИРОВАНИЕ Эта концепция подразумевает, что каждый из миллионов отдельных запахов (одорантов) имеет уникальный код; кодирование происходит на уровне органа обоняния, а декодирование — в обонятельных центрах. В пользу существования такого обонятельного кода свидетельствуют следующие факты. Каждый обонятельный нейрон экспрессирует один тип обонятельных рецепторов. Конкретный тип обонятельного рецептора распознаёт несколько одорантов (в среднем четыре). Конкретная молекула одоранта может активировать несколько разных типов обонятельных рецепторов. Минимальные изменения конформации одоранта, его химической структуры, а также изменения концентрация молекулы пахучего вещества изменяют код одоранта. Последнее обстоятельство позволило объяснить феномен формирования ощущения разных запахов при вдыхании разных концентраций одного и того же одоранта.
КОМБИНАТОРНОЕ КОДИРОВАНИЕ Эта концепция подразумевает, что каждый из миллионов отдельных запахов (одорантов) имеет уникальный код; кодирование происходит на уровне органа обоняния, а декодирование — в обонятельных центрах. В пользу существования такого обонятельного кода свидетельствуют следующие факты. Каждый обонятельный нейрон экспрессирует один тип обонятельных рецепторов. Конкретный тип обонятельного рецептора распознаёт несколько одорантов (в среднем четыре). Конкретная молекула одоранта может активировать несколько разных типов обонятельных рецепторов. Минимальные изменения конформации одоранта, его химической структуры, а также изменения концентрация молекулы пахучего вещества изменяют код одоранта. Последнее обстоятельство позволило объяснить феномен формирования ощущения разных запахов при вдыхании разных концентраций одного и того же одоранта.
, гипосмию (уменьшение обонятельной чувствительности), дизосмию (искажение чувства") НАРУШЕНИЯ ОБОНЯНИЯ включают аносмию (отсутствие чувства запаха), гипосмию (уменьшение обонятельной чувствительности), дизосмию (искажение чувства обоняния), гиперосмию (усиление обоняния). У человека известно несколько видов аносмий, касающихся отсутствия восприятия запаха для какого нибудь одного из членов семейства обонятельных рецепторов. Пороги восприятия запахов увеличиваются с возрастом. Примерно 75% лиц старше 80 лет имеют нарушенное восприятие запахов.
НАРУШЕНИЯ ОБОНЯНИЯ включают аносмию (отсутствие чувства запаха), гипосмию (уменьшение обонятельной чувствительности), дизосмию (искажение чувства обоняния), гиперосмию (усиление обоняния). У человека известно несколько видов аносмий, касающихся отсутствия восприятия запаха для какого нибудь одного из членов семейства обонятельных рецепторов. Пороги восприятия запахов увеличиваются с возрастом. Примерно 75% лиц старше 80 лет имеют нарушенное восприятие запахов.
 Вкусовой анализатор
Вкусовой анализатор
 ВКУС
ВКУС
 Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол мозга к таламусу и проецируются вдоль постцентральной извилины. Периферическая часть вкусового анализатора — вкусовые почки — расположены в слизистой оболочке полости рта, переднего отдела глотки, пищевода и гортани. Основная масса вкусовых почек (>90% их общего количества — до 10 тыс. ) находится в хемочувствительных сосочках языка — листовидных, грибовидных и желобоватых Листовидные сосочки развиты у детей (у взрослых они атрофированы), образуют две группы (4– 8 сосочков в каждой) по левому и правому краям тела языка. Грибовидные сосочки (около 100) расположены по дорсальной поверхности передних двух третях языка, рисунок их расположения индивидуален. Желобоватые сосочки (6– 12 штук) расположены в задней части языка, кпереди от пограничной борозды между телом и корнем языка.
