!Лекция 6. Сцепленное наследование.ppt
- Количество слайдов: 52



СЦЕПЛЕННОЕ НАСЛЕДОВАНИЕ КРОССИНГОВЕР Общая генетика

Сцепленное наследование • Согласно третьему закону Менделя, максимально возможное число генов, которые могут подчиняться закону независимого наследования признаков, у каждого вида не может превышать его гаплоидного числа хромосом. Однако на самом деле количестве генов у каждого вида на несколько порядков выше. Как же наследуются все остальные гены, т. е. гены, локализованные в одной паре гомологичных хромосом? Каким законам они подчиняются?

3 • В 1902 г. Бовери и Сэттон сформулировали основные положения хромосомной теории наследственности – учения о локализации наследственных факторов в хромосомах: • гены расположены в хромосомах • в каждой гомологичной хромосоме находится по одному аллельному гену • гомологичные хромосомы и вместе с ними аллельные гены объединяются в зиготе во время оплодотворения

1. ПОЛНОЕ АУТОСОМНОЕ СЦЕПЛЕНИЕ • Число признаков и свойств организма велико, а число пар хромосом у каждого вида относительно мало (6… 50). Следовательно, в одной хромосоме локализованы не один, а много генов. • Гены, локализованные в одной хромосоме, передаются потомкам совместно. • При наследовании они не подчиняются 2 и 3 законам Менделя, основой которых является свободная перекомбинация генов, находящихся в разных парах гомологичных хромосом.

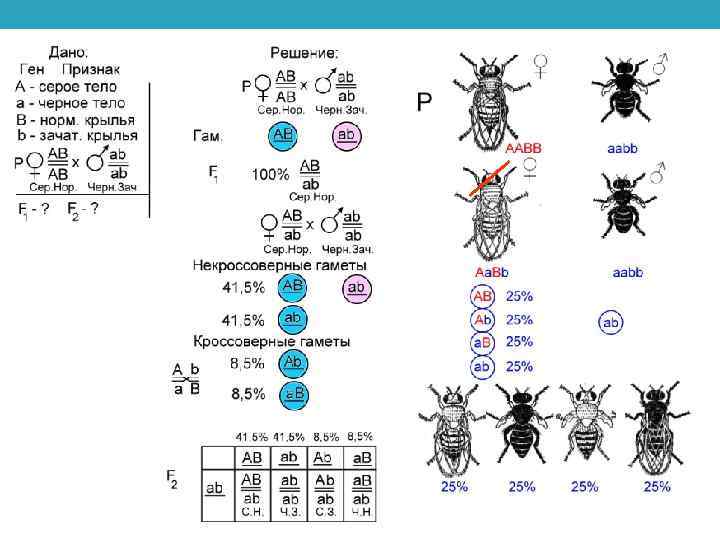
Опыты У. Бэтсона и Р. Пеннета, демонстрирующие сцепление генов Впервые совместное наследование признаков обнаружили У. Бетсон и Р. Пеннет (1906) в опытах по скрещиванию душистого горошка. В дальнейшем оно было обнаружено и у других видов. Вместо менделевского расщепления в F 2: 9 А-В-: 3 А-bb: 3 аа. В-: 1 ааbb, или в FА: 1 А-В-: 1 А-bb: 1 аа. В-: 1 ааbb в F 2 и FА наблюдалось преобладание особей с родительским сочетанием признаков, а новые комбинации признаков отсутствовали или наблюдались с небольшой частотой.

Гипотеза Моргана о сцеплении генов и возможности кроссинговера Изучение причины этого явления впервые провел американский генетик Т. Х. Морган (1910) в опытах на дрозофиле. Он скрестил мух линии вэстиджел в+в+vgvg (серое тело и зачаточные крылья) с мухами линии блэк ввvg+vg+ (черное тело и нормальные крылья)

Скрещивая мушку дрозофилу с серым телом и нормальными крыльями с мушкой, имеющей темную окраску тела и зачаточные крылья, в первом поколении Морган получал гибридов, имеющих серое тело и нормальные крылья. При проведении анализирующего скрещивания самки F 1 с самцом, имевшим рецессивные признаки, теоретически ожидалось получить потомство с комбинациями этих признаков в соотношении 1: 1: 1: 1.

Однако в потомстве было 41, 5% серых длиннокрылых и 41, 5% черных с зачаточными крыльями и лишь незначительная часть мушек имела перекомбинированные признаки (8, 5% черные длиннокрылые и 8, 5% серые с зачаточными крыльями). Морган пришел к выводу, что гены, обусловливающие развитие серой окраски тела и длинных крыльев, локализованы в одной хромосоме, а гены, обусловливающие развитие черной окраски тела и зачаточных крыльев, — в другой.

