

C3- i С4-шлях фотосинтиезу Лобанова Л.В..ppt
- Количество слайдов: 18
 С 3 - i C 4 - шлях фотосинтезу Підготувала студентка 5 курсу Лобанова Л. В.
С 3 - i C 4 - шлях фотосинтезу Підготувала студентка 5 курсу Лобанова Л. В.
 С 3 -шлях фотосинтезу або цикл Кальвіна Цей шлях асиміляції СО 2 виявлений у всіх фотосинтезуючих рослин. Він був названий циклом Кальвіна на честь американського біохіміка М. Кальвіна, який із співробітниками відкрив і вивчив його в 1946 -1956 роках за допомогою методу мічених атомів та хроматографії. Були використані одноклітинні зелені водорості (хлорела тощо) і мічений 14 С 02. Фотосинтезуючі водорості вносили в середовище, що містить 14 С 02, на різні проміжки часу, потім клітини швидко фіксували, екстрагували з них спиротрозчиннi речовини і визначали вміст 14 С в різних з'єднаннях після їх поділу за допомогою хроматографії.
С 3 -шлях фотосинтезу або цикл Кальвіна Цей шлях асиміляції СО 2 виявлений у всіх фотосинтезуючих рослин. Він був названий циклом Кальвіна на честь американського біохіміка М. Кальвіна, який із співробітниками відкрив і вивчив його в 1946 -1956 роках за допомогою методу мічених атомів та хроматографії. Були використані одноклітинні зелені водорості (хлорела тощо) і мічений 14 С 02. Фотосинтезуючі водорості вносили в середовище, що містить 14 С 02, на різні проміжки часу, потім клітини швидко фіксували, екстрагували з них спиротрозчиннi речовини і визначали вміст 14 С в різних з'єднаннях після їх поділу за допомогою хроматографії.
 n Виявилося, що після експозиції протягом 1 хв 14 С включався в С 3 С 7 -цукри, в органічні кислоти (яблучну, щавлевооцтову), в амінокислоти(аланін, аспарагінову кислоту). Якщо ж час експозиції було скорочено до 0, 1 -2 с, то велика частина мітки виявлялася в фосфогліцеріновiй кислотi, а саме в її карбоксильнiй групi: Отже, 3 -фосфогліцерінова кислота (ФГК) є первинним продуктом фотосинтезу.
n Виявилося, що після експозиції протягом 1 хв 14 С включався в С 3 С 7 -цукри, в органічні кислоти (яблучну, щавлевооцтову), в амінокислоти(аланін, аспарагінову кислоту). Якщо ж час експозиції було скорочено до 0, 1 -2 с, то велика частина мітки виявлялася в фосфогліцеріновiй кислотi, а саме в її карбоксильнiй групi: Отже, 3 -фосфогліцерінова кислота (ФГК) є первинним продуктом фотосинтезу.
 Цикл Кальвіна складається з трьох етапів: карбоксилювання, n відновлення, n регенерації первинного акцептора СО 2 і синтезу кінцевого продукту фотосинтезу. n
Цикл Кальвіна складається з трьох етапів: карбоксилювання, n відновлення, n регенерації первинного акцептора СО 2 і синтезу кінцевого продукту фотосинтезу. n
 1. Карбоксилювання. Молекули рибульозо-5 -фосфату фосфорилюються за участю АТФ і фосфорибульозокінази, в результаті чого утворюються молекули рибульозо-1, 5 дифосфату, до яких у свою чергу приєднується С 02 за допомогою рибульозодифосфаткарбоксилази. Отриманий продукт розщеплюється на дві тріози: 2 молекули 3 -фосфогліцериновi кислоти (3 -ФГК).
1. Карбоксилювання. Молекули рибульозо-5 -фосфату фосфорилюються за участю АТФ і фосфорибульозокінази, в результаті чого утворюються молекули рибульозо-1, 5 дифосфату, до яких у свою чергу приєднується С 02 за допомогою рибульозодифосфаткарбоксилази. Отриманий продукт розщеплюється на дві тріози: 2 молекули 3 -фосфогліцериновi кислоти (3 -ФГК).