Рецепторы вкуса — контактные, а пути проведения от них проходят через ствол мозга к таламусу и проецируются вдоль постцентральной извилины. Периферическая часть вкусового анализатора — вкусовые почки — расположены в слизистой оболочке полости рта, переднего отдела глотки, пищевода и гортани. Основная масса вкусовых почек (>90% их общего количества — до 10 тыс. ) находится в хемочувствительных сосочках языка — листовидных, грибовидных и желобоватых Листовидные сосочки развиты у детей (у взрослых они атрофированы), образуют две группы (4– 8 сосочков в каждой) по левому и правому краям тела языка. Грибовидные сосочки (около 100) расположены по дорсальной поверхности передних двух третях языка, рисунок их расположения индивидуален. Желобоватые сосочки (6– 12 штук) расположены в задней части языка, кпереди от пограничной борозды между телом и корнем языка.
 ВКУСОВЫЕ ПОЧКИ Вкусовые луковицы — тельца яйцевидной формы со средним размером 50– 70 мкм, расположены в пласте многослойного плоского эпителия, занимая всю толщу пласта, от базальной мембраны эпителия до его поверхности. Все клетки вкусовой луковицы постоянно обновляются за счёт окружающих клеток эпителия, но только при условии интактной чувствительной иннервации той области слизистой оболочки, где расположены вкусовые луковицы. Время жизни клеток вкусовой луковицы составляет 10 сут. Каждая вкусовая почка содержит поддерживающие (опорные) и вкусовые (рецепторные) клетки
ВКУСОВЫЕ ПОЧКИ Вкусовые луковицы — тельца яйцевидной формы со средним размером 50– 70 мкм, расположены в пласте многослойного плоского эпителия, занимая всю толщу пласта, от базальной мембраны эпителия до его поверхности. Все клетки вкусовой луковицы постоянно обновляются за счёт окружающих клеток эпителия, но только при условии интактной чувствительной иннервации той области слизистой оболочки, где расположены вкусовые луковицы. Время жизни клеток вкусовой луковицы составляет 10 сут. Каждая вкусовая почка содержит поддерживающие (опорные) и вкусовые (рецепторные) клетки
 РЕГЕНЕРАЦИЯ ВКУСОВЫХ КЛЕТОК Во вкусовом рецепторе происходит постоянное обновление клеток. Из периферической области вкусовой почки клетки перемещаются в центральную её часть со скоростью 0, 06 мкм/час. Средняя продолжительность жизни клеток вкусового рецепторного органа составляет 250± 50 часов. После повреждения нервов, иннервирующих вкусовые почки, последние дегенерируют, а при регенерации нервов происходит их восстановление.
РЕГЕНЕРАЦИЯ ВКУСОВЫХ КЛЕТОК Во вкусовом рецепторе происходит постоянное обновление клеток. Из периферической области вкусовой почки клетки перемещаются в центральную её часть со скоростью 0, 06 мкм/час. Средняя продолжительность жизни клеток вкусового рецепторного органа составляет 250± 50 часов. После повреждения нервов, иннервирующих вкусовые почки, последние дегенерируют, а при регенерации нервов происходит их восстановление.
 части большинства") Вкусовая почка состоит из удлинённых светлых клеток различных типов. На верхушечной (апикальной) части большинства клеток расположены микроворсинки — вкусовые волоски, занимающие вкусовой канал, открывающийся на поверхность эпителия вкусовой порой. В микроворсинки «вмонтированы» рецепторные вкусовые белки. В базальной части вкусовой луковицы рецепторные клетки образуют синапсы с терминальными разветвлениями периферических отростков первичных чувствительных нейронов. Каждая вкусовая почка иннервирована примерно 50 нервными волокнами, а каждый чувствительный нейрон получает сигналы примерно от 5 вкусовых луковиц.
Вкусовая почка состоит из удлинённых светлых клеток различных типов. На верхушечной (апикальной) части большинства клеток расположены микроворсинки — вкусовые волоски, занимающие вкусовой канал, открывающийся на поверхность эпителия вкусовой порой. В микроворсинки «вмонтированы» рецепторные вкусовые белки. В базальной части вкусовой луковицы рецепторные клетки образуют синапсы с терминальными разветвлениями периферических отростков первичных чувствительных нейронов. Каждая вкусовая почка иннервирована примерно 50 нервными волокнами, а каждый чувствительный нейрон получает сигналы примерно от 5 вкусовых луковиц.
 Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 1. первичного чувствительного нейрона; 2. нервной клетки ядра одиночного пути (одиночное ядро, nucleus tractus solitarius); 3. нейрона вентромедиального ядра таламуса
Восходящие пути вкусовой чувствительности состоят из трёх нейронов: 1. первичного чувствительного нейрона; 2. нервной клетки ядра одиночного пути (одиночное ядро, nucleus tractus solitarius); 3. нейрона вентромедиального ядра таламуса
 Первичные чувствительные нейроны Чувствительные нервные волокна от вкусовых почек, расположенных в передних двух третях языка, проходят в составе язычного нерва и далее — барабанной струны (chorda tympani системы лицевого нерва). Тела первичных чувствительных нейронов находятся в узле коленца (ganglion geniculi). Центральные отростки этих нейронов в составе промежуточного нерва (n. intermedius) и одиночного пути подходят к одиночному ядру (nucleus tractus solitarius). Афферентные волокна от вкусовых луковиц задней трети языка проходят в составе языкоглоточного нерва. Тела первичных чувствительных нейронов находятся в нижнем узле (ganglion inferius), расположенном у выхода из яремного отверстия. Центральные отростки нейронов в составе одиночного пути направляются к одиночному ядру. Чувствительные нервные волокна из более каудально расположенных областей входят в состав блуждающего нерва. Тела первичных чувствительных нейронов находятся в верхнем и нижнем узлах (ganglii superius et inferius), а их центральные отростки в составе одиночного пути направляются к одиночному ядру.
Первичные чувствительные нейроны Чувствительные нервные волокна от вкусовых почек, расположенных в передних двух третях языка, проходят в составе язычного нерва и далее — барабанной струны (chorda tympani системы лицевого нерва). Тела первичных чувствительных нейронов находятся в узле коленца (ganglion geniculi). Центральные отростки этих нейронов в составе промежуточного нерва (n. intermedius) и одиночного пути подходят к одиночному ядру (nucleus tractus solitarius). Афферентные волокна от вкусовых луковиц задней трети языка проходят в составе языкоглоточного нерва. Тела первичных чувствительных нейронов находятся в нижнем узле (ganglion inferius), расположенном у выхода из яремного отверстия. Центральные отростки нейронов в составе одиночного пути направляются к одиночному ядру. Чувствительные нервные волокна из более каудально расположенных областей входят в состав блуждающего нерва. Тела первичных чувствительных нейронов находятся в верхнем и нижнем узлах (ganglii superius et inferius), а их центральные отростки в составе одиночного пути направляются к одиночному ядру.
 Нейроны ядра одиночного пути Вторые нейроны восходящих путей вкусовой чувствительности расположены в ростральной части ядра восходящего пути. Входящие в ядро центральные отростки первичных чувствительных нейронов образуют синапсы с дендритами нервных клеток ядра, а их аксоны переходят на другую сторону, присоединяются к медиальной петле (lemniscus medialis) и следуют до вентромедиального ядра таламуса.
Нейроны ядра одиночного пути Вторые нейроны восходящих путей вкусовой чувствительности расположены в ростральной части ядра восходящего пути. Входящие в ядро центральные отростки первичных чувствительных нейронов образуют синапсы с дендритами нервных клеток ядра, а их аксоны переходят на другую сторону, присоединяются к медиальной петле (lemniscus medialis) и следуют до вентромедиального ядра таламуса.
 Нейроны вентромедиального ядра таламуса Аксоны нервных клеток ядра таламуса в составе лучистого венца (corona radiata) направляются к вентральной части постцентральной извилины. Представительство вкуса не имеет отдельной корковой проекционной области, оно представлено в участке постцентральной извилины, получающем проекции кожной чувствительности лица.
Нейроны вентромедиального ядра таламуса Аксоны нервных клеток ядра таламуса в составе лучистого венца (corona radiata) направляются к вентральной части постцентральной извилины. Представительство вкуса не имеет отдельной корковой проекционной области, оно представлено в участке постцентральной извилины, получающем проекции кожной чувствительности лица.