Явление совместного наследования признаков Морган назвал сцеплением. Материальной основой сцепления генов является хромосома. Гены, локализованные в одной хромосоме, наследуются совместно и образуют одну группу сцепления. Поскольку гомологичные хромосомы имеют одинаковый набор генов, количество групп сцепления равно гаплоидному набору хромосом Явление совместного наследования генов, локализованных в одной хромосоме, называют сцепленным наследованием. Сцепленное наследование генов, локализованных в одной хромосоме, называют законом Моргана.

Появление особей с перекомбинированными признаками Морган объяснил кроссинговером во время мейоза. В результате кроссинговера в некоторых клетках происходит обмен участками хромосом между генами А и В, появляются гаметы Ав и а. В, и, как следствие, в потомстве образуются четыре группы фенотипов, как при свободном комбинировании генов. Но поскольку кроссинговер происходит не во всех гаметах, числовое соотношение фенотипов не соответствует соотношению 1: 1: 1: 1.

В зависимости от особенностей образования гамет, различают: некроссоверные гаметы — гаметы с хромосомами, образованными без кроссинговера: кроссоверные гаметы — гаметы с хромосомами, претерпевшими кроссинговер:

Гены, расположенные в одной хромосоме, образуют группу сцепления Число групп сцепления равно гаплоидному числу хромосом вида: у человека – 23 (2 n=46) у ржи – 7 (2 n=14) у мягкой пшеницы – 21 (2 n=42) у дрозофилы – 4 (2 n=8)

Гены, локализованные в одной хромосоме, наследуются совместно и образуют одну группу сцепления. В каждой паре гомологичных хромосом находятся одинаковые группы генов. У человека 23 группы сцепления, у дрозофилы – четыре. Было также показано, что у каждого гена в хромосоме есть строго определенное место — локус.

Вероятность возникновения перекреста между генами зависит от их расположения в хромосоме: чем дальше друг от друга расположены гены, тем выше вероятность перекреста между ними.

2. НЕПОЛНОЕ СЦЕПЛЕНИЕ. КРОССИНГОВЕР • Полное аутосомное сцепление у дрозофилы наблюдается только у самцов, у самок оно никогда не бывает полным. • При скрещивании с самцами линиианализатора самок F 1 в FА было получено четыре фенотипических класса, но их соотношение отличалось от менделевского (1: 1: 1: 1): преобладали особи с родительским сочетанием признаков

За единицу расстояния между генами, находящимися в одной хромосоме, принят 1% кроссинговера. Например, в рассмотренном выше анализирующем скрещивании получено 17% особей с перекомбинированными признаками. Следовательно, расстояние между генами серой окраски тела и длинных крыльев (а также черной окраски тела и зачаточных крыльев) равно 17%.

В честь Т. Моргана единица расстояния между генами названа морганидой, расстояние между этими генами равно 17 морганидам. А сила сцепления высчитывается по формуле: сила сцепления = 100% - % кроссоверных гамет. Сила сцепления между генами окраски тела и формы крыльев равна 100% - 17% = 83%.

в+++ в+ в в vg в+ в++ в в в+ + vg в+ в+ vg+ в+ в+ + vg в в+ в+ vg+ в+ в+ в vg Для объяснения полученных результатов скрещивания Т. Х. Морган выдвинул гипотезу:

Схема опыта Т. Х. Моргана ♂ в+vg х ♀ вvg+ серое тело черное тело зачаточные крылья нормальные крылья гаметы вvg+ в+vg F 1 ♀ в+vg вvg+ х серое тело нормальные крылья некроссоверные гаметы в+vg вvg+ кроссоверные гаметы вvg FA некроссоверы в+vg+ в+vg вvg + вvg серое тело зачаточные крылья черное тело нормальные крылья 41, 5% 83% ♂вvg черное тело зачаточные крылья вvg кроссоверы вvg в+vg + вvg черное тело серое тело зачаточные крылья нормальные крылья 8, 5% 17%

Факторы, влияющие на частоту кроссинговера • Температура – высокая и низкая температура повышает частоту • Рентгеновские лучи – повышают • Возраст – снижает • Мутагены – повышают или снижают • Пол – у самцов дрозофилы кроссинговер не происходит, у самцов млекопитающих интенсивность кроссинговера незначительна • Расстояние между генами • Пищевые эффекты – Са и Mg могут повышать или снижать • Центромерные эффекты – гены в области центромеры менее способны к кроссинговеру

Генетическое доказательство кроссинговера Из анализа расщепления в FА при кроссинговере некроссоверы, несущие родительские сочетания признаков, образуются в равном количественном соотношении (41, 5 и 41, 5%), кроссоверы (с перекомбинированными признаками) тоже в равном количестве (8, 5 и 8, 5%). Но численность некроссоверов (нерекомбинантов) всегда превышает число кроссоверов (рекомбинантов) и их соотношение отличается от менделевского 1: 1: 1: 1 при независимом наследовании, что и является генетическим доказательством кроссинговера.