 СН 2 ОР | | С=О С - ОН | || СНОН С – ОН | | СНОН | | СН 2 ОР Рибульозо Енольна форма 1, 5 - дифосфат РДФ СН 2 ОР | СНОН | СООН | СНОН | СН 2 ОР 3 - фосфогліцеронова к-та (ФГК)
СН 2 ОР | | С=О С - ОН | || СНОН С – ОН | | СНОН | | СН 2 ОР Рибульозо Енольна форма 1, 5 - дифосфат РДФ СН 2 ОР | СНОН | СООН | СНОН | СН 2 ОР 3 - фосфогліцеронова к-та (ФГК)
 2. Відновлення. Фосфогліцераткіназа за участю АТФ фосфорилює 3 -ФГК, внаслідок чого утворюється 1, 3 -дифосфогліцеринова кислота, яка відновлюється за допомогою НАДФ Н і дегідрогенази фосфогліцерінового альдегіду до 3 фосфогліцерінового альдегіду (3 -ФГА).
2. Відновлення. Фосфогліцераткіназа за участю АТФ фосфорилює 3 -ФГК, внаслідок чого утворюється 1, 3 -дифосфогліцеринова кислота, яка відновлюється за допомогою НАДФ Н і дегідрогенази фосфогліцерінового альдегіду до 3 фосфогліцерінового альдегіду (3 -ФГА).
") Цикл Кальвіна (С 3 -шлях фотосинтезу)
Цикл Кальвіна (С 3 -шлях фотосинтезу)
 3. Фаза регенерації первинного акцептора діоксиду вуглецю і синтезу кінцевого продукту фотосинтезу. У результаті описаних вище реакцій при фіксації трьох молекул С 02 і утворенні шести молекул відновлених 3 -фосфотріоз п'ять з них використовуються потім для регенерації рибулозо-5 -фосфату, а одна для синтезу глюкози. 3 -ФГА під дією тріозофосфатізомерази ізомеризується в фосфодіоксіацетон. За участю альдолази 3 -ФГА і фосфодіоксіацетон конденсуються з утворенням фруктозо-1, 6 -дифосфату, у якого відщеплюється один фосфат за допомогою фруктозо-1, 6 -діфосфатази. У подальших реакціях, пов'язаних з регенерацією первинного акцептора С 02, послідовно приймають участь транскетолаза і альдолаза. Транскетолаза каталізує перенесення два вуглецю глiколевого альдегіду від кетози на адьдозу:
3. Фаза регенерації первинного акцептора діоксиду вуглецю і синтезу кінцевого продукту фотосинтезу. У результаті описаних вище реакцій при фіксації трьох молекул С 02 і утворенні шести молекул відновлених 3 -фосфотріоз п'ять з них використовуються потім для регенерації рибулозо-5 -фосфату, а одна для синтезу глюкози. 3 -ФГА під дією тріозофосфатізомерази ізомеризується в фосфодіоксіацетон. За участю альдолази 3 -ФГА і фосфодіоксіацетон конденсуються з утворенням фруктозо-1, 6 -дифосфату, у якого відщеплюється один фосфат за допомогою фруктозо-1, 6 -діфосфатази. У подальших реакціях, пов'язаних з регенерацією первинного акцептора С 02, послідовно приймають участь транскетолаза і альдолаза. Транскетолаза каталізує перенесення два вуглецю глiколевого альдегіду від кетози на адьдозу:
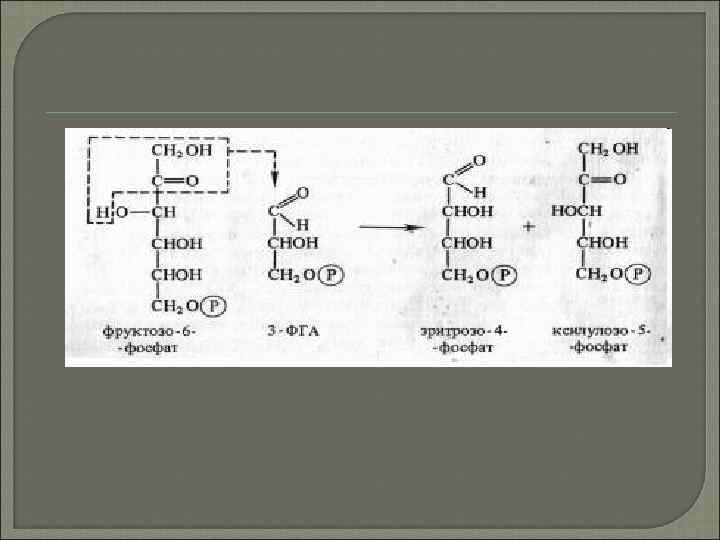
 Альдолаза потім здійснює перенесення трьохвуглецевого залишку фосфодіоксіацетона на альдози, в даному випадку ерітрозо-4 -фосфат, в результаті чого синтезується седо-гептульозо-1 , 7 дифосфат. Останній дефосфорилюється і під дією транскетолази з нього і 3 -ФГА утворюються ксилюлозо -5 -фосфат і рибоза-5 -фосфат. Дві молекули ксилюлозо 5 -фосфату за участю рибульозофосфатепімерази і одна молекула рибоза-5 -фосфату з участю рибозофосфатізомерази перетворюються на три молекули рибульозо-5 -фосфату, з якого починається новий цикл фіксації С 02. Iз невикористаної молекули 3 -ФГА під дією альдолази синтезується (при повторенні циклу) молекула фруктозо-1, 6 -дифосфату, з якої можуть утворюватися глюкоза, сахароза або крохмаль.