, а также") ОСНОВНЫЕ ВКУСЫ Человек различает четыре первичных вкуса (сладкий, кислый, горький и солёный), а также «умами» (от японского «изысканный» , вкус глутамата натрия). Существуют карты топографической привязанности преимущественного восприятия основных вкусов, но необходимо иметь в виду, что такие карты справедливы только для низких концентраций тестируемых вкусовых веществ. Так, при низких концентрациях деполяризуются и формируют рецепторный (генераторный) потенциал лишь отдельные вкусовые клетки. Некоторые вкусовые клетки и вкусовые луковицы отвечают преимущественно на горькие стимулы, в то время как другие — на сладкие, кислые или солёные. Часть вкусовых клеток и луковицы в целом отвечают на две или три вкусовые модальности, а иные даже на все. При увеличении же концентрации вкусовых веществ происходит возбуждение практически во всех вкусовых луковицах самой различной локализации
ОСНОВНЫЕ ВКУСЫ Человек различает четыре первичных вкуса (сладкий, кислый, горький и солёный), а также «умами» (от японского «изысканный» , вкус глутамата натрия). Существуют карты топографической привязанности преимущественного восприятия основных вкусов, но необходимо иметь в виду, что такие карты справедливы только для низких концентраций тестируемых вкусовых веществ. Так, при низких концентрациях деполяризуются и формируют рецепторный (генераторный) потенциал лишь отдельные вкусовые клетки. Некоторые вкусовые клетки и вкусовые луковицы отвечают преимущественно на горькие стимулы, в то время как другие — на сладкие, кислые или солёные. Часть вкусовых клеток и луковицы в целом отвечают на две или три вкусовые модальности, а иные даже на все. При увеличении же концентрации вкусовых веществ происходит возбуждение практически во всех вкусовых луковицах самой различной локализации
 ВКУСОВОЙ ПОРОГ Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и всех остальных первичных вкусов. Так, ощущение солёного возникает при воздействии вещества в концентрации 0, 01 М, а наличие хинина можно выявить при его концентрации в миллион раз меньшей. Пороги вкусовой чувствительности у разных лиц заметно различаются. Значительное влияние на чувствительность оказывает состояние организма (например, стресс, беременность, голодание). Часто наблюдают «вкусовую слепоту» (классический пример — производные тиомочевины, например фенилтиокарбамид, присутствие которого не чувствует каждый пятый). Способность человека различать интенсивность вкусовых ощущений сравнительно груба. Так, 20% раствор сахара ощущается как максимально сладкий, 10% раствор поваренной соли как максимально солёный и т. д.
ВКУСОВОЙ ПОРОГ Наиболее значительные различия наблюдают между веществами, создающими ощущение горького и всех остальных первичных вкусов. Так, ощущение солёного возникает при воздействии вещества в концентрации 0, 01 М, а наличие хинина можно выявить при его концентрации в миллион раз меньшей. Пороги вкусовой чувствительности у разных лиц заметно различаются. Значительное влияние на чувствительность оказывает состояние организма (например, стресс, беременность, голодание). Часто наблюдают «вкусовую слепоту» (классический пример — производные тиомочевины, например фенилтиокарбамид, присутствие которого не чувствует каждый пятый). Способность человека различать интенсивность вкусовых ощущений сравнительно груба. Так, 20% раствор сахара ощущается как максимально сладкий, 10% раствор поваренной соли как максимально солёный и т. д.