Сравнительный анализ сцепленного и несцепленного наследования Гены ~25 000 генов 46 хромосом Несцепленные (в разных хромосомах) А Сцепленные (в одной хромосоме) Полное (абсолютное) сцепление a B b Неполное сцепление А А b B b a B a
 Р:")
I. Гены типа А и В находятся в разных гомологичных хромосомах (независимое наследование) Р: Aa. Bb А G: a A B 25% x B A aabb b b 25% a a B a 25% b 25% 25% a b b 100% F 1: Aa. Bb, Aabb, aa. Bb, aabb 25% Анализирующее скрещивание 25%


II. Полностью сцепленное наследование Р: Aa. Bb А G: b aabb Анализирующее скрещивание a B x a А b b b a B a 50% F 1: Aa. Bb, aabb 50% a b 100%


III. Гены А и В сцеплены в одной паре гомологичных хромосом, но между ними наблюдается кроссинговер. (его вероятность – 20%) Анализирующее Р: Aa. Bb x aabb скрещивание А B G: a b A a A b b a B B a a A a b a 100% b A 40% B некр A B a a 40% b b B некр 10% кросс10%

F 1: Aa. Bb, Aabb, aa. Bb, aabb 40% 10% 40% некр. кросс. некр. Некроссоверные гаметы >>> Кроссоверные гаметы


Анализ наследования трех сцепленных генов

Сравнительный анализ сцепленного и несцепленного наследования • Критерии для разграничения сцепленного и не сцепленного наследования являются результаты анализирующего скрещивания. Если процент рекомбинантных потомков равен 50, то признаки не сцеплены (наследуются независимо, находятся в разных парах гомологичных хромосом). Если процент рекомбинантных (кроссоверных) потомков < 50, то признаки сцеплены (находятся в одной паре гомологичных хромосом). • Чем ближе расположены признаки, тем меньший процент кроссинговера. Таким образом, определяя силу сцепления между генами, можно определить расстояние между ними в хромосоме.

Значение кроссинговера • Кроссинговер — широко распространенное явление • Происходит практически у всех организмов, которые размножаются половым путем • Этот процесс является молекулярной основой комбинативной изменчивости • В результате рекомбинации генов могут появляться новые полезные признаки и их сочетания. Поэтому кроссинговер имеет большое значение для выживания и размножения • Этот процесс также увеличивает генетическое разнообразие потомков, что очень важно для приспособления и эволюции • Определение частоты кроссинговера лежит в основе картирования генов хромосом, то есть определения места расположения разных генов в хромосоме
. Современная хромосомная теория наследственности создана выдающимся американским")
Хромосомная теория наследственности: Томас Морган (1866— 1945). Современная хромосомная теория наследственности создана выдающимся американским генетиком Томасом Морганом (1866 — 1945). 1. Гены располагаются в хромосомах; различные хромосомы содержат неодинаковое число генов, причем набор генов каждой из негомологичных хромосом уникален; 2. Каждый ген имеет определенное место (локус) в хромосоме; в идентичных локусах гомологичных хромосом находятся аллельные гены; 3. Гены расположены в хромосомах в определенной линейной последовательности; 4. Гены, локализованные в одной хромосоме, наследуются совместно, образуя группу сцепления; число групп сцепления равно гаплоидному набору хромосом и постоянно для каждого вида организмов;

Хромосомная теория наследственности: 5. Сцепление генов может нарушаться в процессе кроссинговера; это приводит к образованию рекомбинантных хромосом; 6. Частота кроссинговера является функцией расстояния между генами: чем больше расстояние, тем больше величина кроссинговера (прямая зависимость); 7. Каждый вид имеет характерный только для него набор хромосом — кариотип. Явление кроссинговера помогло ученым установить расположение каждого гена в хромосоме, создать генетические карты хромосом. Чем дальше друг от друга расположены на хромосоме два гена, тем чаще они будут расходиться в разные хромосомы в процессе кроссинговера.

ВЕЛИЧИНА КРОССИНГОВЕРА. ЛИНЕЙНОЕ РАСПОЛОЖЕНИЕ ГЕНОВ В ХРОМОСОМЕ • Величина кроссинговера равна процентному содержанию кроссоверных особей в потомстве от анализирующего скрещивания, т. е. выражается в процентах. Частота кроссинговера не зависит от аллельного состояния генов, участвующих в скрещивании, если в качестве родителей брали мух вvg и в+vg+ вvg в+vg+ то в FА кроссоверы получались с той же частотой: 17%. • Следовательно, сцепление генов реально существует и положение их локусов в хромосоме фиксировано.