Альдолаза потім здійснює перенесення трьохвуглецевого залишку фосфодіоксіацетона на альдози, в даному випадку ерітрозо-4 -фосфат, в результаті чого синтезується седо-гептульозо-1 , 7 дифосфат. Останній дефосфорилюється і під дією транскетолази з нього і 3 -ФГА утворюються ксилюлозо -5 -фосфат і рибоза-5 -фосфат. Дві молекули ксилюлозо 5 -фосфату за участю рибульозофосфатепімерази і одна молекула рибоза-5 -фосфату з участю рибозофосфатізомерази перетворюються на три молекули рибульозо-5 -фосфату, з якого починається новий цикл фіксації С 02. Iз невикористаної молекули 3 -ФГА під дією альдолази синтезується (при повторенні циклу) молекула фруктозо-1, 6 -дифосфату, з якої можуть утворюватися глюкоза, сахароза або крохмаль.
 С 4 -шлях фотосинтезу або цикл Хетча-Слека Австралійськими вченими М. Хетчем і К. Слеком був описаний С 4 -шлях фотосинтезу, характерний для тропічних і субтропічних рослин (цукрова тростина, кукурудза та ін. ) Листки цих рослин мають хлоропласти двох типів: звичайні в клітинах мезофіла і великі хлоропласти, що не мають гран і фотосистеми II, в клітинах обкладки, якi оточуючують провідні пучки. У цитоплазмі клітин мезофіла фосфоенолпiруваткарбоксилаза приєднує СО 2 до фосфоенолпіровиноградної кислоти, утворюючи щавлевооцтову кислоту. Вона транспортується в хлоропласти, де відновлюється до яблучної кислоти за участю НАДФ Н. В присутності іонів амонію щавлевооцтова кислота перетворюється в аспарагінову кислоту.
С 4 -шлях фотосинтезу або цикл Хетча-Слека Австралійськими вченими М. Хетчем і К. Слеком був описаний С 4 -шлях фотосинтезу, характерний для тропічних і субтропічних рослин (цукрова тростина, кукурудза та ін. ) Листки цих рослин мають хлоропласти двох типів: звичайні в клітинах мезофіла і великі хлоропласти, що не мають гран і фотосистеми II, в клітинах обкладки, якi оточуючують провідні пучки. У цитоплазмі клітин мезофіла фосфоенолпiруваткарбоксилаза приєднує СО 2 до фосфоенолпіровиноградної кислоти, утворюючи щавлевооцтову кислоту. Вона транспортується в хлоропласти, де відновлюється до яблучної кислоти за участю НАДФ Н. В присутності іонів амонію щавлевооцтова кислота перетворюється в аспарагінову кислоту.
 аспарагінова кислоти переходять в хлоропласти клітин обкладки, декарбоксилються до піровиноградної кислоти") Яблучна і (або) аспарагінова кислоти переходять в хлоропласти клітин обкладки, декарбоксилються до піровиноградної кислоти і СО 2 включається в цикл Кальвіна, а піровиноградна кислота переноситься в клітини мезофілла, де перетворюється в фосфоенолпіровіноградну кислоту. Такий механізм дозволяє рослинам фотосинтезувати iз закритими продихами, через високу температуру.
Яблучна і (або) аспарагінова кислоти переходять в хлоропласти клітин обкладки, декарбоксилються до піровиноградної кислоти і СО 2 включається в цикл Кальвіна, а піровиноградна кислота переноситься в клітини мезофілла, де перетворюється в фосфоенолпіровіноградну кислоту. Такий механізм дозволяє рослинам фотосинтезувати iз закритими продихами, через високу температуру.
.") Цикл Хетча і Слека (С 4 -шлях фотосинтезу).
Цикл Хетча і Слека (С 4 -шлях фотосинтезу).