, гипогевзию (уменьшение вкусовой чувствительности)") НАРУШЕНИЯ ВКУСА К нарушениям вкуса относят агевзию (отсутствие чувства вкуса), гипогевзию (уменьшение вкусовой чувствительности) и дисгевзию (извращение вкуса). Гипогевзию наблюдают при многих заболеваниях. Некоторые вещества (например, содержащие сульфгидрильные группы каптоприл и пеницилламин) вызывают временную потерю вкусовой чувствительности
НАРУШЕНИЯ ВКУСА К нарушениям вкуса относят агевзию (отсутствие чувства вкуса), гипогевзию (уменьшение вкусовой чувствительности) и дисгевзию (извращение вкуса). Гипогевзию наблюдают при многих заболеваниях. Некоторые вещества (например, содержащие сульфгидрильные группы каптоприл и пеницилламин) вызывают временную потерю вкусовой чувствительности
 Механизмы возбуждения вкусовых клеток Вкусовое вещество – молекулярный вкусовой рецептор вкусовых волосков – вторые посредники – деполяризация клеточной мембраны и генерация рецепторного потенциала – поступление Са в цитоплазму из межклеточного пространства и/или внутриклеточных кальциевых депо – экзоцитоз синаптических пузырьков из базальной части клетки – постсинаптический потенциал – генерация ПД
Механизмы возбуждения вкусовых клеток Вкусовое вещество – молекулярный вкусовой рецептор вкусовых волосков – вторые посредники – деполяризация клеточной мембраны и генерация рецепторного потенциала – поступление Са в цитоплазму из межклеточного пространства и/или внутриклеточных кальциевых депо – экзоцитоз синаптических пузырьков из базальной части клетки – постсинаптический потенциал – генерация ПД
 Сенсорная система кожи
Сенсорная система кожи
 Чувствительные нервные окончания А — свободные нервные окончания в соединительной ткани; Б — Комплекс клетки Меркеля с нервной терминалью; В — Тельце Майсснера; Г — инкапсулированное тельце Пачини. 1 — нервная терминаль, 2 — вспомогательные клетки, 3 — миелиновые нервные волокна, 4 — капсула. Масштаб между изображениями А–Г не выдержан.
Чувствительные нервные окончания А — свободные нервные окончания в соединительной ткани; Б — Комплекс клетки Меркеля с нервной терминалью; В — Тельце Майсснера; Г — инкапсулированное тельце Пачини. 1 — нервная терминаль, 2 — вспомогательные клетки, 3 — миелиновые нервные волокна, 4 — капсула. Масштаб между изображениями А–Г не выдержан.
 Восходящие пути чувствительности А. Путь от чувствительных нейронов спинномозговых узлов (первый, или первичный чувствительный нейрон) через вторые нейроны (вставочные нейроны спинного мозга или нервные клетки клиновидного и тонкого ядра продолговатого мозга) к третьим нейронам пути — таламическим. Аксоны этих нейронов направляются к коре головного мозга. Б. Расположение нейронов, передающих разные модальности, в пластинах (римские цифры) спинного мозга
Восходящие пути чувствительности А. Путь от чувствительных нейронов спинномозговых узлов (первый, или первичный чувствительный нейрон) через вторые нейроны (вставочные нейроны спинного мозга или нервные клетки клиновидного и тонкого ядра продолговатого мозга) к третьим нейронам пути — таламическим. Аксоны этих нейронов направляются к коре головного мозга. Б. Расположение нейронов, передающих разные модальности, в пластинах (римские цифры) спинного мозга
 Проводящие пути проприоцептивной и тактильной чувствительности Тонкий и клиновидный пучки — проходят в составе заднего канатика той же стороны спинного мозга и заканчиваются в тонком и клиновидном ядрах продолговатого мозга. Аксоны нейронов этих ядер по медиальной петле (отсюда и название — лемнисковая система) переходят на противоположную сторону и направляются к таламусу
Проводящие пути проприоцептивной и тактильной чувствительности Тонкий и клиновидный пучки — проходят в составе заднего канатика той же стороны спинного мозга и заканчиваются в тонком и клиновидном ядрах продолговатого мозга. Аксоны нейронов этих ядер по медиальной петле (отсюда и название — лемнисковая система) переходят на противоположную сторону и направляются к таламусу