• Т. Х. Морган скрестил гетерозиготных по трем сцепленным генам самок y+w+bi+ ywbi с самцами, у которых эти гены были в рецессивном состоянии ywbi. В потомстве мухи дрозофилы было получено такое соотношение в величине кроссинговера: желтое (у) __ белые (w) _____ вильчатые (bi) тело 1, 2% глаза 3, 5% крылья 4, 7% Такая зависимость характеризует положение трех точек на прямой линии.

• Для нахождения локуса гена в хромосоме скрещивания проводят так, чтобы локус определяемого гена был третьей точкой. Это необходимо для установления порядка расположения генов в хромосоме. • Величина кроссинговера между генами y и w была равна 1, 2%, между генами w и bi – 3, 5%. На основании этих данных нельзя определить взаиморасположение генов у-w и w-bi (справа или слева друг от друга). • Только определив величину кроссинговера между генами у и bi (4, 7%), можно сделать заключение о том, что ген w находится между этими генами.

ПРИНЦИПЫ СОСТАВЛЕНИЯ ХРОМОСОМНЫХ КАРТ • Карты хромосом – это графическое представление относительного расположения и расстояния между генами в группе сцепления (хромосоме) • Различают генетические карты: 1. Факториальные (генетические) 2. Физические
. Поэтому")
Факториальные карты хромосом • Факториальные строят по проценту кроссинговера (или сцепления между генами). Поэтому они являются относительными. • Сила сцепления пропорциональна проценту кроссинговера. • В хромосомах существуют горячие точки, где кроссинговер происходит чаще. Факториальные генетические карты являются относительными (а последовательность генов они отображают правильно).

Физические карты хромосом • Физическое картирование использует молекулярно-биологические методы для непосредственного исследования молекул ДНК и создания карт, обозначающих позиции определенных последовательностей, в том числе генов.

Типы физических карт 1. Цитологические строятся под микроскопом с учётом морфологических особенностей хромосом (полигенные хромосомы находятся в стадии конъюгации, объединены в 4 -ре бивалента; находятся в слюнных железах двукрылых; хромосомы толстые, видны в интерфазе; нити спирализованы неравномерно: гетерохроматин спирализован – он неактивен; эухроматин деспирализован – он активен). 2. Рестрикционные 3. Секвенирование хромосомы

Определение группы сцепления • Для локализации новой мутации на генетических картах проводят 2 этапа: • 1. Определение группы сцепления. • 2. Определение местоположения мутации в пределах данной хромосомы.

Определение группы сцепления Пусть Х – новая мутация у дрозофилы. У дрозофилы 4 -ре пары хромосом. Чтобы определить где находится мутация Х, необходимо иметь маркёрные гены в каждой хромосоме. Проводим анализирующее скрещивание и определяем результаты. сцепление Х с: a x 50 % (не сцеплен) b х 49 % (не сцеплен) c х 25 % (сцеплен) d х 50 % (не сцеплен) => мутация Х сцеплена с геном d, т. е. находится в 3 -ей хромосоме.

Локализация гена в хромосоме
")
Определение силы сцепления между тремя генами (1)
")
Определение силы сцепления между тремя генами (2)
 Теоретически ожидаемая вероятность двойного кроссинговера = 0,")
Определение силы сцепления между тремя генами (3) Теоретически ожидаемая вероятность двойного кроссинговера = 0, 19 x 0, 29 = 0, 0551 Практически полученная частота двойных кроссинговеров = 40/100 = 0, 04
 • Теоретически ожидаемая вероятность кроссинговера = 0,")
Определение силы сцепления между тремя генами (4) • Теоретически ожидаемая вероятность кроссинговера = 0, 19 x 0, 29 = 0, 0551 • Практически полученная частота кроссинговеров = 40/100 = 0, 04. двойного двойных Чаще всего наблюдается несоответствие теоретически ожидаемых и практически получаемых вероятностей. Это несоответствие называется коэффициентом конциденции. Происходит интерференция (повышение или уменьшение количества кроссоверов). С – коэффициент С = % = 72 %

Участок генетической карты II хромосомы дрозофилы

Генетическая карта II хромосомы томата • D – нормальная высота растения; • d – карликовые; • O – круглый плод; • o – овальный; • Ne – нормальные листья; • ne – пораженные листья болезнью; • Bk – круглый плод; • bk – плод с заостренным концом

Генетическая карта ХI хромосомы томата • F – гладкий плод • f – ребристый плод • Lf – Необлиственное соцветие • lf – облиственное соцветие
!Лекция 6. Сцепленное наследование.ppt