) також пристосувалися здійснювати фотосинтез в умовах") САМ-метаболізм Сукуленти (кактуси і рослини Родини Товстолисті (Crassulaceae)) також пристосувалися здійснювати фотосинтез в умовах різко посушливого клімату. Для них характерний добовий цикл метаболізму С 4 -кислот з утворенням яблучної кислоти вночі. Відповідно з англійським виразом Crassulacean Acid Metabolism (САМ) цей тип фотосинтезу часто скорочено називають САМметаболізм. Продихи цих рослин вдень зазвичай закриті, що запобігає втраті води, і відкриваються вночі. С 02 надходить в листки, де за участю ФЕП-карбоксилази взаємодіє з фосфоенолпіруватом, утворюючи оксалоацетат. Джерелом ФЕП служить крохмаль. Те ж саме відбувається і з С 02, який звільняється в клітинах у процесі дихання. Оксалоацетат, що утворився відновлюється під дією NADH-залежної малатдегідрогенази до яблучної кислоти, яка накопичується у вакуолях клітин листка.
САМ-метаболізм Сукуленти (кактуси і рослини Родини Товстолисті (Crassulaceae)) також пристосувалися здійснювати фотосинтез в умовах різко посушливого клімату. Для них характерний добовий цикл метаболізму С 4 -кислот з утворенням яблучної кислоти вночі. Відповідно з англійським виразом Crassulacean Acid Metabolism (САМ) цей тип фотосинтезу часто скорочено називають САМметаболізм. Продихи цих рослин вдень зазвичай закриті, що запобігає втраті води, і відкриваються вночі. С 02 надходить в листки, де за участю ФЕП-карбоксилази взаємодіє з фосфоенолпіруватом, утворюючи оксалоацетат. Джерелом ФЕП служить крохмаль. Те ж саме відбувається і з С 02, який звільняється в клітинах у процесі дихання. Оксалоацетат, що утворився відновлюється під дією NADH-залежної малатдегідрогенази до яблучної кислоти, яка накопичується у вакуолях клітин листка.
.") САМ-метаболізм (за В. В. Польовим).
САМ-метаболізм (за В. В. Польовим).
 Це призводить до закислення клітинного соку в нічний час. Як і у С 4 -рослин, оксалоацетат може бути джерелом аспартату, проте цей шлях тут менш виражений. Вдень в умовах високої температури, коли продихи закриті, малат транспортується з вакуоль в цитоплазму і там декарбоксилюється за участю малатдегідрогенази з утворенням С 02 і пірувату. С 02 надходить в хлоропласти і включається в них у цикл Кальвіна, беручи участь в синтезі цукрів. Таким чином, у рослин з фотосинтезом за типом Товстолистих багато спільного з С 4 -шляхом фотосинтезу. Однак при САМ-метаболізмі фіксація С 02 з утворенням малату (вночі) та декарбоксилювання малату з вивільненням С 02 і пірувату (вдень) розділені в часі. У С 4 рослин ці ж реакції розмежовані в просторі: перша відбувається у хлоропластах мезофіла, друга - в клітинах обкладки. При достатній кількості води ряд рослин з метаболізмом за типом Товстолистих можуть вести себе як С 3 -рослини. У свою чергу деякі рослини з С 3 -шляхом фотосинтезу при нестачі води проявляють риси САМметаболізму.
Це призводить до закислення клітинного соку в нічний час. Як і у С 4 -рослин, оксалоацетат може бути джерелом аспартату, проте цей шлях тут менш виражений. Вдень в умовах високої температури, коли продихи закриті, малат транспортується з вакуоль в цитоплазму і там декарбоксилюється за участю малатдегідрогенази з утворенням С 02 і пірувату. С 02 надходить в хлоропласти і включається в них у цикл Кальвіна, беручи участь в синтезі цукрів. Таким чином, у рослин з фотосинтезом за типом Товстолистих багато спільного з С 4 -шляхом фотосинтезу. Однак при САМ-метаболізмі фіксація С 02 з утворенням малату (вночі) та декарбоксилювання малату з вивільненням С 02 і пірувату (вдень) розділені в часі. У С 4 рослин ці ж реакції розмежовані в просторі: перша відбувається у хлоропластах мезофіла, друга - в клітинах обкладки. При достатній кількості води ряд рослин з метаболізмом за типом Товстолистих можуть вести себе як С 3 -рослини. У свою чергу деякі рослини з С 3 -шляхом фотосинтезу при нестачі води проявляють риси САМметаболізму.
 Дякую за увагу!!!
Дякую за увагу!!!