 Тактильные и прессорные ощущения от механорецепторов кожи Спиноталамический путь вентральный проекционный афферентный путь, проходящий в переднем канатике противоположной стороны. Центральные отростки этих нейронов вступают через задние корешки в задние канатики, где поднимаются на 2– 15 сегментов и образуют синапсы с вставочными нейронами задних рогов. Аксоны этих нейронов переходят на противоположную сторону и проходят далее в передней периферической зоне переднебоковых канатиков. Отсюда волокна пути восходят к заднелатеральному вентральному ядру таламуса вместе с латеральным спиноталамическим путём
Тактильные и прессорные ощущения от механорецепторов кожи Спиноталамический путь вентральный проекционный афферентный путь, проходящий в переднем канатике противоположной стороны. Центральные отростки этих нейронов вступают через задние корешки в задние канатики, где поднимаются на 2– 15 сегментов и образуют синапсы с вставочными нейронами задних рогов. Аксоны этих нейронов переходят на противоположную сторону и проходят далее в передней периферической зоне переднебоковых канатиков. Отсюда волокна пути восходят к заднелатеральному вентральному ядру таламуса вместе с латеральным спиноталамическим путём
 Путь проведения болевой и температурной чувствительности Спиноталамический путь латеральный — проекционный афферентный путь, проходящий в боковом канатике. Периферическими рецепторами являются свободные нервные окончания кожи. Центральные отростки псевдоуниполярных нейронов спинномозговых узлов входят в противоположную часть спинного мозга через латеральные отделы задних корешков и, поднявшись в спинном мозге на 1– 2 сегмента, образуют синапсы с нейронами роландова студенистого вещества. Аксоны этих нейронов фактически образуют латеральный спиноталамический путь. Они идут на противоположную сторону и поднимаются в латеральных отделах боковых канатиков. Спиноталамические пути проходят через ствол мозга и заканчиваются в вентро латеральных ядрах таламуса.
Путь проведения болевой и температурной чувствительности Спиноталамический путь латеральный — проекционный афферентный путь, проходящий в боковом канатике. Периферическими рецепторами являются свободные нервные окончания кожи. Центральные отростки псевдоуниполярных нейронов спинномозговых узлов входят в противоположную часть спинного мозга через латеральные отделы задних корешков и, поднявшись в спинном мозге на 1– 2 сегмента, образуют синапсы с нейронами роландова студенистого вещества. Аксоны этих нейронов фактически образуют латеральный спиноталамический путь. Они идут на противоположную сторону и поднимаются в латеральных отделах боковых канатиков. Спиноталамические пути проходят через ствол мозга и заканчиваются в вентро латеральных ядрах таламуса.
 Соматосенсорная область I") Соматосенсорная кора Передняя половина теменной доли (рецепция и интерпритация соматосенсорных сигналов) Соматосенсорная область I располагается непосредственно позади центральной борозды (поля 1, 2, 3 по Бродману) В ней плотно расположены точки восприятия различных частей тела – корковое представительство Задняя половина теменной доли – высший уровень интерпитации Соматосенсорная область II получает сигналы от туловища и конечностей, а также других ассоциативных сенсорных областей (слуховой, зрительной и т. д. )
Соматосенсорная кора Передняя половина теменной доли (рецепция и интерпритация соматосенсорных сигналов) Соматосенсорная область I располагается непосредственно позади центральной борозды (поля 1, 2, 3 по Бродману) В ней плотно расположены точки восприятия различных частей тела – корковое представительство Задняя половина теменной доли – высший уровень интерпитации Соматосенсорная область II получает сигналы от туловища и конечностей, а также других ассоциативных сенсорных областей (слуховой, зрительной и т. д. )
 Соматотопическое представительство областей тела сенсомоторной области I
Соматотопическое представительство областей тела сенсомоторной области I
 Функциональная единица соматосенсорной коры – колонки нейронов Каждая колонка имеет диаметр от 0, 3 до 0, 5 мм Содержит около 10 тыс. тел нервных клеток Обслуживает отдельную специфическую модальность Осуществляют начальный синтез значения сенсорных сигналов
Функциональная единица соматосенсорной коры – колонки нейронов Каждая колонка имеет диаметр от 0, 3 до 0, 5 мм Содержит около 10 тыс. тел нервных клеток Обслуживает отдельную специфическую модальность Осуществляют начальный синтез значения сенсорных сигналов